
Genome-Wide Identification and Expression of the AP2/ERF Gene Family in Morus notabilis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sources
2.2. Identification of Members of the MnAP2/ERF Gene Family
2.3. Chromosomal Mapping and Numbering of MnAP2/ERF Gene Family Members
2.4. Analysis of Physicochemical Properties of MnAP2/ERF Gene Family Members
2.5. Evolutionary Tree Construction of MnAP2/ERF Gene Family Members
2.6. Motif Analysis and Gene Structure Analysis of MnAP2/ERF Gene Family
2.7. Promoter Cis-Acting Elements of the MnAP2/ERF Gene Family
2.8. Tertiary Structure and Protein–Protein Interaction (PPI) Analysis of MnAP2/ERF Gene Family Proteins
2.9. Gene Repeat Events and Collinearity Analysis of the MnAP2/ERF Gene Family
2.10. Analysis of the Expression Pattern of the MnAP2/ERF Gene Family in Different Organs of M. notabilis
3. Results and Analysis
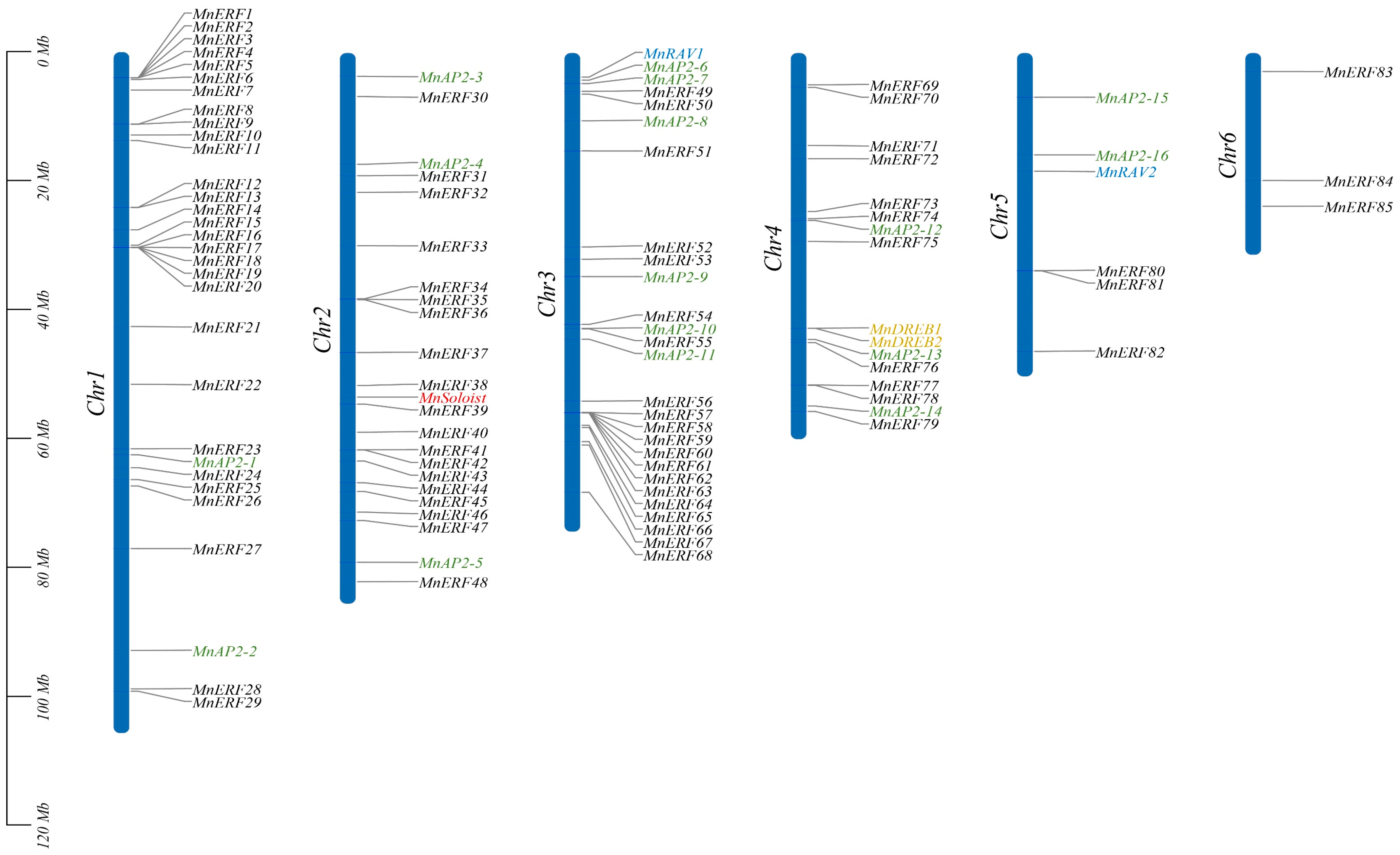
3.1. Chromosomal Location and Nomenclature of MnAP2/ERF Gene Family Members
3.2. Physicochemical Properties of MnAP2/ERF Gene Family Members
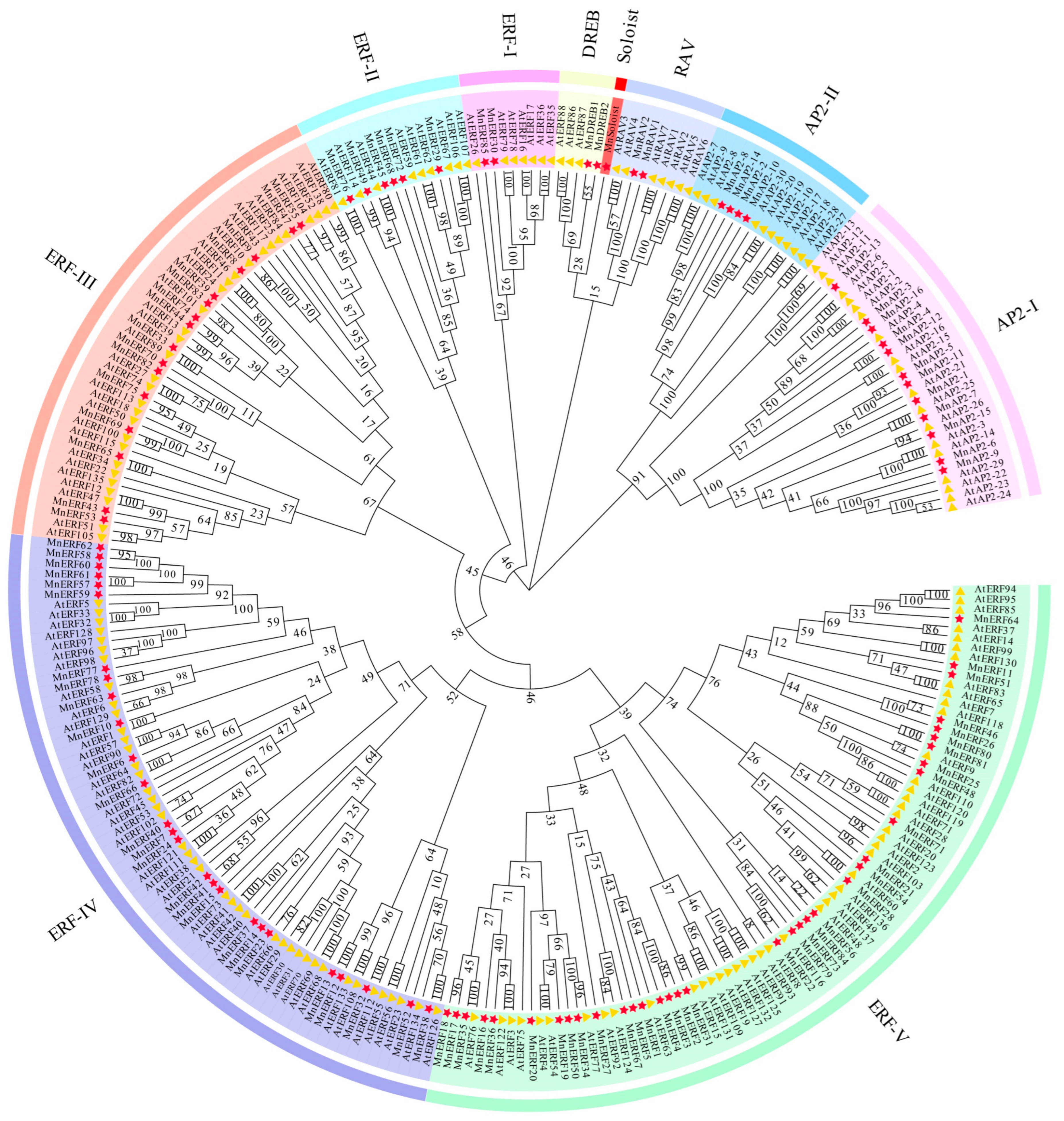
3.3. Evolutionary Analysis of AP2/ERF Gene Family
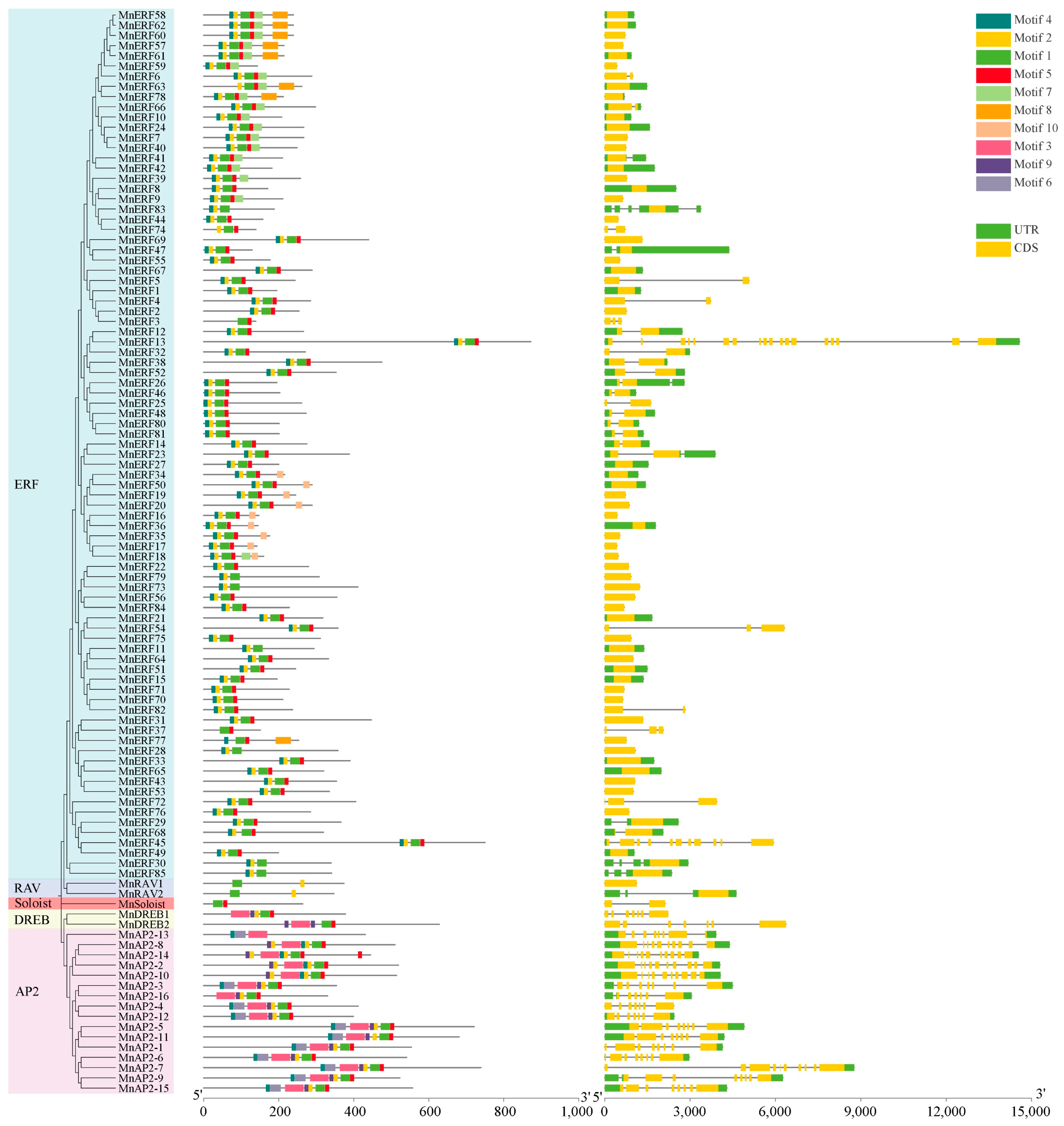
3.4. Conserved Motifs and Gene Structures of MnAP2/ERF Gene Family Members
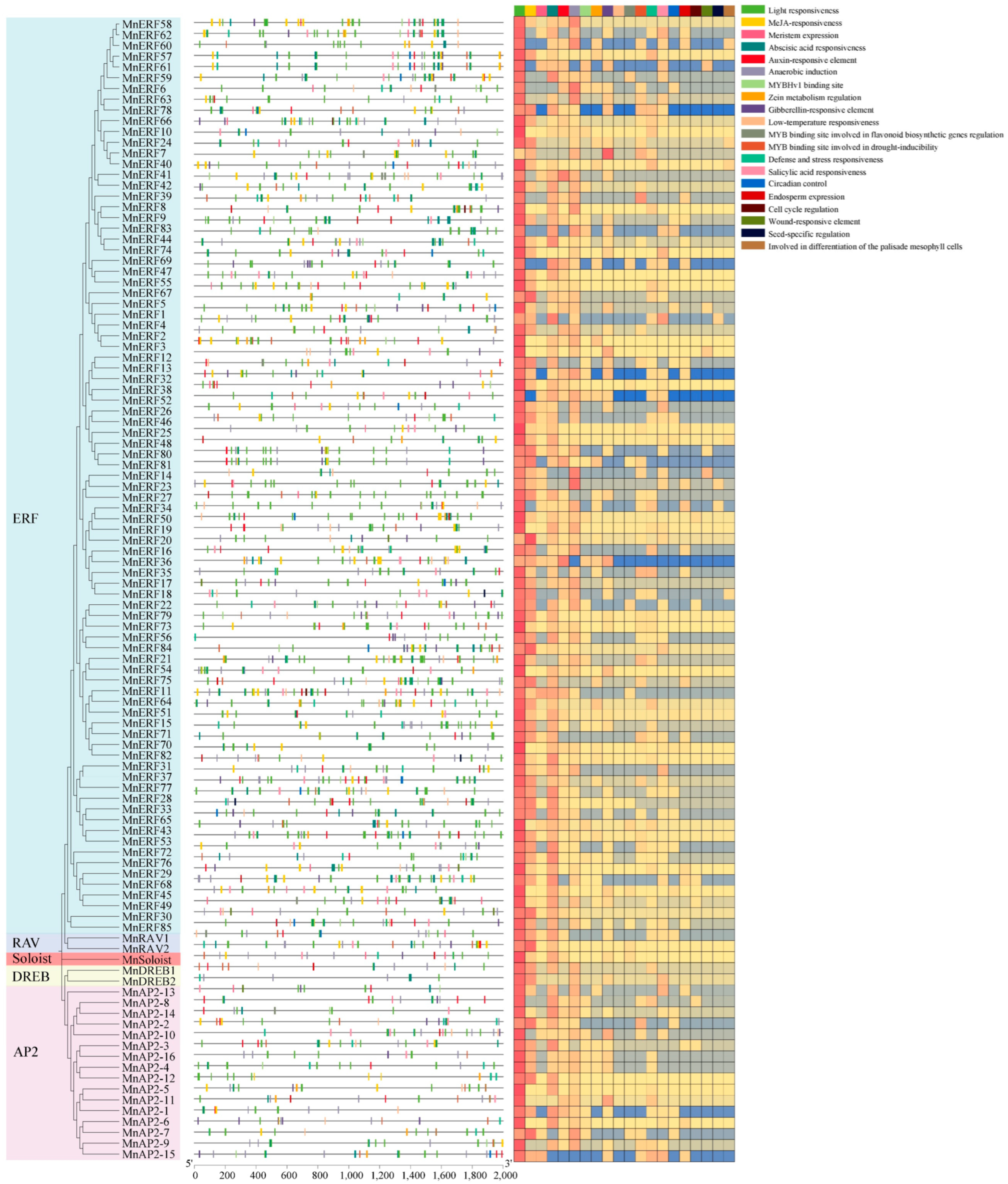
3.5. Cis-Acting Elements in the Promoter Region of MnAP2/ERF
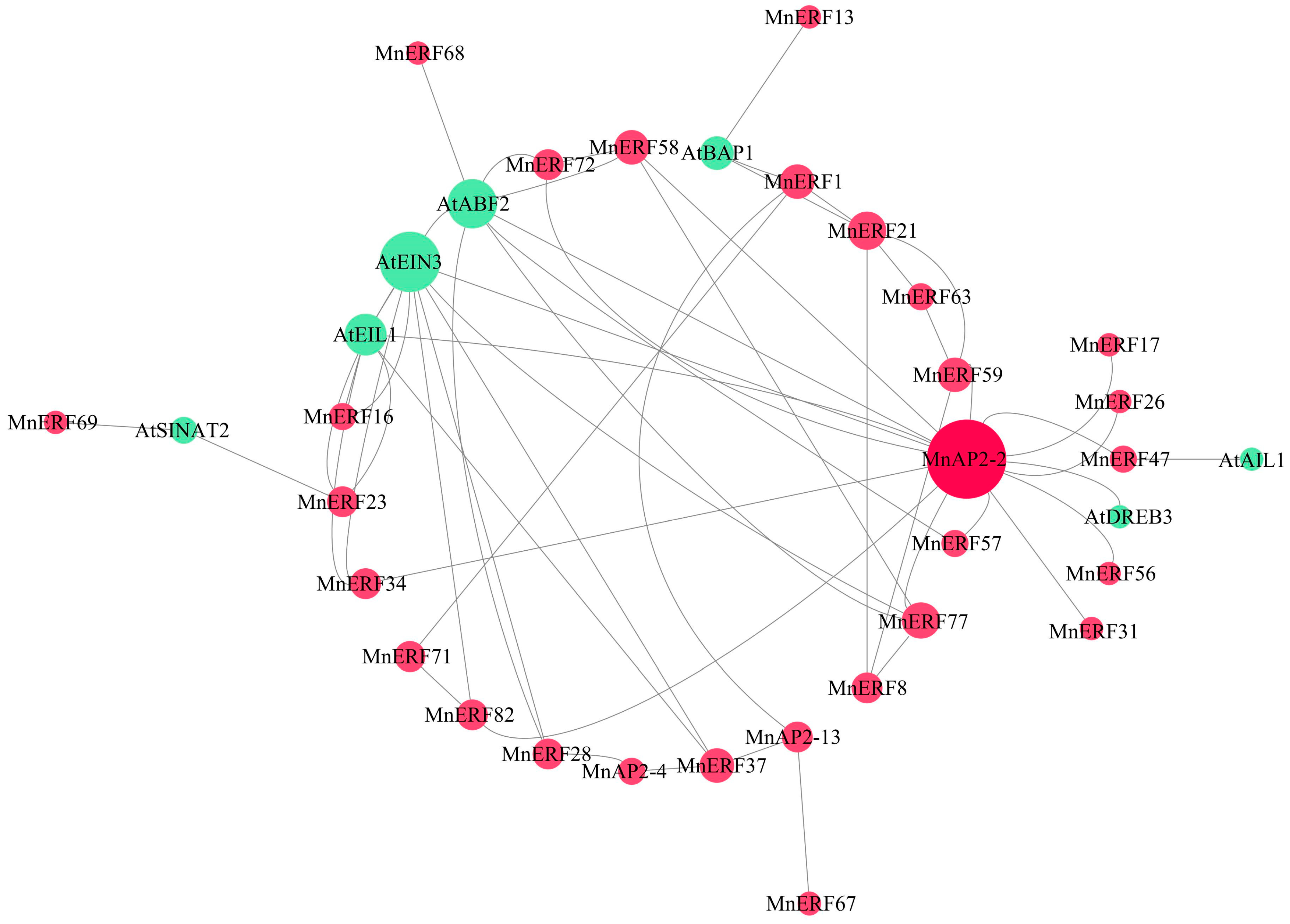
3.6. Tertiary Structure and Protein–Protein Interaction Association of MnAP2/ERF Gene Family Proteins
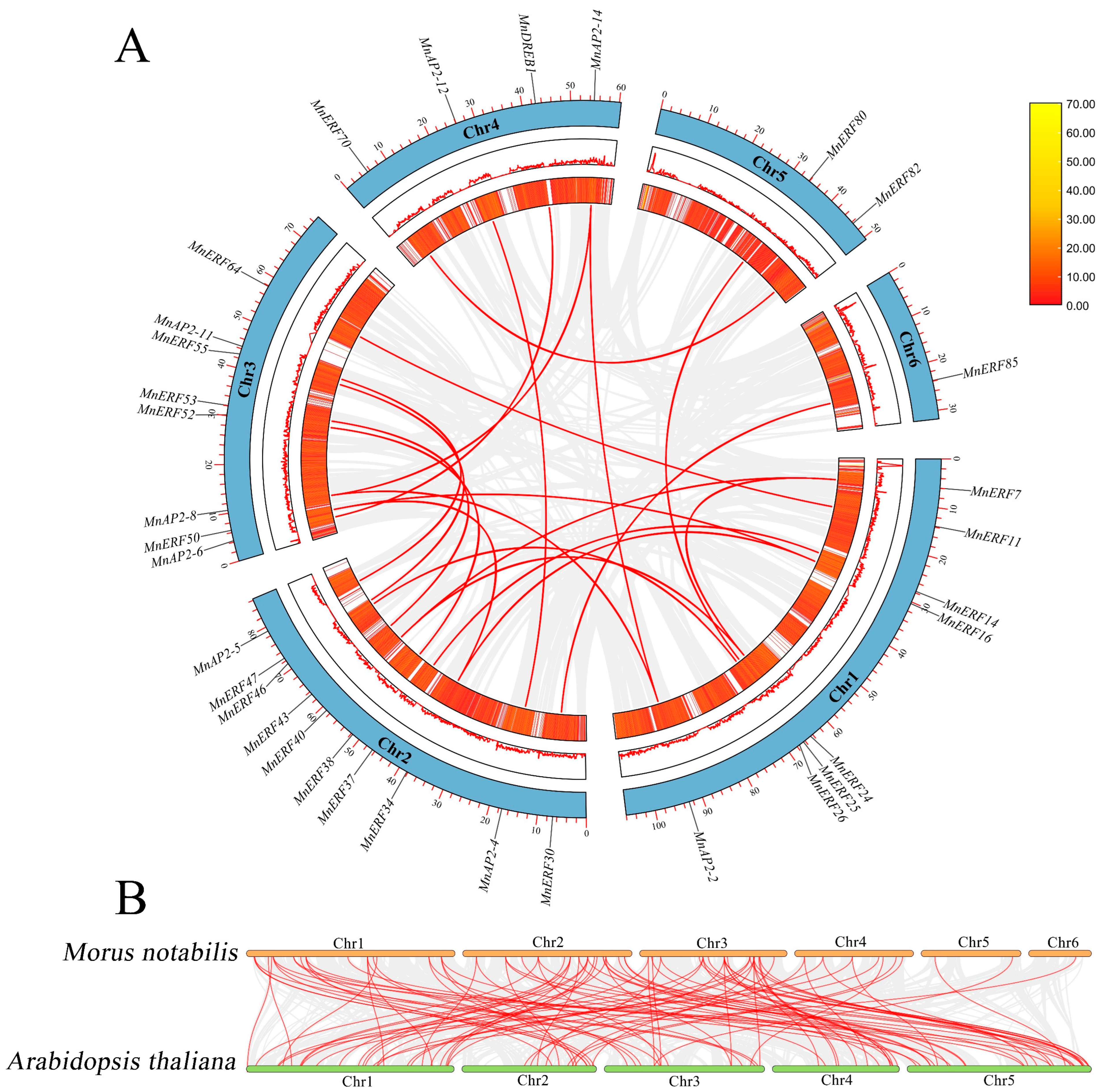
3.7. Gene Duplication Events and Species Collinearity of the MnAP2/ERF Gene Family
3.8. Expression Analysis of MnAP2/ERF Gene Family in M. notabilis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baciu, E.-D.; Baci, G.-M.; Moise, A.R.; Dezmirean, D.S. A Status Review on the Importance of Mulberry (Morus spp.) and Prospects towards Its Cultivation in a Controlled Environment. Horticulturae 2023, 9, 444. [Google Scholar] [CrossRef]
- Rohela, G.K.; Shukla, P.; Muttanna; Kumar, R.; Chowdhury, S.R. Mulberry (Morus spp.): An ideal plant for sustainable development. In Trees, Forests and People; Elsevier: Amsterdam, The Netherlands, 2020; Volume 2, p. 100011. [Google Scholar] [CrossRef]
- Memete, A.R.; Timar, A.V.; Vuscan, A.N.; Miere Groza, F.; Venter, A.C.; Vicas, S.I. Phytochemical Composition of Different Botanical Parts of Morus Species, Health Benefits and Application in Food Industry. Plants 2022, 11, 152. [Google Scholar] [CrossRef]
- Jan, B.; Parveen, R.; Zahiruddin, S.; Khan, M.U.; Mohapatra, S.; Ahmad, S. Nutritional constituents of mulberry and their potential applications in food and pharmaceuticals: A review. Saudi J. Biol. Sci. 2021, 28, 3909–3921. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjya, D.; Sadat, A.; Dam, P.; Buccini, D.F.; Mondal, R.; Biswas, T.; Biswas, K.; Sarkar, H.; Bhuimali, A.; Katı, A.; et al. Current concepts and prospects of mulberry fruits for nutraceutical and medicinal benefits. Curr. Opin. Food Sci. 2021, 40, 121–135. [Google Scholar] [CrossRef]
- Guha, A.; Reddy, A.R. Leaf Functional Traits and Stem Wood Characteristics Influencing Biomass Productivity of Mulberry (Morus spp. L) Genotypes Grown in Short-Rotation Coppice System. Bioenerg. Res. 2013, 6, 547–563. [Google Scholar] [CrossRef]
- Lu, L.; Tang, Y.; Xie, J.; Yuan, Y. The role of marginal agricultural land-based mulberry planting in biomass energy production. Renew. Energ. 2009, 34, 1789–1794. [Google Scholar] [CrossRef]
- Andallu, B.; Suryakantham, V.; Lakshmi Srikanthi, B.; Reddy, G.K. Effect of mulberry (Morus indica L.) therapy on plasma and erythrocyte membrane lipids in patients with type 2 diabetes. Clin. Chim. Acta. 2001, 314, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Dubey, V.; Khan, S.; Shah, K.W.; Raghuwanshi, R. Standardization of Protocol for In Vitro Micropropagation of Morus alba L., An Important Economical and Medicinal Plant. J. Pharm. Biomed. 2020, 8, 46. [Google Scholar] [CrossRef]
- Baranwal, V.; Negi, N.; Khurana, P. Genome-wide Identification and Structural, Functional and Evolutionary Analysis of WRKY Components of Mulberry. Sci. Rep. 2016, 6, 30794. [Google Scholar] [CrossRef]
- Vijayan, K.; Arunakumar, G.S.; Gnanesh, B.N.; Sangannavar, P.A.; Ramesha, A.; Zhao, W. Genomic Designing for Biotic Stress Resistance in Mulberry. In Genomic Designing for Biotic Stress Resistant Technical Crops; Kole, C., Ed.; Springer: Cham, Switzerland, 2022. [Google Scholar] [CrossRef]
- Guy, J.L.; Mor, G.G. Transcription Factor-Binding Site Identification and Enrichment Analysis. Methods Mol. Biol. 2021, 2255, 241–261. [Google Scholar] [CrossRef]
- Garg, A.; Futcher, B.; Leatherwood, J.K. A new transcription factor for mitosis: In Schizosaccharomyces pombe, the RFX transcription factor Sak1 works with forkhead factors to regulate mitotic expression. Nucleic Acids Res. 2015, 43, 6874–6888. [Google Scholar] [CrossRef]
- Jofuku, K.D.; den Boer, B.G.; Van Montagu, M.; Okamuro, J.K. Control of Arabidopsis flower and seed development by the homeotic gene APETALA2. Plant Cell. 1994, 6, 1211–1225. [Google Scholar] [CrossRef] [PubMed]
- Wessler, S.R. Homing into the origin of the AP2 DNA binding domain. Trends Plant Sci. 2005, 10, 54–56. [Google Scholar] [CrossRef]
- Agarwal, P.K.; Agarwal, P.; Reddy, M.K.; Sopory, S.K. Role of DREB transcription factors in abiotic and biotic stress tolerance in plants. Plant Cell Rep. 2006, 25, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Hu, L.; Jiang, W. Understanding AP2/ERF Transcription Factor Responses and Tolerance to Various Abiotic Stresses in Plants: A Comprehensive Review. Int. J Mol. Sci. 2024, 25, 893. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhuo, W.; Luo, J.; Yang, S.Y.; Lu, L.M.; Li, L.Q. Expression vector construction and expression pattern analysis of ERF gene NtRAP2-7 in tobacco. Acta Agri. Boreali Sin. 2018, 33, 104–111. [Google Scholar]
- Gutterson, N.; Reuber, T.L. Regulation of disease resistance pathways by AP2/ERF transcription factors. Curr. Opin. Plant Biol. 2004, 7, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Karanja, B.K.; Xu, L.; Wang, Y.; Tang, M.; M’mbone Muleke, E.; Dong, J.; Liu, L. Genome-wide characterization of the AP2/ERF gene family in radish (Raphanus sativus L.): Unveiling evolution and patterns in response to abiotic stresses. Gene 2019, 718, 144048. [Google Scholar] [CrossRef] [PubMed]
- Ritonga, F.N.; Ngatia, J.N.; Wang, Y.; Khoso, M.A.; Farooq, U.; Chen, S. AP2/ERF, an important cold stress-related transcription factor family in plants: A review. Physiol. Mol. Biol. Plants 2021, 27, 1953–1968. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, Y.C.; Lee, J.H.; Oh, S.K.; Chung, E.; Lee, S.; Lee, Y.H.; Choi, D.; Park, J.M. Identification of a CaRAV1 possessing an AP2/ERF and B3 DNA-binding domain from pepper leaves infected with Xanthomonas ax-onopodis pv. glycines 8ra by differential display. Biochim. Biophys. Acta 2005, 1729, 141–146. [Google Scholar] [CrossRef]
- Hassan, S.; Berk, K.; Aronsson, H. Evolution and identification of DREB transcription factors in the wheat genome: Modeling, docking and simulation of DREB proteins associated with salt stress. J. Biomol. Struct. Dyn. 2022, 40, 7191–7204. [Google Scholar] [CrossRef]
- Zhao, M.; Haxim, Y.; Liang, Y.; Qiao, S.; Gao, B.; Zhang, D.; Li, X. Genome-wide investigation of AP2/ERF gene family in the desert legume Eremosparton songoricum: Identification, classification, evolution, and expression profiling under drought stress. Front. Plant Sci. 2022, 13, 885694. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.L.; Yao, W.J.; Wang, S.J.; Jiang, T.B.; Zhou, B.R. Bioinformatics analysis of poplar AP2/ERF transcription factor family. J. Northeast Univ. 2015, 43, 21–29. [Google Scholar]
- Chen, Y.; Sun, M.Z.; Jia, B.W.; Leng, Y.; Sun, X.L. Advances in molecular mechanisms of rice AP2/ERF transcription factors involved in stress response. Zuo Wu Xue Bao 2022, 48, 781–790. [Google Scholar]
- Zhang, J.; Liao, J.; Ling, Q.; Xi, Y.; Qian, Y. Genome-wide identification and expression profiling analysis of maize AP2/ERF superfamily genes reveal essential roles in abiotic stress tolerance. BMC Genom. 2022, 23, 125. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zhang, J.; Liu, J.F.; Yang, J.B.; Li, M.X.; Chen, J.B.; Yang, S.Q. Bioinformatics identification and characterization of mung bean AP2/ERF transcription factor family. Mol. Plant Breed. 2020, 18, 6605–6617. [Google Scholar]
- Huang, Y.Z.; Qian, W.; Qiu, S.; Wang, W.X.; Huang, H.H.; Lin, E.P. Identification and expression analysis of AP2/ERF gene family in Betula alba. J. Zhejiang AF Univ. 2022, 39, 1183–1193. [Google Scholar]
- Hu, Y.X.; Wang, Y.X.; Liu, X.F.; Li, J.Y. Arabidopsis RAV1 is down-regulated by brassinosteroid and may act as a negative regulator during plant development. Cell Res. 2004, 14, 8–15. [Google Scholar] [CrossRef]
- Sohn, K.H.; Lee, S.C.; Jung, H.W.; Hong, J.K.; Hwang, B.K. Expression and functional roles of the pepper pathogen-induced transcription factor RAV1 in bacterial disease resistance, and drought and salt stress tolerance. Plant Mol. Biol. 2006, 61, 897–915. [Google Scholar] [CrossRef]
- Li, X.J.; Li, M.; Zhou, Y.; Hu, S.; Hu, R.; Chen, Y.; Li, X.B. Overexpression of cotton RAV1 gene in Arabidopsis confers transgenic plants high salinity and drought sensitivity. PLoS ONE 2015, 10, e0118056. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Chen, M.; Li, L.; Xu, Z.; Chen, X.; Guo, J.; Ma, Y. Overexpression of the soybean GmERF3 gene, an AP2/ERF type transcription factor for increased tolerances to salt, drought, and diseases in transgenic tobacco. J. Exp. Bot. 2009, 60, 3781–3796. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Qi, L.; Liu, X.; Cai, S.; Xu, H.; Huang, R.; Li, J.; Wei, X.; Zhang, Z. The wheat ethylene response factor transcription factor pathogen-induced ERF1 mediates host responses to both the necrotrophic pathogen Rhizoctonia cerealis and freezing stresses. Plant Physiol. 2014, 164, 1499–1514. [Google Scholar] [CrossRef]
- Hong, J.P.; Kim, W.T. Isolation and functional characterization of the Ca-DREBLP1 gene encoding a dehydration-responsive element binding-factor-like protein 1 in hot pepper (Capsicum annuum L. cv. Pukang). Planta 2005, 220, 875–888. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, Z.; Zhang, H.; Wang, X.C.; Huang, R. Transcriptional modulation of ethylene response factor protein JERF3 in the oxidative stress response enhances tolerance of tobacco seedlings to salt, drought, and freezing. Plant Physiol. 2008, 148, 1953–1963. [Google Scholar] [CrossRef]
- Ito, Y.; Katsura, K.; Maruyama, K.; Taji, T.; Kobayashi, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of rice DREB1/CBF-type transcription factors involved in cold-responsive gene expression in transgenic rice. Plant Cell Physiol. 2006, 47, 141–153. [Google Scholar] [CrossRef]
- Qin, F.; Kakimoto, M.; Sakuma, Y.; Maruyama, K.; Osakabe, Y.; Tran, L.S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Regulation and functional analysis of ZmDREB2A in response to drought and heat stresses in Zea mays L. Plant J. 2007, 50, 54–69. [Google Scholar] [CrossRef]
- Gilmour, S.J.; Sebolt, A.M.; Salazar, M.P.; Everard, J.D.; Thomashow, M.F. Overexpression of the Arabidopsis CBF3 transcriptional activator mimics multiple biochemical changes associated with cold acclimation. Plant Physiol. 2000, 124, 1854–1865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Pan, X.; Liu, S.; Lin, W.; Li, Y.; Zhang, X. Genome-wide analysis of AP2/ERF transcription factors in pineapple reveals functional divergence during flowering induction mediated by ethylene and floral organ development. Genomics 2021, 113, 474–489. [Google Scholar] [CrossRef]
- Ye, B.B.; Shang, G.D.; Pan, Y.; Xu, Z.G.; Zhou, C.M.; Mao, Y.B.; Bao, N.; Sun, L.; Xu, T.; Wang, J.W. AP2/ERF Transcription Factors Integrate Age and Wound Signals for Root Regeneration. Plant Cell 2020, 32, 226–241. [Google Scholar] [CrossRef]
- Soma, F.; Kitomi, Y.; Kawakatsu, T.; Uga, Y. Life-Cycle Multiomics of Rice Shoots Reveals Growth Stage-Specific Effects of Drought Stress and Time-Lag Drought Responses. Plant Cell Physiol. 2024, 65, 156–168. [Google Scholar] [CrossRef]
- Dossa, K.; Wei, X.; Li, D.; Fonceka, D.; Zhang, Y.; Wang, L.; Yu, J.; Boshou, L.; Diouf, D.; Cissé, N.; et al. Insight into the AP2/ERF transcription factor superfamily in sesame and expression profiling of DREB subfamily under drought stress. BMC Plant Biol. 2016, 16, 171. [Google Scholar] [CrossRef] [PubMed]
- Swinka, C.; Hellmann, E.; Zwack, P.; Banda, R.; Rashotte, A.M.; Heyl, A. Cytokinin Response Factor 9 Represses Cytokinin Responses in Flower Development. Int. J. Mol. Sci. 2023, 24, 4380. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Wang, H.; Liu, J.; Chen, L.; Xia, X.; Wei, W.; Yang, Z.; Yuan, J.; Luo, Y.; He, N. The gap-free genome of mulberry elucidates the architecture and evolution of polycentric chromosomes. Hortic. Res. 2023, 10, uhad111. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; et al. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant. 2023, 16, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Jiao, F.; Luo, R.; Dai, X.; Liu, H.; Yu, G.; Han, S.; Lu, X.; Su, C.; Chen, Q.; Song, Q.; et al. Chromosome-Level Reference Genome and Population Genomic Analysis Provide Insights into the Evolution and Improvement of Domesticated Mulberry (Morus alba). Mol. Plant 2020, 13, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Charfeddine, M.; Saïdi, M.N.; Charfeddine, S.; Hammami, A.; Gargouri Bouzid, R. Genome-wide analysis and expression profiling of the ERF transcription factor family in potato (Solanum tuberosum L.). Mol. Biotechnol. 2015, 57, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.P.; Bo, F.S.; Tian, L.Z.; Jiang, H.P.; Zhao, X. Bioinformatic analysis of AP2/EREBP transcription factors associated with resistance to soybean cystonematode disease. J. Northeast. Univ. 2020, 51, 1–8+73. [Google Scholar]
- Sharma, M.K.; Kumar, R.; Solanke, A.U.; Sharma, R.; Tyagi, A.K.; Sharma, A.K. Identification, phylogeny, and transcript profiling of ERF family genes during development and abiotic stress treatments in tomato. Mol. Genet. Genom. 2020, 284, 455–475. [Google Scholar] [CrossRef]
- Kasuga, M.; Miura, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. A combination of the Arabidopsis DREB1A gene and stress-inducible rd29A promoter improved drought- and low-temperature stress tolerance in tobacco by gene transfer. Plant Cell Physiol. 2004, 45, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Krishnaswamy, S.; Verma, S.; Rahman, M.H.; Kav, N.N. Functional characterization of four APETALA2-family genes (RAP2.6, RAP2.6L, DREB19 and DREB26) in Arabidopsis. Plant Mol. Biol. 2011, 75, 107–127. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, L.; Wang, L.; Wang, S.H.; Cheng, X.Z. VrDREB2A, a DREB-binding transcription factor from Vigna radiata, increased drought and high-salt tolerance in transgenic Arabidopsis thaliana. J. Plant Res. 2016, 129, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Rong, W.; Qi, L.; Wang, A.; Ye, X.; Du, L.; Liang, H.; Xin, Z.; Zhang, Z. The ERF transcription factor TaERF3 promotes tolerance to salt and drought stresses in wheat. Plant Biotechnol. J. 2014, 12, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yang, R.; Liang, Y.; Wang, Y.; Li, X. The DREB A-5 Transcription Factor ScDREB5 from Syntrichia caninervis Enhanced Salt Tolerance by Regulating Jasmonic Acid Biosynthesis in Transgenic Arabidopsis. Front. Plant Sci. 2022, 13, 857396. [Google Scholar] [CrossRef]
- Zhu, Q.; Zhang, J.; Gao, X.; Tong, J.; Xiao, L.; Li, W.; Zhang, H. The Arabidopsis AP2/ERF transcription factor RAP2.6 participates in ABA, salt and osmotic stress responses. Gene 2010, 57, 1–12. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Motif | Length/aa | Conserved Amino Acid Sequences |
|---|---|---|
| Motif 1 | 26 | VWLGTFDTAEEAARAYDVAALKLRGS |
| Motif 2 | 11 | RPWGKWVAEIR |
| Motif 3 | 50 | RAYDLAALKYWGPSTTINFPLSBYEKELEEMKNMTKZEFVASLRRKSSGF |
| Motif 4 | 11 | RRSSKYRGVRR |
| Motif 5 | 11 | KAKLNFPELTY |
| Motif 6 | 29 | HRWTGRYEAHLWDNSCRREGQSRKGRQGA |
| Motif 7 | 23 | SLPRPASTDAKDIQAAAAEAAAA |
| Motif 8 | 41 | EEEZRVFYVDEEEVFGMPSFFADLAEGLLLSPPPSYMSSDD |
| Motif 9 | 11 | GASQYRGVTRH |
| Motif 10 | 17 | VIELEYLDDKYLEELLD |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dou, H.; Wang, T.; Zhou, X.; Feng, X.; Tang, W.; Quan, J.; Bi, H. Genome-Wide Identification and Expression of the AP2/ERF Gene Family in Morus notabilis. Forests 2024, 15, 697. https://doi.org/10.3390/f15040697
Dou H, Wang T, Zhou X, Feng X, Tang W, Quan J, Bi H. Genome-Wide Identification and Expression of the AP2/ERF Gene Family in Morus notabilis. Forests. 2024; 15(4):697. https://doi.org/10.3390/f15040697
Chicago/Turabian StyleDou, Hao, Tiantian Wang, Xin Zhou, Xi Feng, Wu Tang, Jin’e Quan, and Huitao Bi. 2024. "Genome-Wide Identification and Expression of the AP2/ERF Gene Family in Morus notabilis" Forests 15, no. 4: 697. https://doi.org/10.3390/f15040697
APA StyleDou, H., Wang, T., Zhou, X., Feng, X., Tang, W., Quan, J., & Bi, H. (2024). Genome-Wide Identification and Expression of the AP2/ERF Gene Family in Morus notabilis. Forests, 15(4), 697. https://doi.org/10.3390/f15040697