Response of the Invasive Grass Imperata cylindrica to Disturbance in the Southeastern Forests, USA


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
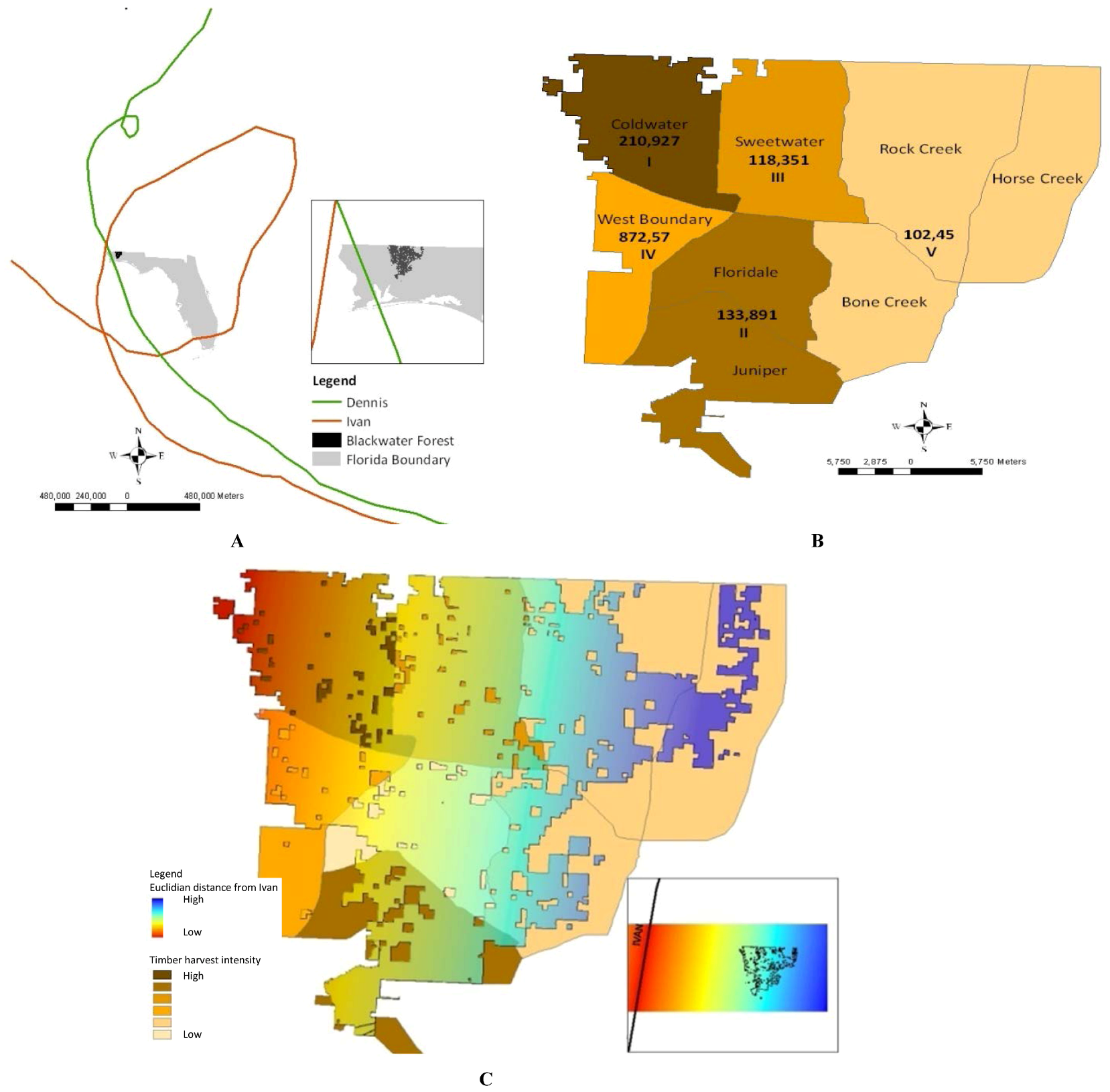
2. Experimental Section


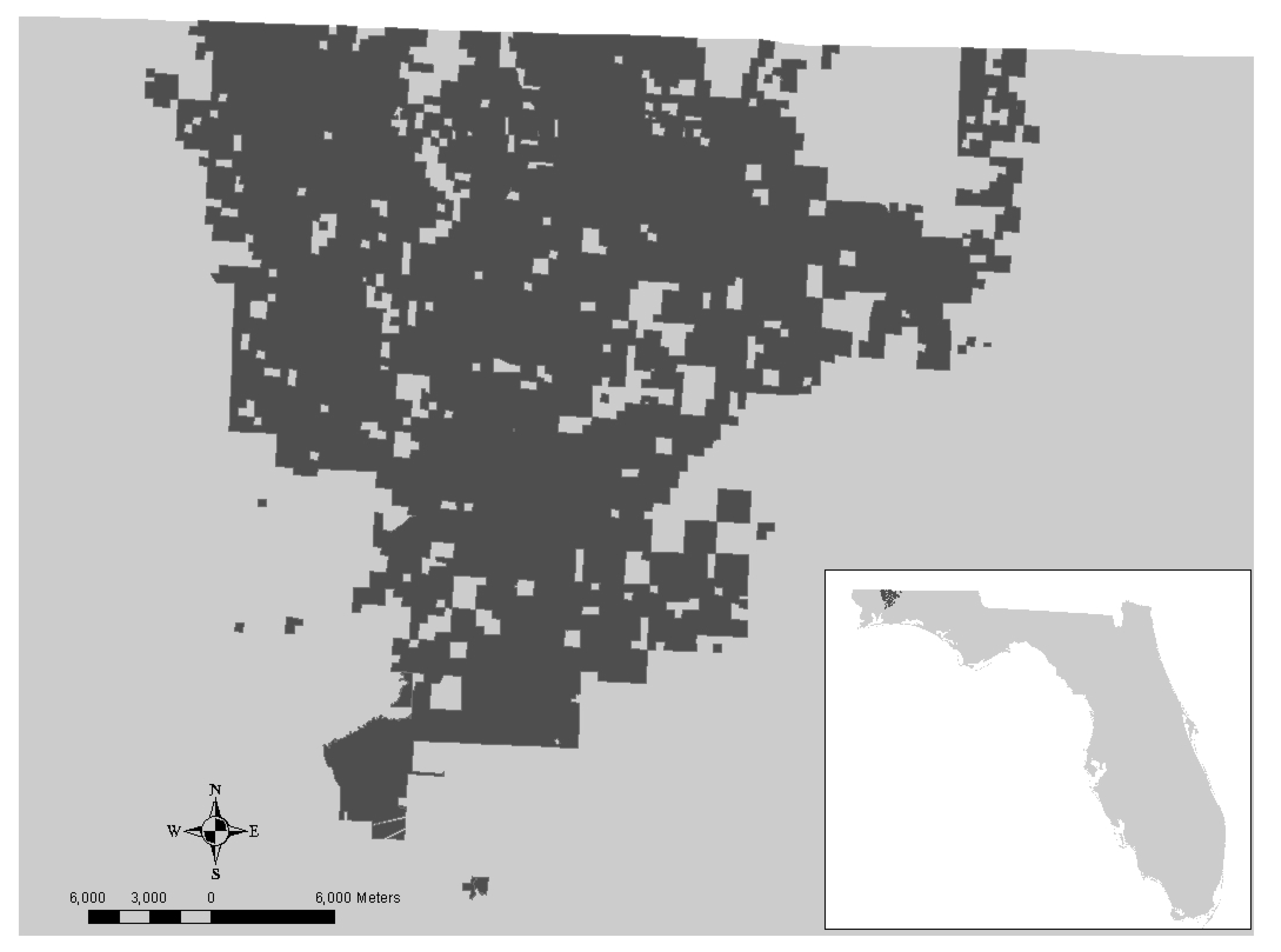
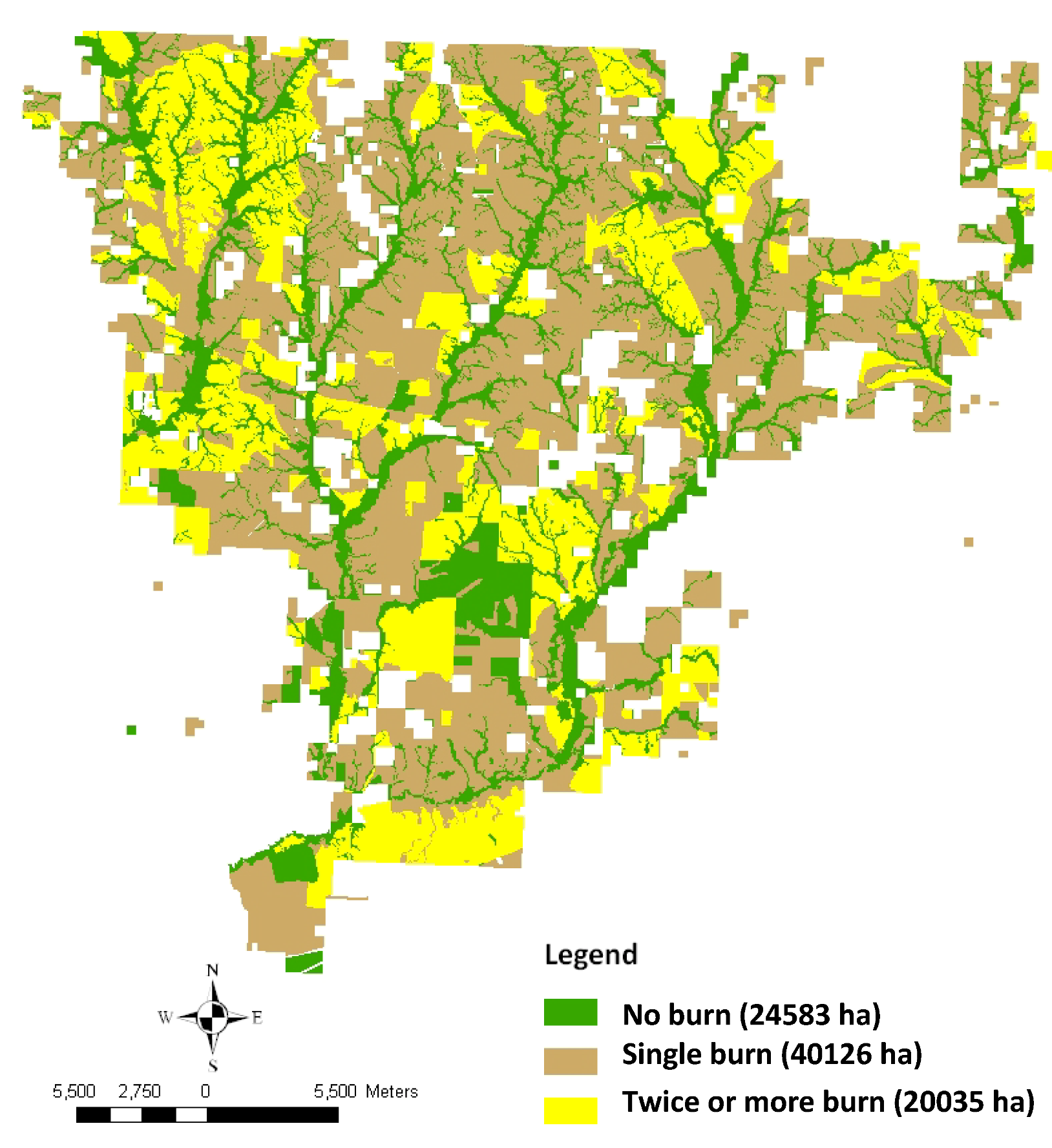
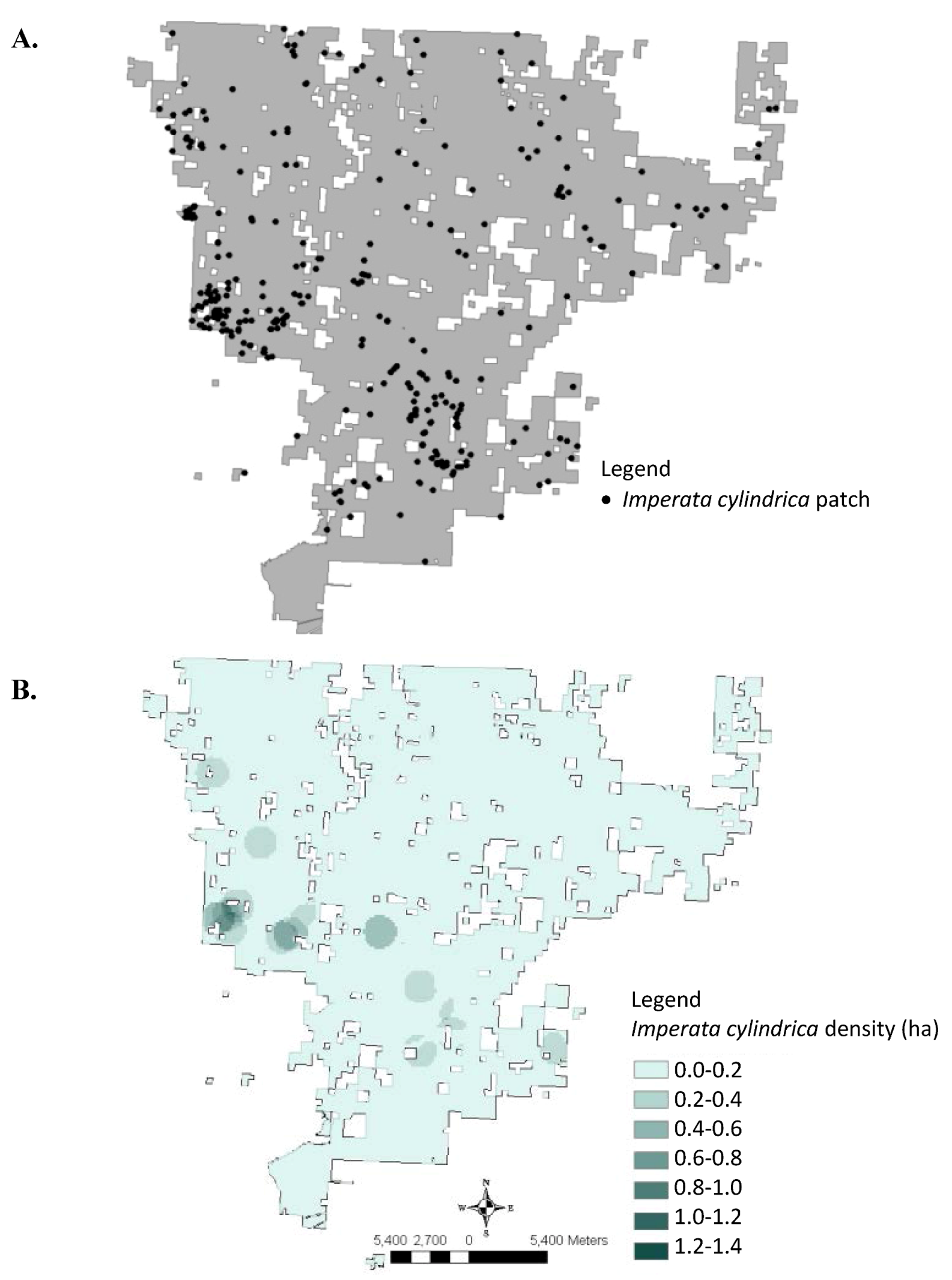
3. Results and Discussion




4. Conclusions
Acknowledgments
Conflict of Interest
References
- MacDonald, G.E. Cogongrass: The Plant’s Biology, Distribution, and Impacts in the Southeastern US. In Proceedings of the Regional Cogongrass Conference: A Cogongrass Management Guide; Loewenstein, N.J., Miller, J.H., Eds.; Alabama Cooperative Extension System, Auburn University: Mobile, AL, USA, 2007; pp. 10–23. [Google Scholar]
- Dozier, H.; Gaffney, J.F.; McDonald, S.K.; Johnson, E.R.R.L.; Shilling, D.G. Cogongrass in the United States. History, ecology, impacts, and management. Weed Technol. 1998, 12, 737–743. [Google Scholar]
- MacDonald, G.E. Cogongrass (Imperata cylindrica)—Biology, ecology and management. Crit. Rev. Plant Sci. 2004, 23, 367–380. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjela, S.; de Poorter, M. 100 of the World’s Worst Invasive Alien Species A Selection from the Global Invasive Species Database; The Invasive Species Specialist Group (ISSG): Auckland, New Zealand, 2004; p. 12. [Google Scholar]
- Brewer, S. Declines in plant species richness and endemic plant species in longleaf pine savannas invaded by Imperata cylindrica. Biol. Invasions 2008, 10, 1257–1264. [Google Scholar] [CrossRef]
- Collins, A.R.; Jose, S. Cogongrass Invasion Alters Soil Chemical Properties of Natural and Planted Forestlands. In Invasive Plants and Forest Ecosystems; Kohli, R.K., Jose, S., Batish, D., Singh, H., Eds.; CRC: Boca Raton, FL, USA, 2008. [Google Scholar]
- Daneshgar, P.; Jose, S. Imperata cylindrica, an alien invasive grass, maintains control over nitrogen availability in an establishing pine forest. Plant Soil 2009, 320, 209–218. [Google Scholar] [CrossRef]
- Daneshgar, P.; Jose, S.; Ramsey, C.; Collins, A.R. Impacts of an invasive grass on the productivity of an establishing pine forest. Forest Sci. 2008, 54, 579–587. [Google Scholar]
- Willard, T.R.; Hall, D.W.; Shilling, D.G.; Lewis, J.A.; Currey, W.L. Cogongrass (Imperata cylindrica) distribution on Florida highway rights-of-way. Weed Technol. 1990, 4, 658–660. [Google Scholar]
- King, S.E.; Grace, J.B. The effects of gap size and disturbance type on invasion of wet pine savanna by cogongrass, Imperata cylindrica (Poaceae). Am. J. Bot. 2000, 87, 1279–1286. [Google Scholar] [CrossRef]
- Ayeni, A.O.; Duke, W.B. The influence of rhizome features on subsequent regenerative capacity in speargrass [Imperata cylindrica (L.) Veauv.]. Agric. Ecosyst. Environ. 1985, 13, 309–317. [Google Scholar] [CrossRef]
- Holzmueller, E.J.; Jose, S. Invasion success of cogongrass, an alien C4 perennial grass, in the southeastern United States: Exploration of the ecological basis. Biol. Invasions 2011, 13, 435–442. [Google Scholar] [CrossRef]
- Yager, L.Y.; Jones, J.; Miller, D.L. Military training and road effects on Imperata cylindrica (L.) Beauv. (Cogongrass). Southeast. Nat. 2009, 8, 695–708. [Google Scholar] [CrossRef]
- Patterson, M.; Teem, D.; Faircloth, W. Mapping, Control, and Revegetation of Cogongrass infestations on Alabama Rights of Way. ALDOT Research Project 930–486; Alabama Department of Transportation and Auburn University Alabama Agricultural Experiment Station: Auburn, AL, USA, 2004. [Google Scholar]
- Jose, S.; Cox, J.; Miller, D.L.; Shilling, D.G.; Merritt, S. Alien plant invasions: The story of cogongrass in southeastern Florida. J. For. 2002, 100, 41–44. [Google Scholar]
- Lippincott, C.L. Effects of Imperata cylindrica (L.) Beauv. (Cogongrass) invasion on fire regime in Florida sandhill (USA). Nat. Area J. 2000, 20, 140–149. [Google Scholar]
- Yager, L.Y.; Miller, D.L.; Jones, J. Susceptibility of longleaf pine forest associations in south Mississippi to invasion by Cogongrass [Imperata cylindrica (L.) Beauv.]. Nat. Area J. 2010, 30, 226–232. [Google Scholar] [CrossRef]
- Mortensen, D.A.; Rauschert, E.S.J.; Nord, A.N.; Jones, B.P. Forest roads facilitate the spread of invasive plants. Invasive Plant Sci. Manag. 2009, 2, 191–199. [Google Scholar]
- Glitzenstein, J.S.; Platt, W.J.; Streng, D.R. Effects of fire regime and habitat on tree dynamics in north Florida longleaf pine savannas. Ecol. Monogr. 1995, 65, 441–476. [Google Scholar]
- D’Antonio, C.M.; Vitousek, P.M. Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annu. Rev. Ecol. Syst. 1992, 23, 63–87. [Google Scholar]
- Brooks, M.L.; D’Antonio, C.M.; Richardson, D.M.; Grace, J.B.; Keeley, J.E.; Ditomaso, J.M.; Hobbs, R.J.; Pellant, M.; Pyke, D. Effects of invasive alien plants on fire regimes. BioScience 2004, 54, 677–688. [Google Scholar]
- Ramsey, C.L.; Jose, S.; Miller, D.L.; Cox, J.; Portier, K.M.; Shilling, D.G.; Merritt, S.M. Cogongrass [Imperata cylindrica (L.) Beauv.] response to herbicide and disking on a cutover site in a mid-rotation pine plantation in Southern USA. For. Ecol. Manag. 2003, 179, 195–209. [Google Scholar] [CrossRef]
- Marshall, J.M.; Buckley, D.S. Effects of microsites created by selective harvesting on growth of Microstegium vimineum in a central hardwood forest. For. Sci. 2008, 54, 534–542. [Google Scholar]
- Veldman, J.W.; Mostacedo, B.; Pena-Claros, M.; Putz, F.E. Selective logging and fire as drivers of alien grass invasion in a Bolivian tropical dry forest. For. Ecol. Manag. 2009, 258, 1643–1649. [Google Scholar]
- Brewer, J.S.; Bertz, C.A.; Cannon, J.B.; Chesser, J.D.; Maynard, E.E. Do natural disturbances or the forestry practices that follow them convert forests to early-successional communities? Ecol. Appl. 2012, 22, 442–458. [Google Scholar] [CrossRef]
- Holzmueller, E.J.; Jose, S. Response of cogongrass (Imperata cylindrica) to imazapyr herbicides on a reclaimed phosphate-mine site in central Florida, USA. Ecol. Restor. 2010, 28, 300–303. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Holzmueller, E.J.; Jose, S. Response of the Invasive Grass Imperata cylindrica to Disturbance in the Southeastern Forests, USA. Forests 2012, 3, 853-863. https://doi.org/10.3390/f3040853
Holzmueller EJ, Jose S. Response of the Invasive Grass Imperata cylindrica to Disturbance in the Southeastern Forests, USA. Forests. 2012; 3(4):853-863. https://doi.org/10.3390/f3040853
Chicago/Turabian StyleHolzmueller, Eric J., and Shibu Jose. 2012. "Response of the Invasive Grass Imperata cylindrica to Disturbance in the Southeastern Forests, USA" Forests 3, no. 4: 853-863. https://doi.org/10.3390/f3040853
APA StyleHolzmueller, E. J., & Jose, S. (2012). Response of the Invasive Grass Imperata cylindrica to Disturbance in the Southeastern Forests, USA. Forests, 3(4), 853-863. https://doi.org/10.3390/f3040853