
Structural and Successional Dynamics of Old-Growth Mixed Oak Forests in the West Qinling Mountains

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Monitoring Plot Data
2.2. Data Analysis
3. Results
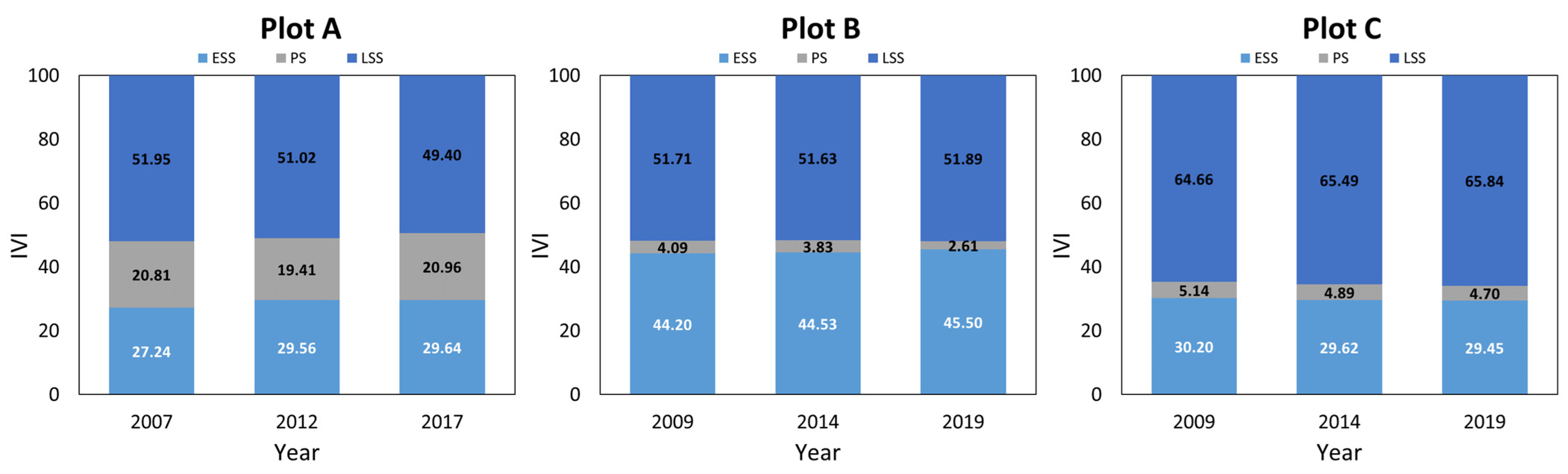
3.1. Tree Species Composition and Diversity
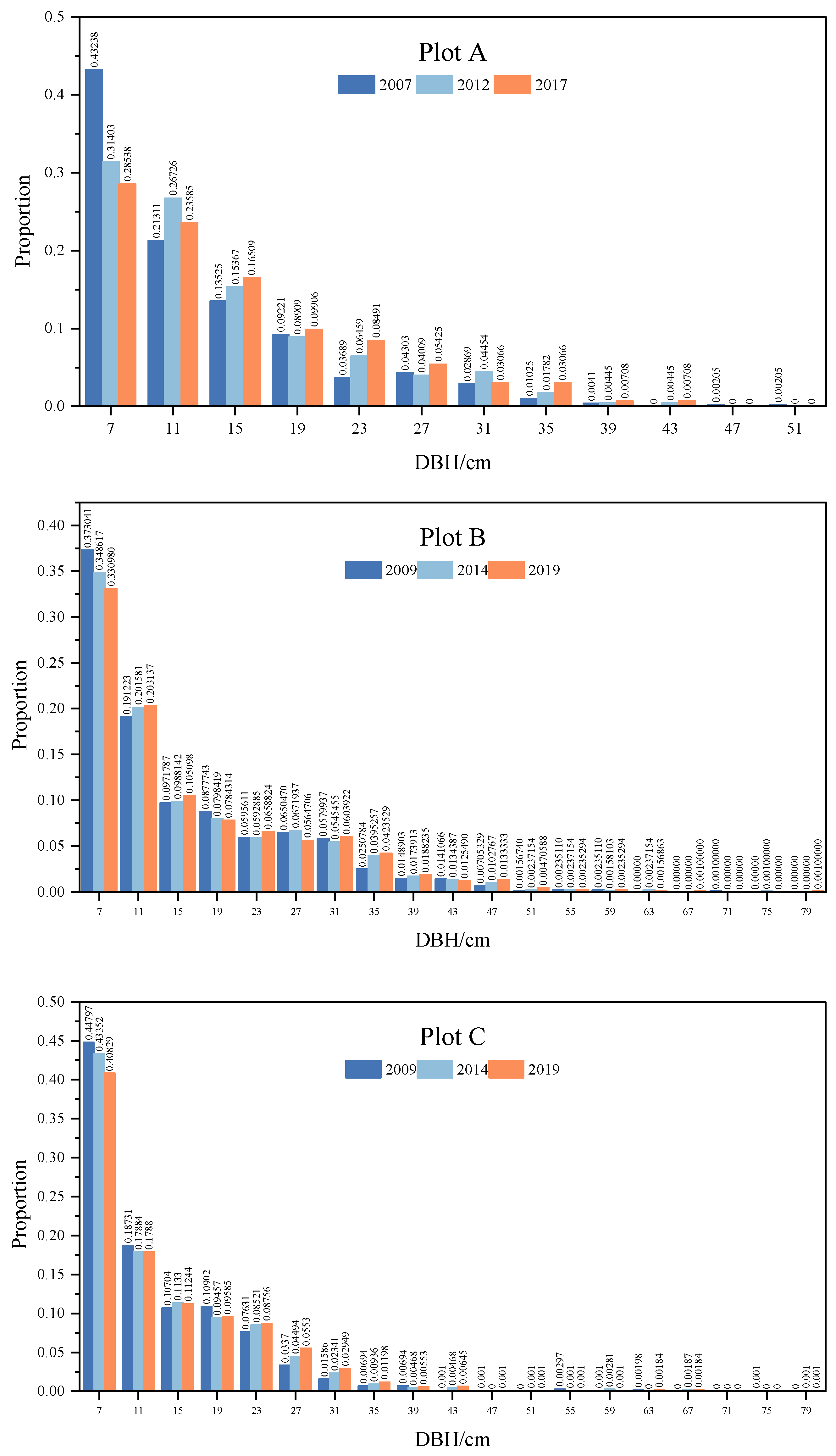
3.2. Tree Diameter Distribution and Demographic Parameters
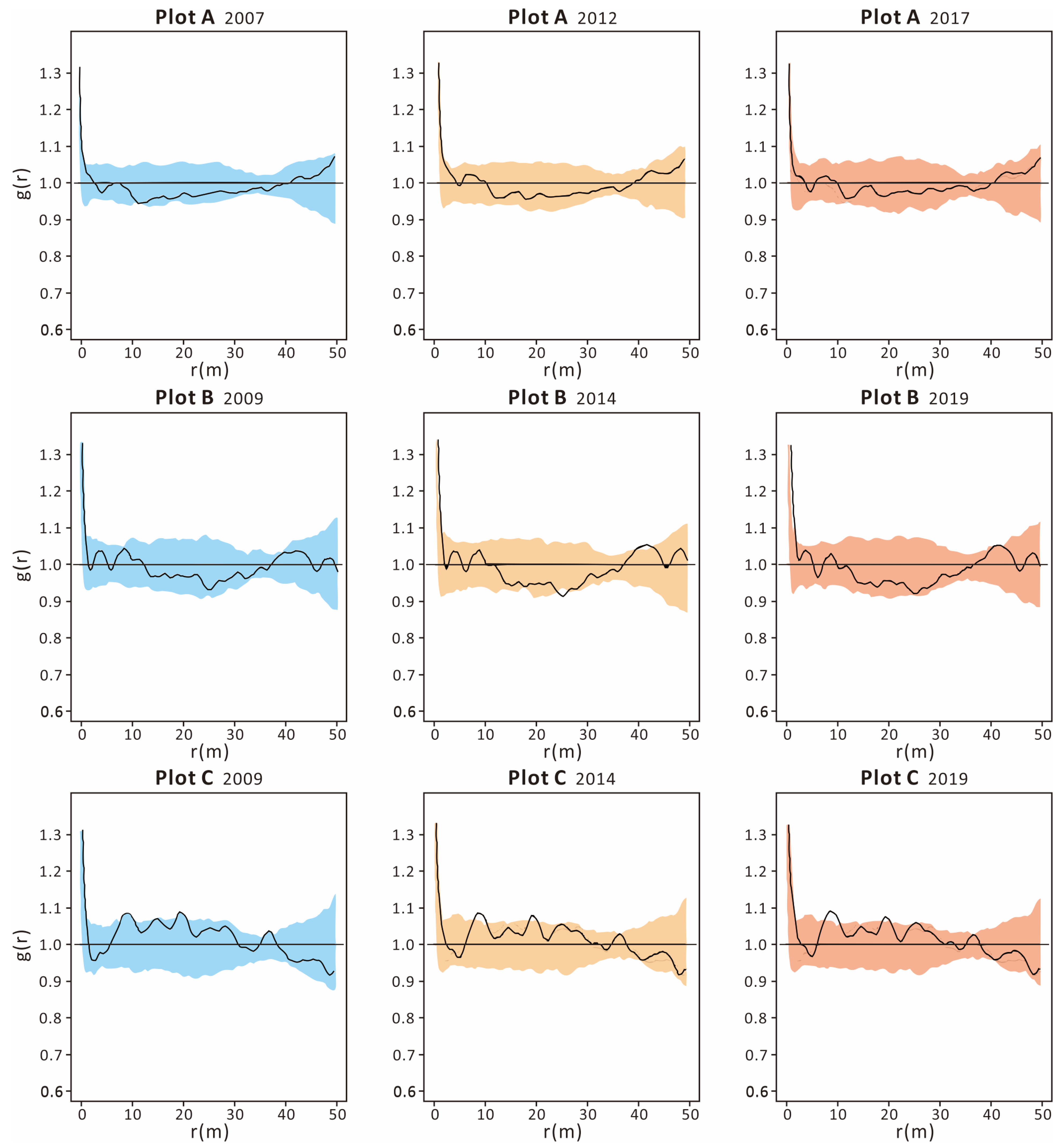
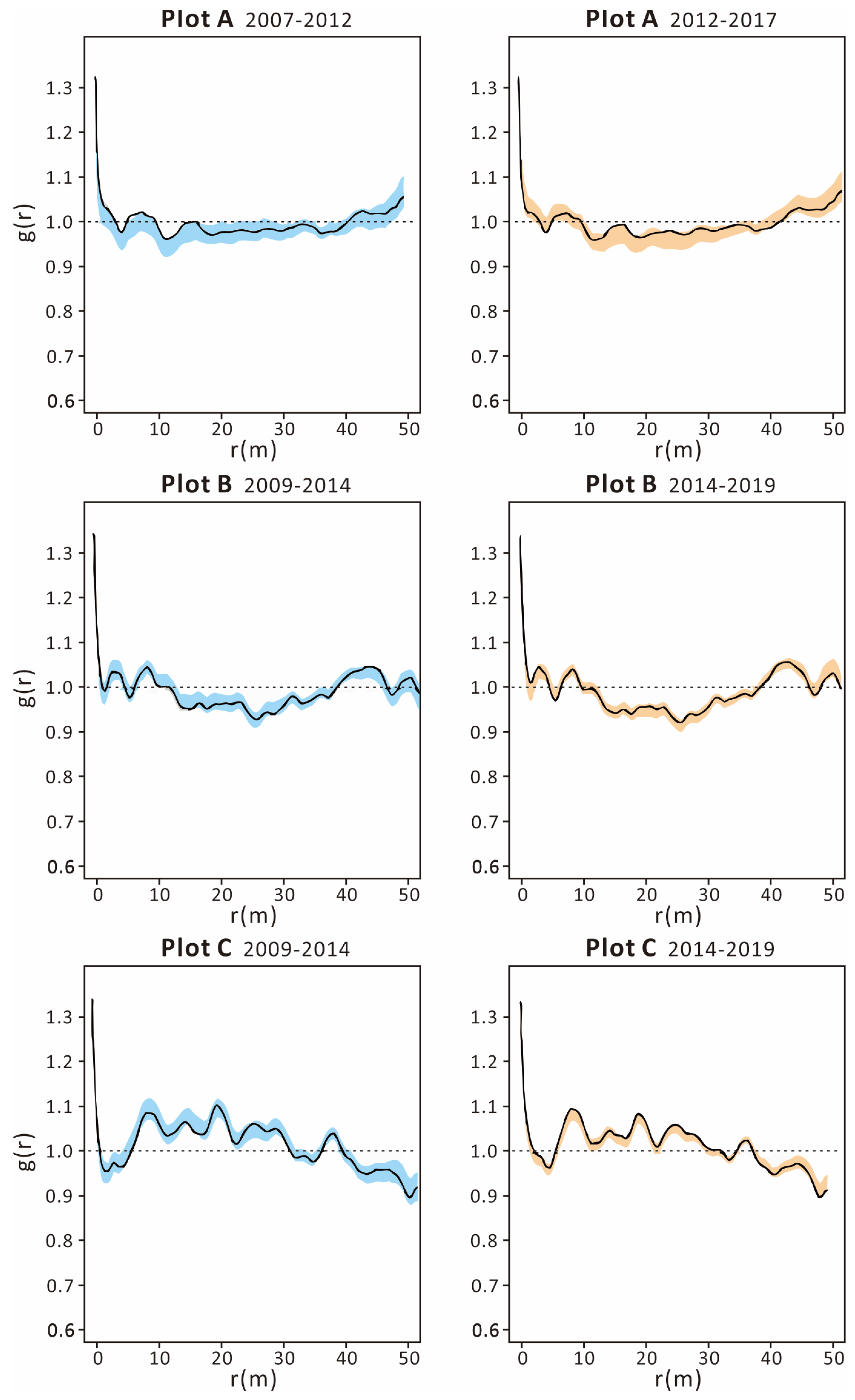
3.3. Tree Dynamics Pattern and Individual Distribution Pattern

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Group | Recruitment Rate (%) | Mortality Rate (%) | Dynamics Rate (%) | Half-Life Time (Year) | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plot A | Plot B | Plot C | Plot A | Plot B | Plot C | Plot A | Plot B | Plot C | Plot A | Plot B | Plot C | |||||||||||||
| 2007– 2012 | 2012– 2017 | 2009– 2014 | 2014– 2019 | 2009– 2014 | 2014– 2019 | 2007– 2012 | 2012– 2017 | 2009– 2014 | 2014– 2019 | 2009– 2014 | 2014– 2019 | 2007– 2012 | 2012– 2017 | 2009– 2014 | 2014– 2019 | 2009– 2014 | 2014– 2019 | 2007– 2012 | 2012– 2017 | 2009– 2014 | 2014– 2019 | 2009– 2014 | 2014– 2019 | |
| Pioneer | 0.00 | 1.06 | 1.38 | 0.70 | 0.00 | 0.56 | 1.80 | 3.02 | 3.81 | 1.37 | 1.08 | 0.56 | 0.90 | 2.04 | 2.60 | 1.04 | 0.54 | 0.56 | 38.1 | 22.6 | 17.9 | 50.2 | 64.1 | 123.0 |
| Early successional | 0.07 | 1.70 | 1.11 | 0.80 | 2.81 | 0.96 | 1.72 | 1.70 | 1.37 | 0.70 | 1.24 | 0.51 | 0.90 | 1.70 | 1.24 | 0.75 | 2.03 | 0.74 | 39.9 | 40.3 | 50.4 | 99.3 | 55.4 | 134.7 |
| Late-successional | 0.00 | 0.00 | 0.51 | 0.42 | 1.38 | 0.50 | 0.87 | 1.85 | 0.38 | 0.13 | 1.02 | 0.50 | 0.44 | 0.93 | 0.45 | 0.28 | 1.20 | 0.50 | 79.2 | 37.2 | 180.8 | 549.3 | 67.3 | 138.9 |
| Species Group | Recruitment Rate (%) | Mortality Rate (%) | Dynamics Rate (%) | Half-Life Time (Year) | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plot A | Plot B | Plot C | Plot A | Plot B | Plot C | Plot A | Plot B | Plot C | Plot A | Plot B | Plot C | |||||||||||||
| 2007– 2012 | 2012–2017 | 2009–2014 | 2014–2019 | 2009–2014 | 2014–2019 | 2007– 2012 | 2012–2017 | 2009–2014 | 2014–2019 | 2009–2014 | 2014–2019 | 2007– 2012 | 2012–2017 | 2009–2014 | 2014–2019 | 2009–2014 | 2014–2019 | 2007– 2012 | 2012–2017 | 2009–2014 | 2014–2019 | 2009–2014 | 2014–2019 | |
| Pioneer | 1.86 | 1.76 | 4.61 | 1.88 | 2.90 | 2.02 | 6.39 | 4.12 | 2.99 | 0.11 | 0.21 | 1.45 | 4.13 | 2.94 | 3.80 | 1.00 | 1.56 | 1.74 | 10.5 | 16.5 | 22.8 | 628.5 | 329.2 | 47.4 |
| Early successional | 3.13 | 2.82 | 2.04 | 2.01 | 3.66 | 2.60 | 2.12 | 0.83 | 1.09 | 0.55 | 0.53 | 0.54 | 2.63 | 1.83 | 1.57 | 1.28 | 2.10 | 1.57 | 32.3 | 83.5 | 63.5 | 126.4 | 129.5 | 126.9 |
| Late-successional | 3.62 | 3.04 | 2.04 | 1.37 | 3.52 | 2.39 | 0.29 | 0.99 | 0.13 | 0.11 | 0.43 | 0.10 | 1.96 | 2.02 | 1.09 | 0.74 | 1.98 | 1.25 | 260.3 | 69.0 | 525.3 | 598.9 | 159.8 | 685.9 |
4. Discussion
4.1. Tree Species Composition and Diversity
4.2. Tree Size Differentiation
4.3. Dynamics Characteristics
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Species | Group | Attribute |
|---|---|---|
| Lindera obtusiloba Blume | Early successional species group | shrub |
| Sorbus alnifolia (Siebold & Zucc.) C. Koch | macrophanerophytes | |
| Carpinus turczaninowii Hance | macrophanerophytes | |
| Carpinus stipulata H. J. P. Winkl. | macrophanerophytes | |
| Meliosma cuneifolia Franch. var. glabriuscula Cuf | macrophanerophytes | |
| Carpinus henryana (H. J. P. Winkl.) H. J. P. Winkl. | macrophanerophytes | |
| Cornus hemsleyi C. K. Schneid. & Wangerin | macrophanerophytes | |
| Carya cathayensis Sarg. | macrophanerophytes | |
| Acer davidii Frarich. | macrophanerophytes | |
| Amelanchier alnifolia | deciduous tree or shrub | |
| Malus hupehensis (Pamp.) Rehder | macrophanerophytes | |
| Fraxinus sogdiana Bunge | macrophanerophytes | |
| Tilia paucicostata Maxim. | macrophanerophytes | |
| Acer tetramerum Pax | macrophanerophytes | |
| Staphylea holocarpa Hemsl. | deciduous tree or shrub | |
| Crataegus kansuensis E. H. Wilson | macrophanerophytes or shrub | |
| Viburnum dilatatum Thunb. | shrub | |
| Fraxinus paxiana Lingelsh. | macrophanerophytes | |
| Tilia tuan Szyszyl. | macrophanerophytes | |
| Ulmus macrocarpa Hance | deciduous tree or shrub | |
| Prunus polytricha Koehne in Sarg. | macrophanerophytes or shrub | |
| Viburnum erubescens Wall. var. gracilipes Rehd. | macrophanerophytes or shrub | |
| Sambucus williamsii Hance | shrub | |
| Prunus clarofolia C. K. Schneid. | macrophanerophytes or shrub | |
| Forsythia suspensa (Thunb.) Vahl | shrub | |
| Philadelphus incanus Koehne | shrub | |
| Rhus potaninii Maxim. | macrophanerophytes | |
| Dipelta floribunda Maxim. | deciduous tree or shrub | |
| Morus alba L. | macrophanerophytes or shrub | |
| Rhus punjabensis var. sinica (Diels) Rehd.et Wils. | macrophanerophytes | |
| Malus baccata (L.) Borkh. | macrophanerophytes | |
| Kalopanax septemlobus (Thunb.) Koidz. | macrophanerophytes | |
| Lindera aggregata (Sims) Kosterm var. playfairii (Hemsl.) H. P. Tsui | macrophanerophytes or shrub | |
| Corylus heterophylla var. sutchuenensi Franch. | macrophanerophytes or shrub | |
| Cotoneaster acuminatus Lindl. | shrub | |
| Picrasma quassioides (D. Don) Benn. | macrophanerophytes | |
| Quercus aliena var. acutiserrata Maximowicz ex Wenzig | macrophanerophytes | |
| Fraxinus platypoda Oliv. | Late successional species group | macrophanerophytes |
| Acer mono Maxim. | macrophanerophytes | |
| Tilia oliveri | macrophanerophytes | |
| Toxicodendron vernicifluum (Stokes) F. A. Barkley | macrophanerophytes | |
| Quercus wutaishansea Mary | macrophanerophytes | |
| Ulmus bergmanniana C. K. Schneid. | macrophanerophytes | |
| Tilia oliveri Szyszyl. | macrophanerophytes | |
| Pinus armandii Franch. | macrophanerophytes | |
| Tilia chinensis Maxim. | macrophanerophytes | |
| Ulmus davidiana Planch. var. japonica(Rehd.) Nakai | macrophanerophytes | |
| Sorbus koehneana C. K. Schneid. | macrophanerophytes or shrub | |
| Betula platyphylla Sukaczev | Pioneer Species Group | macrophanerophytes |
| Populus purdomii Rehder | macrophanerophytes | |
| Lindera obtusiloba Blume | shrub |
References
- Condit, R. Research in large, long-term tropical forest plots. Trends Ecol. Evol. 1995, 10, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Mitsuishi, D.; Uemura, S.; Suzuki, J.-I.; Hara, T. Stand structure and dynamics during a 16-year period in a sub-boreal conifer–hardwood mixed forest, northern Japan. For. Ecol. Manag. 2003, 174, 39–50. [Google Scholar] [CrossRef]
- Ke-Ping, M.A. Large Scale Permanent Plots: Important Platform for Long Term Research on Biodiversity in Forest Ecosystem. Chin. J. Plant Ecol. 2008, 32, 237. [Google Scholar]
- Yamada, T.; Aiba, S.I.; Kubota, Y.; Okubo, K.; Miyata, I.; Suzuki, E.; Maenaka, H.; Nagano, M. Dynamics of species diversity in a Japanese warm-temperate secondary forest. Ecosphere 2011, 2, 1–19. [Google Scholar] [CrossRef]
- Feeley, K.J.; Davies, S.J.; Perez, R.; Hubbell, S.P.; Foster, R.B. Directional changes in the species composition of a tropical forest. Ecology 2011, 92, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Gaines, S.D.; Denny, M.W. The largest, smallest, highest, lowest, longest, and shortest: Extremes in ecology. Ecology 1993, 74, 1677. [Google Scholar] [CrossRef]
- Sheil, D.; Jennings, S.; Savill, P. Long-term permanent plot observations of vegetation dynamics in Budongo, a Ugandan rain forest. J. Trop. Ecol. 2000, 16, 865–882. [Google Scholar] [CrossRef]
- Condit, R.; Hubbell, S.P.; Foster, R.B. Short-Term Dynamics of a Neotropical Forest. Bioscience 1992, 42, 822–828. [Google Scholar] [CrossRef]
- Condit, R. Ecological Implications of Changes in Drought Patterns: Shifts in Forest Composition in Panama. Clim. Chang. 1998, 39, 413–427. [Google Scholar] [CrossRef]
- Peng, S.; Fang, W.; Ren, H.; Huang, Z.; Zhang, D. The Dynamics on Organization in the Successional Process of Dinghushan Cryptocarya Community. Chin. J. Plant Ecol. 1998, 22, 245. [Google Scholar]
- Rees, M.; Condit, R.; Crawley, M.; Pacala, S.; Tilman, D. Long-term studies of vegetation dynamics. Science 2001, 293, 650–655. [Google Scholar] [CrossRef]
- Laurance, W.F.; Oliveira, A.A.; Laurance, S.G.; Condit, R.; Nascimento, H.E.M.; Sanchez-Thorin, A.C.; Lovejoy, T.E.; Andrade, A.; D’Angelo, S.; Ribeiro, J.E. Pervasive alteration of tree communities in undisturbed Amazonian forests. Nature 2004, 428, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.L.; Phillips, O.L.; Sheil, D.; Vinceti, B.; Baker, T.R.; Brown, S.; Graham, A.W.; Higuchi, N.; Hilbert, D.W.; Laurance, W.F. Tropical forest tree mortality, recruitment and turnover rates: Calculation, interpretation and comparison when census intervals vary. J. Ecol. 2004, 92, 929–944. [Google Scholar] [CrossRef]
- Weckel, M.; Tirpak, J.M.; Nagy, C.; Christie, R. Structural and compositional change in an old-growth eastern hemlock Tsuga canadensis forest, 1965–2004. For. Ecol. Manag. 2006, 231, 114–118. [Google Scholar] [CrossRef]
- Fauset, S.; Baker, T.R.; Lewis, S.L.; Feldpausch, T.R.; Affum-Baffoe, K.; Foli, E.G.; Hamer, K.C.; Swaine, M.D. Drought-induced shifts in the floristic and functional composition of tropical forests in Ghana. Ecol. Lett. 2012, 15, 1120–1129. [Google Scholar] [CrossRef] [PubMed]
- Holzmueller, E.J.; Gibson, D.J.; Suchecki, P.F. Accelerated succession following an intense wind storm in an oak-dominated forest. For. Ecol. Manag. 2012, 279, 141–146. [Google Scholar] [CrossRef]
- Marimon, B.S.; Felfili, J.M.; Fagg, C.W.; Marimon-Junior, B.H.; Umetsu, R.K.; Oliveira-Santos, C.; Morandi, P.S.; Lima, H.S.; Nascimento, A.R.T. Monodominance in a forest of Brosimum rubescens Taub. (Moraceae): Structure and dynamics of natural regeneration. Acta Oecologica 2012, 43, 134–139. [Google Scholar] [CrossRef]
- Ge, J.; Xiong, G.; Zhao, C.; Shen, G.; Xie, Z. Short-term dynamic shifts in woody plants in a montane mixed evergreen and deciduous broadleaved forest in central China. For. Ecol. Manag. 2013, 310, 740–746. [Google Scholar] [CrossRef]
- Shen, Y.; Santiago, L.S.; Ma, L.; Lin, G.J.; Lian, J.Y.; Cao, H.L.; Ye, W.H. Forest dynamics of a subtropical monsoon forest in Dinghushan, China: Recruitment, mortality and the pace of community change. J. Trop. Ecol. 2013, 29, 131–145. [Google Scholar] [CrossRef]
- Zhao, Z. Forest Naturalness Evaluation Method Based on Stand State Characters:A Case Study of Gansu Xiaolongshan Forests. Sci. Silvae Sin. 2011, 47, 9–16. [Google Scholar]
- Zhao, Z.H.; Hui, G.Y.; Yuan, S.Y.; Liu, W.Z. Tree Species Diversity and Structure Characteristics of Quercus aliena var. acuteserrata Natural Forest on Xiaolongshan. For. Res. 2008, 21, 605–610. [Google Scholar]
- Zhao, Z. Spatial Structure Characteristic of Quercus aliena var. acuteserrata Natural Forest in Xiaolongshan. Sci. Silvae Sin. 2009, 45, 1–6. [Google Scholar]
- Ma, Y.; Liu, W.; Zhao, Z.; Shi, X.; Li, A. Change Analysis of Spatial Pattern and Correlation for Quercus aliena var. acuteserrata Forest Population in Xiaolong Mountains. Acta Bot. Boreali-Occident. Sin. 2014, 34, 1878–1886. [Google Scholar]
- Mooney, H.A. Plants in Changing Environments: Linking Physiological, Population and Community Ecology. Tree Physiol. 1997, 17, 489. [Google Scholar] [CrossRef]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; Updated Edition; John Wiley & Sons: Hoboken, NJ, USA, 1996. [Google Scholar]
- Allen, C.D.; Breshears, D.D. Drought-induced shift of a forest–woodland ecotone: Rapid landscape response to climate variation. Proc. Natl. Acad. Sci. USA 1998, 95, 14839–14842. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.B.; Clark, D.A.; Oberbauer, S.F. Annual wood production in a tropical rain forest in NE Costa Rica linked to climatic variation but not to increasing CO2. Glob. Chang. Biol. 2010, 16, 747–759. [Google Scholar] [CrossRef]
- Feeley, K.J.; Wright, S.J.; Supardi, M.N.N.; Kassim, A.R.; Davies, S.J. Decelerating growth in tropical forest trees. Ecol. Lett. 2007, 10, 461–469. [Google Scholar] [CrossRef]
- Phillips, O.L.; Baker, T.R.; Arroyo, L.; Higuchi, N.; Vinceti, B. Pattern and process in Amazon tree turnover, 1976–2001. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 381–407. [Google Scholar] [CrossRef]
- Breshears, D.D.; Myers, O.B.; Meyer, C.W.; Barnes, F.J.; Zou, C.B.; Allen, C.D.; McDowell, N.G.; Pockman, W.T. Tree die-off in response to global change-type drought: Mortality insights from a decade of plant water potential measurements. Front. Ecol. Environ. 2009, 7, 185–189. [Google Scholar] [CrossRef]
- van Mantgem, P.J.; Stephenson, N.L.; Byrne, J.C.; Daniels, L.D.; Franklin, J.F.; Fule, P.Z.; Harmon, M.E.; Larson, A.J.; Smith, J.M.; Taylor, A.H.; et al. Widespread increase of tree mortality rates in the western United States. Science 2009, 323, 521–524. [Google Scholar] [CrossRef]
- Peng, C.; Ma, Z.; Lei, X.; Zhu, Q.; Chen, H.; Wang, W.; Liu, S.; Li, W.; Fang, X.; Zhou, X. A drought-induced pervasive increase in tree mortality across Canada’s boreal forests. Nat. Clim. Chang. 2011, 1, 467–471. [Google Scholar] [CrossRef]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J. Regional vegetation die-off in response to global-change-type drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef] [PubMed]
- Woods, K.D. Dynamics in late-successional hemlock–hardwood forests over three decades. Ecology 2000, 81, 110–126. [Google Scholar]
- Wang, M.; Shu, L.; Wang, Q.; Zhao, F.; Tian, X.; Yan, H.; Du, J. Forest fire occurrence in short term under the impacts of snow damage in south China—Case study in Hunan Province. Sci. Silvae Sin 2008, 44, 64–68. [Google Scholar]
- Cao, K.-F.; Chang, J. The ecological effects of an unusual climatic disaster: The destruction to forest ecosystems by the extremely heavy glaze and snow storms occurred in early 2008 in southern China. Chin. J. Plant Ecol. 2010, 34, 123. [Google Scholar]
- Chen, Y.; Niu, S.; Zhao, X.; He, J.; Fan, J.; Mao, S. Damage and recovery of a evergreen broadleaved forest in Jiulianshan National Nature Reserve after affected by freezing rain. J. Northeast. For. Univ. 2011, 39, 20–34. [Google Scholar]
- Su, Z.; Liu, G.; Ou, Y.; Li, Z. Storm damage in a montane evergreen broadleaved forest of Chebaling National Nature Reserve, South China. Chin. J. Plant Ecol. 2010, 34, 213–222. (In Chinese) [Google Scholar]
- Wang, J.; Wen, X.F.; Wang, H.M.; Wang, J.Y. The effects of ice storms on net primary productivity in a subtropical coniferous plantation. Acta Ecol. Sin. 2014, 34, 5030–5039. [Google Scholar]
- Ning, S.A.; Zhen, J.T.; Hua, Z.J.; Hua, W.Q. Analysis of biodiversity characteristics of Quercus aliena var. acuteserrata community on Mt.Xiaolong in Gansu. Acta Bot. Boreali-Occident. Sin. 2004, 24, 1877–1881. [Google Scholar]
- Anning, S.; Tianzhen, J.U.; Junhua, Z.; Jianping, G.E. Biomass structure of Quercus aliena var. acuteserrata community on Mt.Xiaolongshan in Gansu. Chin. J. Ecol. 2005, 4, 377. [Google Scholar]
- Editorial Committee of English Version; Li, L.; Yang, Q.; Li, F.Y. Vegetation of China; China Agricultural University Press: Beijing, China, 2020. [Google Scholar]
- Zhang, G.; Wang, D.; Zhang, M.; Liu, W.; Guo, X. Interspecific Association and Correlation of Main Species in the Succession Process of Pine Oak Forest Community on the South-facing Slopes in Qinling Mountain. Acta Bot. Boreali-Occident. Sin. 2015, 35, 1657–1668. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1963. [Google Scholar]
- Pielou, E.C. Ecological Diversity. A Wiley Interscience Publication; John Wiley & Sons: New York, NY, USA; London, UK; Sydney, Australia; Toronto, ON, Canada, 1975. [Google Scholar]
- Lévesque, M.; McLaren, K.P.; McDonald, M.A. Recovery and dynamics of a primary tropical dry forest in Jamaica, 10 years after human disturbance. For. Ecol. Manag. 2011, 262, 817–826. [Google Scholar] [CrossRef]
- Cascante-Marín, A.; Meza-Picado, V.; Estrada-Chavarría, A. Tree turnover in a premontane neotropical forest (1998–2009) in Costa Rica. Plant Ecol. 2011, 212, 1101–1108. [Google Scholar] [CrossRef]
- Kolmogorov, A. Sulla determinazione empirica di una lgge di distribuzione. Inst. Ital. Attuari Giorn. 1933, 4, 83–91. [Google Scholar]
- Smirnoff, N.W. On the estimation of the discrepancy between empirical curves of distribution for two independent samples. Bull. Math. Univ. Moscou 1939, 2, 3–14. [Google Scholar]
- Van Lieshout, M.N.M. Book Review: Statistical Analysis and Modelling of Spatial Point Patterns. By J. Illian, A. Penttinen, H. Stoyan, and D. Stoyan. Biom. J. 2008, 50, 627. [Google Scholar] [CrossRef]
- Pommerening, A.; Stoyan, D. Edge-correction needs in estimating indices of spatial forest structure. Can. J. For. Res. 2006, 36, 1723–1739. [Google Scholar] [CrossRef]
- Wickham, H.; Chang, W. Package ggplot2: Create Elegant Data Visualisations Using the Grammar of Graphics; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Enquist, B.J.; Enquist, C.A. Long-term change within a Neotropical forest: Assessing differential functional and floristic responses to disturbance and drought. Glob. Chang. Biol. 2011, 17, 1408–1424. [Google Scholar] [CrossRef]
- Mori, A.S.; Mizumachi, E.; Komiyama, A. Roles of disturbance and demographic non-equilibrium in species coexistence, inferred from 25-year dynamics of a late-successional old-growth subalpine forest. For. Ecol. Manag. 2007, 241, 74–83. [Google Scholar] [CrossRef]
- Lewis, S.L.; Lloyd, J.; Sitch, S.; Mitchard, E.T.A.; Laurance, W.F. Changing Ecology of Tropical Forests: Evidence and Drivers. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 529–549. [Google Scholar] [CrossRef]





| Plot | Census Time/Year | Density/Trees·ha−1 | Tree Species Number | Tree Species Composition |
|---|---|---|---|---|
| Plot A | 2007 | 1356 | 49 | 4 Quercus aliena 1 Acer davidii. 5 Other |
| 2012 | 1247 | 47 | 4 Quercus aliena 1 Rhusverniciflus 5 Other | |
| 2017 | 1178 | 46 | 4 Quercus aliena 1 Rhusverniciflus 5 Other | |
| Plot B | 2009 | 1276 | 49 | 4 Quercus aliena 1 Carpinus turczaninowii 5 Other |
| 2014 | 1265 | 50 | 4 Quercus aliena 1 Carpinus turczaninowii 5 Other | |
| 2019 | 1275 | 48 | 4 Quercus aliena 1 Carpinus turczaninowii 5 Other | |
| Plot C | 2009 | 1009 | 48 | 3 Quercus aliena 1 Carpinus turczaninowii 1 Sorbus alnifolia 1 Carpinus stipulata 4 Other |
| 2014 | 1068 | 50 | 3 Quercus aliena 1 Carpinus turczaninowii 1 Sorbus alnifolia 1 Carpinus stipulata 4 Other | |
| 2019 | 1085 | 51 | 3 Quercus aliena 1 Carpinus turczaninowii 1 Sorbus alnifolia 1 Carpinus stipulata 4 Other |
| Plot | Census Time/Year | Basal Area/m2·ha−1 | Canopy Closure | Mean DBH/ cm | Mean Height/m | Diversity and Evenness | |||
|---|---|---|---|---|---|---|---|---|---|
| Shannon–Wiener (Stem) | Shannon–Wiener (Basal Area) | Pielou (Stem) | Pielou (Basal Area) | ||||||
| Plot A | 2007 | 22.786 | 0.80 | 14.6 ± 8.0 | 11.9 | 2.911 | 2.571 | 0.748 | 0.661 |
| 2012 | 24.861 | 0.80 | 15.9 ± 8.2 | 12.5 | 2.901 | 2.492 | 0.754 | 0.647 | |
| 2017 | 26.500 | 0.85 | 16.9 ± 8.8 | 13.1 | 2.901 | 2.480 | 0.758 | 0.648 | |
| Plot B | 2009 | 34.337 | 0.85 | 18.5 ± 10.4 | 12.7 | 3.069 | 2.370 | 0.788 | 0.609 |
| 2014 | 37.244 | 0.85 | 19.4 ± 11.0 | 13.3 | 3.060 | 2.340 | 0.782 | 0.598 | |
| 2019 | 39.795 | 0.90 | 19.9 ± 11.3 | 13.6 | 3.057 | 2.308 | 0.790 | 0.596 | |
| Plot C | 2009 | 18.195 | 0.80 | 15.2 ± 8.3 | 11.2 | 3.135 | 2.816 | 0.810 | 0.727 |
| 2014 | 21.230 | 0.80 | 15.9 ± 8.7 | 10.9 | 3.147 | 2.844 | 0.804 | 0.727 | |
| 2019 | 23.559 | 0.85 | 16.6 ± 9.1 | 11.0 | 3.152 | 2.847 | 0.802 | 0.724 | |
| Plot | Census Time/Year | Living Trees | Dead Trees | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Density/ Trees·ha−1 | Mean DBH/cm | Mean Height/m | Tree Species Number | Density/ Trees·ha−1 | Mean DBH/cm | Mean Height/m | Tree Species Number | ||
| Plot A | 2007 | 1356 | 14.6 b | 11.9 b | 49 | - | - | - | - |
| 2012 | 1247 | 15.9 b | 12.5 b | 47 | 111 | 13.1 | 9.7 | 16 | |
| 2017 | 1178 | 16.9 b | 13.1 b | 46 | 89 | 11.3 | 10.6 | 14 | |
| Plot B | 2009 | 1276 | 18.5 a | 12.7 a | 49 | - | - | - | - |
| 2014 | 1265 | 19.4 a | 13.3 a | 50 | 66 | 10.6 | 9.3 | 24 | |
| 2019 | 1275 | 19.9 a | 13.6 a | 48 | 30 | 10.4 | 9.8 | 15 | |
| Plot C | 2009 | 1009 | 15.2 b | 11.2 b | 48 | - | - | - | - |
| 2014 | 1068 | 15.9 b | 10.9 b | 50 | 58 | 8.4 | 8.3 | 19 | |
| 2019 | 1085 | 16.6 b | 11.0 b | 51 | 27 | 10.7 | 9.3 | 11 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lian, Q.; Sa, R.; Liu, W.; Zhao, Z. Structural and Successional Dynamics of Old-Growth Mixed Oak Forests in the West Qinling Mountains. Forests 2024, 15, 765. https://doi.org/10.3390/f15050765
Lian Q, Sa R, Liu W, Zhao Z. Structural and Successional Dynamics of Old-Growth Mixed Oak Forests in the West Qinling Mountains. Forests. 2024; 15(5):765. https://doi.org/10.3390/f15050765
Chicago/Turabian StyleLian, Qi, Rina Sa, Wenzhen Liu, and Zhonghua Zhao. 2024. "Structural and Successional Dynamics of Old-Growth Mixed Oak Forests in the West Qinling Mountains" Forests 15, no. 5: 765. https://doi.org/10.3390/f15050765
APA StyleLian, Q., Sa, R., Liu, W., & Zhao, Z. (2024). Structural and Successional Dynamics of Old-Growth Mixed Oak Forests in the West Qinling Mountains. Forests, 15(5), 765. https://doi.org/10.3390/f15050765