Why Do Some Evergreen Species Keep Their Leaves for a Second Winter, While Others Lose Them?

Abstract
:1. Introduction

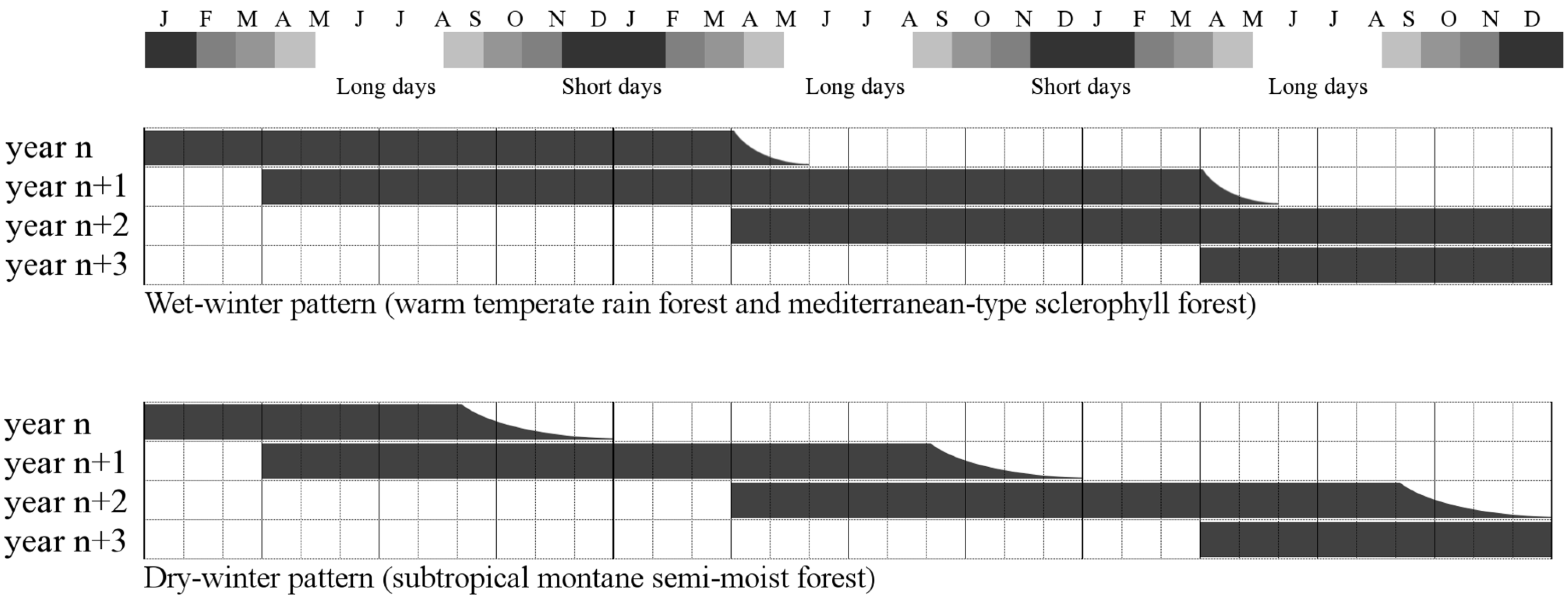{kind=link}
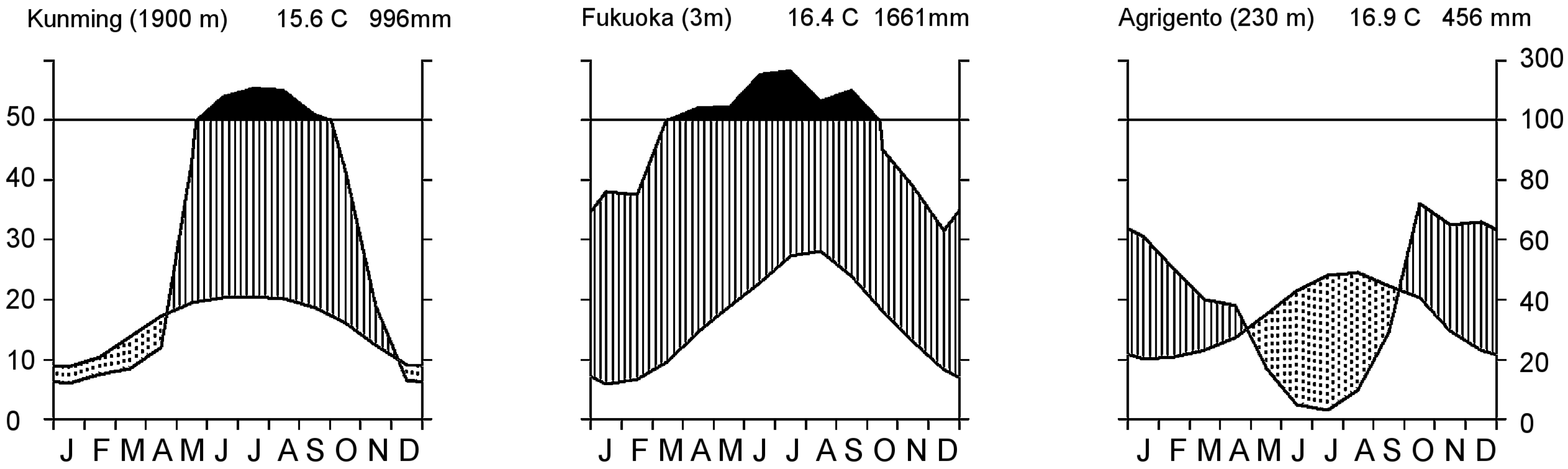{kind=link}
{kind=link}
| Species | Family | Months when new leaves grew out | Months in which leaf fall was concentrated |
|---|---|---|---|
| WTRF Myrsine neriifolia | Primulaceae | April–May | April–June and September–December |
| Neolitsea sericea | Lauraceae | April–May | May–December |
| Cleyera japonica | Pentaphylacaceae | April–May | July–October |
| Symplocos prunifolia | Symplocaceae | April–May | Continuous |
| Quercus salicina | Fagaceae | May | March–May |
| Persea thunbergii | Lauraceae | May | May–September |
| Cinnamomum japonicum | Lauraceae | May | May–October (1995) |
| May–June, August– September and December–February (1994–1995) | |||
| Quercus acuta | Fagaceae | May–June | May–November |
| Illicium religiosum | Illiciaceae | May–June | June–December |
| Eurya japonica | Pentaphylacaceae | May–September and December–February | May–September and November–February |
| MSF | |||
| Myrtus communis | Myrtaceae | March–June | May–June |
| Pistacia lentiscus | Anacardiaceae | March–June | Not recorded * |
| Viburnum tinus | Adoxaceae | March–June | May–June |
| Pinus halepensis | Pinaceae | March–July | Peaks in spring |
| P. pinaster | Pinaceae | March–July | July–August |
| Buxus sempervirens | Buxaceae | March–August | May–July |
| Quercus ilex | Fagaceae | April–May (+ some waves to September) | May + September to December |
| Arbutus unedo | Ericaceae | April–June | May–June |
| Quercus suber | Fagaceae | April–June | June–August |
| Phillyrea angustifolia | Oleaceae | April–August | April–July |
| Quercus coccifera | Fagaceae | May (+ some August–September) | Mostly May–July |
| Rhamnus alaternus | Rhamnaceae | May–August | May–August |
| Species | Mean leaf longevity (year) determined in Greece | Mean leaf longevity (year) determined in Spain | Range of longevity (year) determined in France |
|---|---|---|---|
| Pinus halepensis | 3.1 | 2–3, most <2.5 | |
| P. pinea | 3.0 | ||
| P. pinaster | 2.1 | (2)–3+, mostly >3 | |
| Quercus ilex | 2.0 | 2–3 | |
| Q. coccifera | 1.1 | 1.3 | 2.1–2.3 |
| Arbutus unedo | 1.2 | 2 (–3) | |
| Phillyrea | |||
| angustifolia | 2.1–2.2 | ||
| Pistacia lentiscus | 0.94 | Not >2 | |
| Ceratonia siliqua | 2.0 | ||
| Olea europaea | 2.0 | ||
| Myrtus communis | 1.9 | 1.1–1.2 | |
| Buxus | |||
| sempervirens | 1–2 | ||
| Quercus suber | 1.3 | 1.1–1.3 | |
| Arbutus andrachne | 1.1 |


2. Material and Methods
2.1. Observations in Yunnan
2.2. Initial Survey in Scotland
2.3. Critical Observations on Leaf Fall in Scotland
| Species | Subgenus and subsection | Natural distribution * | Provenance | Mean number of leaves studied per shoot |
|---|---|---|---|---|
| R. decorum ssp. decorum | Hymenanthes: Fortunea | G, S, X, Y; M | 2 NW Yunnan 1 W Yunnan | 8.0 (reduced by shoots dying to 6.0) |
| R. pubescens | Pseudorhodorastrum: Scabrifolia | S, Y | 3 Yunnan | 9.4 |
| R. rubiginosum var. rubiginosum | Rhododendron: Heliolepida | S, X, Y; M | 1 NW Yunnan 2 SW Sichuan | 9.4 |
| R. wallichii | Hymenanthes: Campanulata | X; B, I, N, S | 1 Bengal 2 Nepal | 4.9 |
| R. wardii var. wardii | Hymenanthes: Campylocarpa | S, X, Y | 1 W Yunnan 2 SE Xizang | 5.1 |
3. Results
3.1. Observations on Cohort Number in Yunnan
| Family | Species | Site |
|---|---|---|
| One cohort | ||
| Araliaceae | Metapanax delavayi* | 2 |
| Ericaceae | Rhododendron spiciferum | 1 |
| R. cyanocarpum | 3 | |
| R. racemosum | 3 | |
| R. rubiginosum | 3 | |
| R. xanthostephanum | 3 | |
| Fagaceae | Castanopsis orthacantha* | 3 |
| Lithocarpus dealbatus* | 1 | |
| Castanopsis delavayi | 1 | |
| Lithocarpus variolosus | 2 | |
| Myricaceae | Myrica nana* | 2 |
| Oleaceae | Osmanthus yunnanensis | 1 |
| Pentaphylacaceae | Eurya cf. nitida* | 2 |
| Ternstroemia gymnanthera* | 2 | |
| Sabiaceae | Meliosma yunnanensis | 2 |
| Theaceae | Schima argentea | 2 |
| Schima sp.* | 1 | |
| Two cohorts | ||
| Buxaceae | Sarcococca hookeriana | 2 |
| Ericaceae | Craibiodendron yunnanense* | 1 |
| Pieris formosa | 1 | |
| Magnoliaceae | Michellia yunnanensis | 1 |
| Rosaceae | Laurocerasus cf. zippeliana | 1 |
| Three cohorts | ||
| Theaceae | Camellia pitardii | 2 |
3.2. Observations on Cohort Number in Scotland
| Species, subspecies and variety | Mean length of leaf blade (cm) | Usual lower altitudinal limit (m) | Usual upper altitudinal limit (m) | Provinces where recorded |
|---|---|---|---|---|
| Leaves lasting >3 years | ||||
| R. strigillosum | 12.0 | 1600 | 3800 | S, Y |
| Three cohorts of old leaves present in spring | ||||
| R. faucium | 8.0 | 2600 | 3400 | X |
| R. rieri (I) | 10.3 | 1700 | 1800 | S |
| R. insigne | 10.5 | 700 | 2000 | S |
| R. arizelum | 14.0 | 2500 | 4000 | X, Y |
| R. rex ssp. rex | 24.5 | 2300 | 3300 | S, Y |
| Two cohorts of old leaves present in spring | ||||
| R. haematodes ssp. haematodes | 5.1 | 3200 | 4000 | Y |
| R. dichroanthum ssp. apodectum | 5.5 | 2600 | 3600 | Y |
| R. sanguineum var. sanguineum | 5.9 | 2800 | 4300 | X, Y |
| R. neriiflorum ssp. neriiflorum | 6.5 | 2500 | 3600 | X, Y |
| R. piercei | 7.5 | 3900 | 4200 | X |
| R. sperabile (I) | 9.1 | 2600 | 4200 | Y |
| R. bureavi | 10.0 | 2800 | 4500 | S, Y |
| R. arboreum ssp. arboreum | 10.5 | 1500 | 3600 | X |
| R. argyrophyllum | 10.5 | 1600 | 2300 | G, S, Y |
| R. seminoides | 20.5 | 3500 | 3900 | X, Y |
| R. rex ssp. fictolacteum | 24.5 | 2900 | 4000 | S, X, Y |
| R. magnificum | 26.0 | 1800 | 2400 | X |
| R. praestans | 26.5 | 3100 | 4200 | X, Y |
| Only one cohort of old leaves | ||||
| present in spring | ||||
| R. pubescens (I) | 2.2 | 2000 | 2600 | S, Y |
| R. hemitrichotum (I) | 2.3 | 2200 | 4000 | S, Y |
| R. fletcherianum | 3.8 | 3400 | 3400 | X, Y |
| R. davidsonianum | 4.3 | 1500 | 2800 | S |
| R. searsiae | 4.3 | 2300 | 3300 | S |
| R. concinnum | 5.0 | 2300 | 3000 | G, S, Y |
| R. siderophyllum (I) | 5.0 | 1800 | 3000 | G, S, Y |
| R. thomsonii | 5.0 | 3000 | 4000 | X |
| R. haematodes ssp. chaetomallum | 5.1 | 3100 | 4000 | X, Y |
| R. rubiginosum var. rubiginosum (BI) | 5.8 | 2800 | 3600 | S, X, Y |
| R. cerasinum | 6.5 | 3200 | 3800 | X |
| R. wardii (I) | 6.5 | 3000 | 4600 | S, X, Y |
| R. heliolepis | 8.5 | 3000 | 3700 | S, X, Y |
| R. neriiflorum ssp. appropinquans | 9.5 | 2100 | 3600 | X, Y |
| R. wallichii (BI) | 9.5 | 3000 | 4300 | X |
| R. sidereum | 15.0 | 2400 | 3400 | Y |
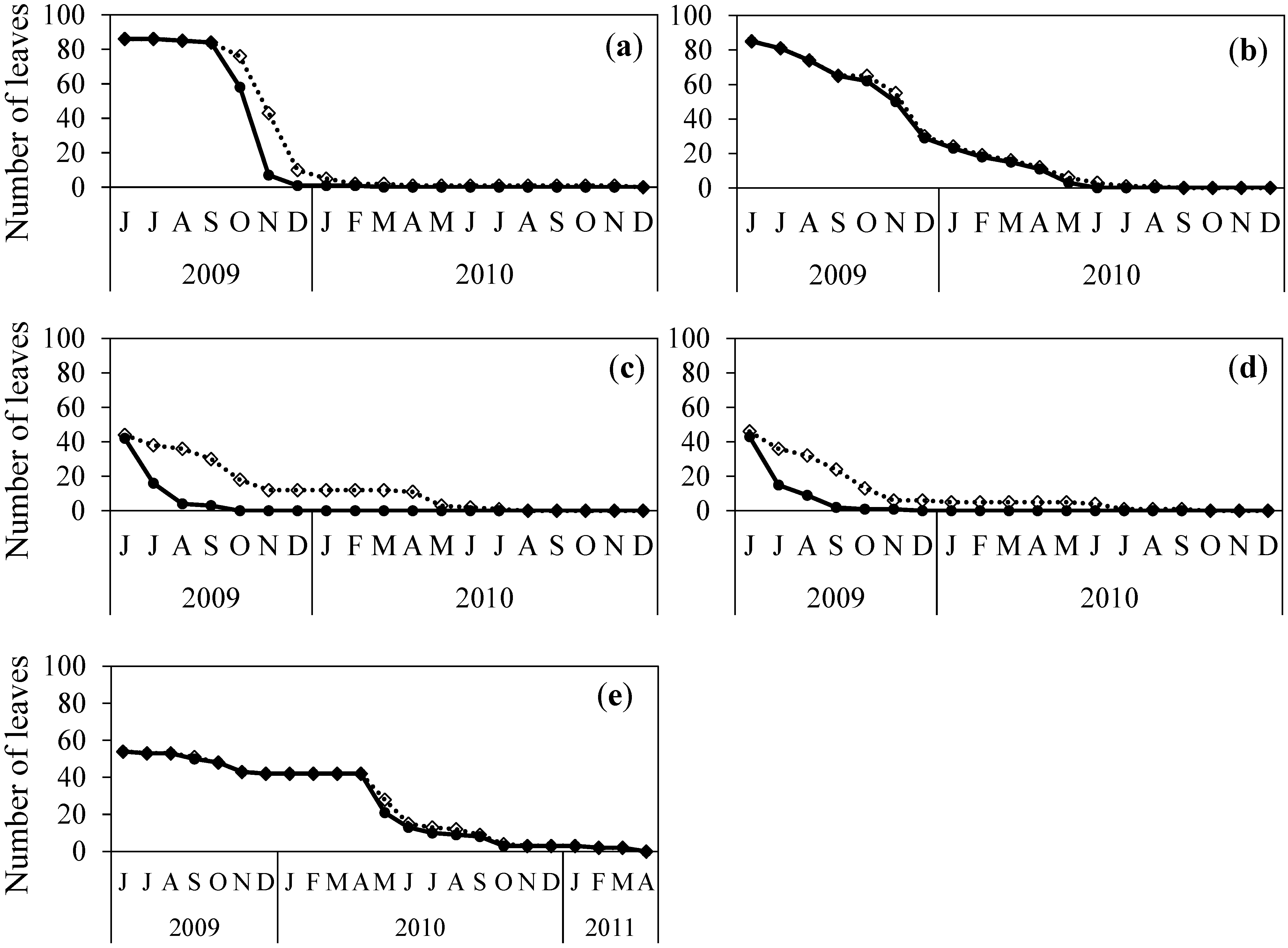
3.3. Observations on the Timing of Leaf Fall and Leaf Senescence in Scotland
3.3.1. Leaf fall

3.3.2. Changes in Leaf Colour
4. Discussion
4.1. Leaf Behaviour Established So Far
4.2. Hypothesis Proposed
4.3. Wider Application of the Hypothesis
5. Conclusions
Acknowledgements
Author Contributions
Conflict of interest
References
- Kikuzawa, K.; Lechowicz, M.J. Ecology of Leaf Longevity; Springer: Tokyo, Japan, 2011. [Google Scholar]
- Small, E. Photosynthetic Rates in Relation to Nitrogen Cycling as an Adaptation to Nutrient Deficiency in Peat Bog Plants. Can. J. Bot. 1972, 50, 2227–2233. [Google Scholar] [CrossRef]
- Chabot, B.F.; Hicks, D.J. The Ecology of Leaf Life Spans. A. Rev. Ecol. System. 1982, 13, 229–259. [Google Scholar] [CrossRef]
- Escudero, A.; Del Arco, J.M.; Garrido, M.V. The Efficiency of Nitrogen Retranslocation from Leaf Biomass in Quercus ilex Ecosystems. Vegetatio 1992, 99/100, 225–237. [Google Scholar]
- Mediavilla, S.; Escudero, A. Leaf Life Time Differs from Retention Time of Biomass and Nutrients in the Crowns of Evergreen Species. J. Ecol. 2003, 17, 541–548. [Google Scholar]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; Flexas, J.; Garnier, E.; Groom, P.K.; Gulias, J.; Hikosaka, K.; Lamont, B.B.; Lee, T.; Lee, W.; Lusk, C.; Midgley, J.J.; Navas, N.-L.; Niinemets, Ü.; Oleksyn, J.; Osada, N.; Poorter, H.; Poot, P.; Prior, L.; Pyankov, P.I.; Roumet, C.; Thomas, S.C.; Tjoelker, M.G.; Veneklaas, E.J.; Villar, R. The Worldwide Leaf Economics Spectrum. Nature 2004, 428, 821–827. [Google Scholar]
- Marty, C.; Lamaze, T.; Pornon, A. Leaf Life-span Optimises Annual Biomass rather than Plant Photosynthetic Capacity in an Evergreen Shrub. New Phytol. 2010, 187, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Suehiro, K.; Kameyama, K. Leaf Age Composition of Evergreen Broadleaved Trees. Jap. J. Ecol. 1992, 42, 137–147, (In Japanese with English Synopsis). [Google Scholar]
- Reich, P.B.; Uhl, C.; Walters, M.B.; Prugh, L.; Ellsworth, D.S. Leaf Demography and Phenology in Amazon Rain Forest: A Census of 40,000 Leaves of 23 Species. Ecol. Monogr. 2004, 74, 3–23. [Google Scholar] [CrossRef]
- Nitta, I.; Ohsawa, M. Leaf Dynamics and Shoot Phenology of Eleven Warm-Temperate Evergreen Broad-Leaved Trees Near Their Northern Limit in Central Japan. Plant Ecol. 1997, 130, 71–88. [Google Scholar] [CrossRef]
- Rapp, M. Production de Litière et Apport au Sol d’Eléments dans Deux Écosystèmes Méditerranéens: La Forêt de Quercus ilex L. et la Garigue de Quercus coccifera L. Oecol. Plant 1969, 4, 377–410. [Google Scholar]
- Diamantoglou, S.; Mitrakos, K. Leaf longevity in Mediterranean evergreen sclerophylls. In Components of Productivity of Mediterranean-Climate Regions—Basic and Applied Aspects; Margaris, N.S., Mooney, H.A., Eds.; Junk: The Hague, Netherlands, 1981; pp. 17–19. [Google Scholar]
- Floret, C.; Galan, M.J.; Floc’h, E.L.; Leprince, F.; Romane, F. France. In Plant Pheno-Morphological Studies in Mediterranean Type Ecosystems; Orshan, G., Ed.; Kluwer: Dordrecht, The Netherlands, 1989; pp. 9–97. [Google Scholar]
- Castro-Díez, P.; Montserrat-Martí, G. Phenological Patterns of Fifteen Mediterranean Phanerophytes from Quercus ilex Communities in NE-Spain. Plant Ecol. 1998, 139, 103–112. [Google Scholar] [CrossRef]
- Montserrat-Martí, G.; Pérez-Rontomé, C. Fruit Growth Dynamics and their Effects on the Phenological Pattern of Native Pistacia populations in NE Spain. Flora 2002, 197, 161–174. [Google Scholar] [CrossRef]
- Escudero, A.; Del Arco, J.M.; Sanz, I.C.; Ayala, J. Effects of Leaf Longevity and Retranslocation Efficiency on the Retention Time of Nutrients in the Leaf Biomass of Different Woody Species. Oecologia 1992, 90, 80–87. [Google Scholar] [CrossRef]
- Grubb, P.J.; Bellingham, P.J.; Kohyama, T.S.; Piper, F.I.; Valido, A. Disturbance Regimes, Gap-demanding Trees and Seed Mass Related to Tree Height in Warm Temperate Rain Forests Worldwide. Biol. Rev. 2013, 88, 701–744. [Google Scholar] [CrossRef] [PubMed]
- Walter, H.; Lieth, H. Klimadiagrammen-Weltatlas; Fischer: Jena, Germany, 1960–1967; p. 4. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.E.; Jones, P.G.; Jarvis, A. Very High Resolution Interpolated Climate Surfaces for Global Land Areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Wu, C.Y. Vegetation of Yunnan; Science Press: Beijing, China, 1987; pp. 231–274. (In Chinese) [Google Scholar]
- Song, Y. The Essential Characteristics and Main Types of the Broad-leaved Evergreen Forest in China. Phytocoenologia 1988, 16, 105–123. [Google Scholar] [CrossRef]
- Wu, Z.Y.; Raven, P.H.; Hong, D.Y. Flora of China; Science Press: Beijing, China; Missouri Botanical Garden Press: St Louis, MO, USA, 1996–2008; Volumes 4, 7, 11–15, 19. [Google Scholar]
- Stevens, P.F. Angiosperm Phylogeny Website. Available online: www.mobot.org/MOBOT/resarch/APWeb/ (accessed on 21 July 2014).
- Fang, M.; Fang, R.; He, M.; Hu, L.; Yang, H.; Chamberlain, D.F. Rhododendron. In Flora of China, Volume 14; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St Louis, MO, USA, 2005; pp. 260–455. [Google Scholar]
- Nilsen, E.T. Quantitative Phenology and Leaf Survivorship of Rhododendron maximum L. in Contrasting Irradiance Environments of Southern Appalachian Mountains. Amer. J. Bot. 1986, 72, 822–831. [Google Scholar]
- Nilsen, E.T.; Sharifi, M.R.; Rundel, P.W. Leaf Dynamics in an Evergreen and a Deciduous Species with Even-aged Leaf Cohorts from Different Environments. Amer. Midl. Nat. 1987, 118, 46–55. [Google Scholar] [CrossRef]
- Lancaster, R. Travels in China. A Plantsman’s Paradise; Antique Collector’s Club: Woodbridge, UK, 1989; pp. 198–208. [Google Scholar]
- Kusomoto, T. Physiological and Ecological Studies on the Plant Production in Plant Communities. 3. Ecological Consideration of the Temperature-Photosynthesis Curves of Evergreen Broad-leaved Trees. Jap. J. Ecol. 1957, 7, 126–130, (In Japanese with English summary). [Google Scholar]
- Kusomoto, T. Photosynthesis and respiration in leaves of main component species. In JIBP Synthesis, Vol. 18, Biological Production in a Warm-Temperate Evergreen Oak Forest of Japan; Kira, T., Ono, Y., Hosokawa, T., Eds.; University Press: Tokyo, Japan, 1978; pp. 88–98. [Google Scholar]
- Larcher, W. Jahresgang des Assimilations- und Respirationsvermögens von Olea europaea L. ssp. sativa Hoff. et Link., Quercus ilex L., und Quercus pubescens Willd. aus dem nördlichen Gardaseegebiet. Planta 1961, 56, 575–606. [Google Scholar]
- Larcher, W. Assimilationsökologie der immergrünen Olea europea und Quercus ilex und der sommergrünen Quercus pubescens im nördlichen Gardaseegebiet. Planta 1961, 56, 607–617. [Google Scholar] [CrossRef]
- Meletiou, M.S.; Rhizopolou, S.; Diamantoglu, S. Seasonal Changes in Carbohydrates, Lipids and Nitrogen Content in Sun and Shade Leaves from Four Mediterranean Evergreen Sclerophylls. Environm. Exp. Bot. 1994, 34, 129–140. [Google Scholar] [CrossRef]
- Körner, C. Carbon Limitation in Trees. J. Ecol. 2003, 91, 4–17. [Google Scholar] [CrossRef]
- Oliveira, G.; Peñuelas, J. Effects of Winter Cold Stress on Photosynthesis and Photochemical Efficiency of PSII of the Mediterranean Cistus albidus L. and Quecus ilex L. Pl. Ecol. 2004, 175, 179–191. [Google Scholar] [CrossRef]
- Varone, L.; Gratani, L. Physiological Response of Eight Mediterranean Maquis Species to Low Air Temperatures During Winter. Photosynthetica 2007, 45, 385–391. [Google Scholar] [CrossRef]
- Ayub, G.; Smith, R.A.; Tissue, D.T.; Atkin, O.K. Impacts of Drought on Leaf Respiration in Darkness and Light in Eucalyptus saligna Exposed to Industrial-age Atmospheric CO2 and Growth Temperatures. New Phytol. 2011, 190, 1003–1018. [Google Scholar] [CrossRef] [PubMed]
- Galmés, J.; Ribas-Carbó, M.; Medrano, H.; Flexas, J. Response of Leaf Respiration to Water Stress in Mediterranean Species with Different Growth Forms. J. Arid Environ. 2007, 68, 206–222. [Google Scholar] [CrossRef]
- Gratani, L.; Varone, L.; Catoni, R. Relationship between Net Photosynthesis and Leaf Respiration in Mediterranean evergreen species. Photosynthetica 2008, 46, 567–573. [Google Scholar] [CrossRef]
- Metcalfe, D.B.; Lobo-Vale, R.; Chaves, M.M.; Maroco, J.P.; Aragao, L.E.O.C.; Malhi, Y.; Da Costa, A.L.; Braga, A.P.; Gonçalves, P.L.; de Athydes, J.; Da Costa, M.; Almeida, S.S.; Campbell, C.; Hurry, V.; Williams, M.; Meir, P. Impacts of Experimentally Imposed Drought on Leaf Respiration and Morphology in an Amazon Rain Forest. Funct. Ecol. 2010, 24, 524–533. [Google Scholar] [CrossRef]
- Kimura, M.; Suzuki, J.; Tachibana, M.; Tahara, H. Role of Evergreen Foliage in the Nitrogen Economy during Shoot Growth of Ternstroemia gymnanthera, a Warm-Temperate Broadleaf Tree. Bot. Mag. Tokyo 1983, 96, 75–83. [Google Scholar] [CrossRef]
- Yamamura, Y. Matter-economical Roles of the Evergreen Foliage of Aucuba japonica, an Understory Shrub in Warm-Temperate Region of Japan. 1. Leaf Demography, Productivity and Dry Matter Economy. Bot. Mag. Tokyo 1986, 99, 323–332. [Google Scholar]
- Minoletti, M.L.; Boerner, R.E. J. Drought and Site Fertility Effects on Foliar Nitrogen and Phosphorus Dynamics and Nutrient Resorption by the Forest Undershrub Viburnum acerifolium L. Amer. Midl. Nat. 1994, 131, 109–119. [Google Scholar] [CrossRef]
- Kramer, P.J. Plant and Soil Water Relationships: A Modern Synthesis; McGraw-Hil: New York, NY, USA, 1969; pp. 198–200. [Google Scholar]
- Karlsson, P.S. Leaf Longevity in Evergreen Shrubs: Variation Within and Among European Species. Oecologia 1992, 91, 346–349. [Google Scholar] [CrossRef]
- Leopold, A. A Sand County Almanac and Sketches Here and There; Oxford University Press: New York, NY, USA, 1949; p. 87. [Google Scholar]
- Gower, S.J.; Reich, P.B.; Son, Y. Canopy Dynamics and Aboveground Production of Five Tree Species with Different Leaf Longevities. Tree Physiol. 1993, 12, 327–345. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grubb, P.J.; Thompson, C.L.; Harper, G.H. Why Do Some Evergreen Species Keep Their Leaves for a Second Winter, While Others Lose Them? Forests 2014, 5, 2594-2612. https://doi.org/10.3390/f5112594
Grubb PJ, Thompson CL, Harper GH. Why Do Some Evergreen Species Keep Their Leaves for a Second Winter, While Others Lose Them? Forests. 2014; 5(11):2594-2612. https://doi.org/10.3390/f5112594
Chicago/Turabian StyleGrubb, Peter J., Christine L. Thompson, and Geoffrey H. Harper. 2014. "Why Do Some Evergreen Species Keep Their Leaves for a Second Winter, While Others Lose Them?" Forests 5, no. 11: 2594-2612. https://doi.org/10.3390/f5112594
APA StyleGrubb, P. J., Thompson, C. L., & Harper, G. H. (2014). Why Do Some Evergreen Species Keep Their Leaves for a Second Winter, While Others Lose Them? Forests, 5(11), 2594-2612. https://doi.org/10.3390/f5112594