
Characterization of Fungal Pathogens Associated with White Pine Needle Damage (WPND) in Northeastern North America

Abstract
:1. Introduction

2. Methods
2.1. Phylogenetic Analysis

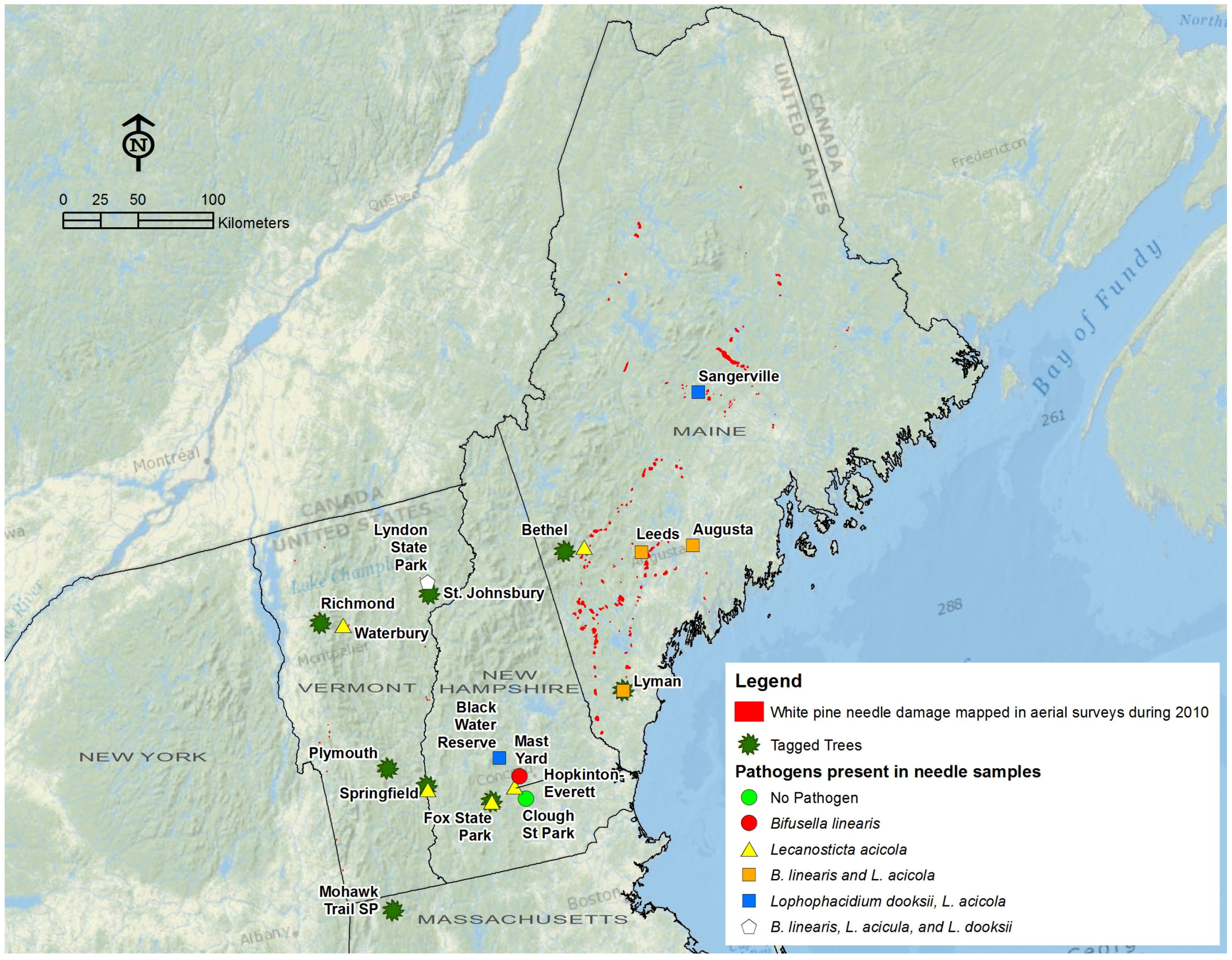{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample I.D. | Location | State | DNA a Source | Species | GenBank b Accession |
|---|---|---|---|---|---|
| EBJul30-10 | Tolland | CT | Fruit body | Hemiphacidium longisporum | KT000188 |
| EBJul30-11 | Tolland | CT | Fruit body | Hemiphacidium longisporum | KT000189 |
| WPF 111-12G | Augusta | ME | Mycelia | Hendersonia pinicola | KT000174 |
| WPF 97-12G | Augusta | ME | Mycelia | Lophodermium macci | KT000171 |
| WPF 180-12G | Augusta | ME | Mycelia | Pyrenochaeta spp. | KT000185 |
| WPF 21-12A | Bethel | ME | Mycelia | Allantophomopsiella pseudotsugae | KT000147 |
| WPF 19-12A | Bethel | ME | Mycelia | Cladosporium | KT000145 |
| WPF 157-12G | Bethel | ME | Mycelia | Hendersonia pinicola | N/A |
| WPF 2-12A | Bethel | ME | Mycelia | Lecanosticta acicola | KT000153 |
| WPF 32-12A | Bethel | ME | Mycelia | Lecanosticta acicola | N/A |
| WPF 33-12A | Bethel | ME | Mycelia | Lecanosticta acicola | N/A |
| WPF 4-12A | Bethel | ME | Mycelia | Lecanosticta acicola | KT000154 |
| WPF 5-12A | Bethel | ME | Mycelia | Lecanosticta acicola | KT000155 |
| WPF 9-12A | Bethel | ME | Mycelia | Lecanosticta acicola | KT000134 |
| WPF 28-12A | Bethel | ME | Mycelia | Phaeosphaeria pontiformis | KT000144 |
| WPF 26-12A | Bethel | ME | Mycelia | Septorioides sp. nov. | KT000142 |
| WPF 15-12A | Bethel | ME | Mycelia | Sydowia polyspora | KT000149 |
| WPF 17-12A | Bethel | ME | Mycelia | Sydowia polyspora | KT000156 |
| WPF 23-12A | Bethel | ME | Mycelia | Sydowia polyspora | KT000151 |
| WPF 69-12G | Blackwater | ME | Mycelia | Septorioides sp. nov. | N/A |
| 1528 | Leeds | ME | Mycelia | Lecanosticta acicola | KT007127 |
| 1666 | Leeds | ME | Mycelia | Lecanosticta acicola | KT007128 |
| WPF 37-12G | Leeds | ME | Mycelia | Lecanosticta acicola | N/A |
| WPF 56-12A | Leeds | ME | Mycelia | Lecanosticta acicola | KT000166 |
| WPF 44-12G | Leeds | ME | Mycelia | Lecanosticta acicola | N/A |
| WPF 51-12G | Leeds | ME | Mycelia | Lophodermium nitens | KT000163 |
| WPF 54-12G | Leeds | ME | Mycelia | Pestalotiopsis jesteri | KT000164 |
| WPF 55-12G | Leeds | ME | Mycelia | Pestalotiopsis jesteri | KT000165 |
| WPF 120-12G | Leeds | ME | Mycelia | Phaeomoniella sp. | KT000175 |
| WPF 48-12G | Leeds | ME | Mycelia | Phialocephala fluminis | KT000162 |
| WPF 42-12G | Leeds | ME | Mycelia | Septorioides sp. nov. | KT000161 |
| WPF 46-12G | Leeds | ME | Mycelia | Septorioides sp. nov. | KT000138 |
| WPF 103-12G | Lyman | ME | Mycelia | Fungal endophyte | KT000172 |
| WPF 93-12G | Lyman | ME | Mycelia | Hendersonia pinicola | N/A |
| 1671 | Lyman | ME | Mycelia | Lecanosticta acicola | KT007129 |
| WPF 102-12G | Lyman | ME | Mycelia | Septorioides sp. nov. | N/A |
| WPF 106-12G | Lyman | ME | Mycelia | Septorioides sp. nov. | KT000173 |
| WPF 158-12G | Lyman | ME | Mycelia | Septorioides sp. nov. | N/A |
| WPF 90-12G | Lyman | ME | Mycelia | Septorioides sp. nov. | KT000170 |
| WPF 151-12G | Sangerville | ME | Mycelia | Fungal endophyte | KT000179 |
| WPF 152-12G | Sangerville | ME | Mycelia | Hendersonia pinicola | N/A |
| WPF 85-12G | Sangerville | ME | Mycelia | Hendersonia pinicola | KT000169 |
| WPF 86-12G | Sangerville | ME | Mycelia | Hendersonia pinicola | N/A |
| WPF 87-12G | Sangerville | ME | Mycelia | Hendersonia pinicola | N/A |
| 1675 | Sangerville | ME | Mycelia | Lecanosticta acicola | KT007130 |
| WPF 14-12A | Sangerville | ME | Mycelia | Paraconiothyrium spp. | KT000176 |
| WPF 150-12G | Sangerville | ME | Mycelia | Rhizosphaera kalkhoffii | KT000178 |
| WPF 121-12G | Sangerville | ME | Mycelia | Septorioides sp. nov. | KT000140 |
| WPF 147-12G | Sangerville | ME | Mycelia | Septorioides sp. nov. | KT000139 |
| WPF 149-12G | Sangerville | ME | Mycelia | Septorioides sp. nov. | KT000135 |
| WPF 18-12A | Sangerville | ME | Mycelia | Septorioides sp. nov. | KT000157 |
| WPF 22-12A | Sangerville | ME | Mycelia | Sydowia polyspora | KT000148 |
| WPF 73-12G | Blackwater | NH | Mycelia | Lecanosticta acicola | KT000168 |
| EBJul30-4 | Gilmanton | NH | Fruit body | Hendersonia pinicola | KT000192 |
| 1678 | Hillsboro | NH | Mycelia | Lecanosticta acicola | KT007131 |
| WPF 75-12G | Hopkinton | NH | Mycelia | Hendersonia pinicola | N/A |
| WPF 21-12A | Hopkinton | NH | Mycelia | Septorioides sp. nov. | N/A |
| WPF 25-12A | Hopkinton | NH | Mycelia | Septorioides sp. nov. | KT000160 |
| WPF 27-12A | Hopkinton | NH | Mycelia | Septorioides sp. nov. | KT000143 |
| WPF 71-12G | Mast Yard | NH | Mycelia | Dothiodiomycete | KT000167 |
| 1679 | Merrimack | NH | Mycelia | Lecanosticta acicola | KT007132 |
| EBJul30-16 | Monadnock | NH | Fruit body | Hendersonia pinicola | KT000191 |
| EBJul30-2 | New Hampton | NH | Fruit body | Hemiphacidium longisporum | KT000190 |
| EBJul30-9 | New Hampton | NH | Fruit body | Lophophacidium dooksii | KT000187 |
| EBJul30-15 | Sanbornton | NH | Fruit body | Bifusella linearis | KT000195 |
| EBJul30-5 | Scituate | RI | Fruit body | Bifusella linearis | KT000193 |
| WPF 16-12A | Bethel | VT | Mycelia | Sydowia polyspora | KT000150 |
| 1681 | Brookfield | VT | Mycelia | Lecanosticta acicola | KT007133 |
| EBJul30-6 | Plymouth | VT | Mycelia | Bifusella linearis | KT000194 |
| EBJul30-1 | Plymouth | VT | Mycelia | Lophophacidium dooksii | KT000186 |
| V-3-4 | Richmond | VT | Mycelia | Septorioides sp. nov. | KT000137 |
| WPF 172-12G | Springfield | VT | Mycelia | Fungal endophyte | KT000183 |
| WPF 165-12G | Springfield | VT | Mycelia | Septorioides sp. nov. | KT000180 |
| WPF 167-12G | Springfield | VT | Mycelia | Septorioides sp. nov. | KT000181 |
| WPF 175-12G | Springfield | VT | Mycelia | Septorioides sp. nov. | KT000184 |
| WPF 170-12G | Springfield | VT | Mycelia | Sydowia polyspora | KT000182 |
| WPF 176-12G | Springfield | VT | Mycelia | Sydowia polyspora | N/A |
| WPF 174-12G | Springfield | VT | Mycelia | Hendersonia pinicola | N/A |
| V-5-2 | Underhill | VT | Mycelia | Septorioides sp. nov. | KT000136 |
| V-1-4 | Waterbury | VT | Mycelia | Hendersonia pinicola | KT000152 |
| 1506 | Waterbury | VT | Mycelia | Lecanosticta acicola | KT007125 |
| 1518 | Waterbury | VT | Mycelia | Lecanosticta acicola | KT007126 |
| WPF 144-12G | Waterbury | VT | Mycelia | Pestalotiopsis uvicola | KT000177 |
| WPF 20-12A | Waterbury | VT | Mycelia | Septorioides sp. nov. | KT000146 |
| WPF 24-12A | Waterbury | VT | Mycelia | Septorioides sp. nov. | KT000141 |
| V-1-9 | Waterbury | VT | Mycelia | Discostroma fuscellum | KT000158 |
2.2. Long-Term White Pine Health Monitoring
3. Results
3.1. Morphologic Diversity of Fungi Associated with WPND


3.2. Genetic Diversity of Fungi Associated with WPND

3.3. Long-Term White Pine Health Monitoring

4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- McDowell, N.G.; Beerling, D.J.; Breshears, R.A.; Breshears, D.D.; Fisher, R.A.; Raffa, K.F.; Stitt, M. The interdependence of mechanisms underlying climate-driven vegetation mortality. Trends Ecol. Evolut. 2011, 26, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Weed, A.S.; Ayres, M.P.; Hicke, J.A. Consequences of climate change for biotic disturbances in North American forests. Ecol. Monogr. 2013, 83, 441–470. [Google Scholar] [CrossRef]
- Woods, A. Is the health of British Columbia’s forests being influenced by climate change? If so, was this predictable? Can. J. Plant Pahol. 2011, 33, 117–126. [Google Scholar] [CrossRef]
- Hicke, J.A.; Allen, C.D.; Desai, A.R.; Dietze, M.C.; Hall, R.J.; Hogg, E.H. (Ted); Kashian, D.M.; Moore, D.; Raffa, K.F.; Sturrock, R.N.; et al. Effects of biotic disturbances on forest carbon cycling in the United States and Canada. Glob. Chang. Biol. 2012, 18, 7–34. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Groenewald, J.Z.; de Jesús Yáñez-Morales, M.; Crous, P.W. DNA barcoding of Mycosphaerella species of quarantine importance to Europe. Persoonia 2012, 29, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Laflamme, G.; Broders, K.; Cote, C.; Munck, I.; Iriarte, G.; Innes, L. Priority of Lophophacidium over Canavirgella: Taxonomic status of Lophophacidium dooksii and Canavirgella banfieldii, causal agents of a white pine needle disease. Mycologia 2015, 107, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Merrill, W.; Wenner, N.G.; Dreisbach, T.A. Canavirgella banfieldii gen. and sp. nov.: A needlecast fungus on pine. Can. J. Bot.-Rev. Can. Bot. 1996, 74, 1476–1481. [Google Scholar] [CrossRef]
- Skilling, D.D.; Nicholls, T.H. Brown Spot Needle Disease: Biology and Control in Scotch Pine Plantatinos; USDA Forest Service Research Paper NC-109; U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station: St. Paul, MN, USA, 1974; p. 23. [Google Scholar]
- Huang, Z.Y.; Smalley, E.B.; Guries, R.P. Differentiation of Mycosphaerella dearnessii by cultural charactes and RAPD analysis. Phytopathology 1995, 85, 522–527. [Google Scholar] [CrossRef]
- Munck, I.A.; Ostrofsky, W.D.; Burns, B. Pest Alert: Eastern White Pine Needle Damage; NA-PR-01-11; USDA Forest Service, Northeastern Area State and Private Forestry: Newtown, PA, USA, 2011. [Google Scholar]
- U.S. Forest Service. Forest Health Highlights. Available online: http://fhm.fs.fed.us/fhh/fhmusamap.shtml (accessed on 16 June 2015).
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Kumar, S.; Tamura, K.; Nei, M. MEGA3: Integrated software for molecular evolutionary analysis and sequence alignment. Brief. Bioinform. 2004, 5, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 6, 227–242. [Google Scholar] [CrossRef]
- Rozas, J.; Sanchez-DelBarrio, J.C.; Messenguer, X.; Rozas, R. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 2003, 19, 2496–2497. [Google Scholar] [CrossRef] [PubMed]
- Corlett, M.; Shoemaker, R.A. Lophophacidium dooksii n. sp., a phacidiaceous fungus on needles of white pine. Can. J. Bot. 1984, 62, 1836–1840. [Google Scholar] [CrossRef]
- Horst, R.K.; Westcott, C. Westcott’s Plant Disease Handbook, 7th ed.; Springer: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Jankovsky, L.; Palovcikova, D.; Tomsovsky, M. Brown spot needle blight associated with Mycosphaerella dearnessii occurs on Pinus rotundata in the Czech Republic. Plant Pathol. 2009, 58. [Google Scholar] [CrossRef]
- Jurc, D.; Jurc, M. Mycosphaerella dearnessii occurs in Slovenia. Plant Pathol. 2010, 59, 808. [Google Scholar] [CrossRef]
- Darker, G.D. A revision of the genera of the Hypodermataceae. Can. J. Bot. 1967, 45, 1399–1444. [Google Scholar] [CrossRef]
- Hyde, K.D.; McKenzie, E.H.C.; KoKo, T.W. Towards incorporating anamorphic fungi in a natural classification—Checklist and notes for 2010. Mycosphere 2011, 2, 1–88. [Google Scholar]
- Wijayawardene, N.N.; McKenzie, E.H.C.; Hyde, K.D. Towards incorporating anamorphic fungi in a natural classification checklist and notes from 2011. Mycosphere 2012, 3, 157–228. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Verkley, G.J.M.; Shin, H.D.; Barreto, R.W.; Alfenas, A.C.; Swart, W.J.; Groenewald, J.Z.; Crous, P.W. Sizing up Septoria. Stud. Mycol. 2013, 75, 307–390. [Google Scholar] [CrossRef] [PubMed]
- Morin, L.; Shivas, R.G.; Piper, M.C.; Tan, Y.P. Austropleospora osteospermi gen. et sp. nov. and its host specificity and distribution on Chrysanthemoides monilifera ssp. rotundata in Australia. Fungal Divers. 2010, 40, 65–74. [Google Scholar] [CrossRef]
- Kaneko, S.; Fujioka, H.; Zinno, Y. A new species of Septoria on Japanese black pine. Trans. Mycol. Soc. Jpn. 1989, 30, 463–466. [Google Scholar]
- Wyka, S.; Broders, K.D. Septorioides strobus sp. nov., a new species, within the new family Septorioideaceae, associated with needle defoliation of eastern white pine (Pinus strobus L.). Fungal. Biol. 2015; submitted for publication. [Google Scholar]
- NOAA National Centers for Environmental Information. State of the Climate: National overview for June 2015. Available online: http://www.ncdc.noaa.gov/sotc/national/201507 (accessed on 10 August 2015).
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Chang. 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Hintsteiner, M.; Cech, T.L.; Halmschlager, E.; Stauffer, C.; Kirisits, T. First report of Mycosphaerella dearnessii on Pinus nigra var. nigra in Austria. For. Pathol. 2012, 42, 437–440. [Google Scholar] [CrossRef]
- Frumhoff, P.C.; McCarthy, J.J.; Melillo, J.M.; Moser, S.C.; Wuebbles, J.D. Confronting Climate Change in the U.S. Northeast; Synthesis Report of the Northeast Climate Impact Assessment (NECIA); Union of Concerned Scientist (UCS): Cambridge, MA, US, 2007. [Google Scholar]
- Broders, K.D.; Wyka, S.; Munck, I. Characterization and distribution of fungi associated with needle defoliation of eastern white pine (Pinus strobus). Int. For. Rev. 2014, 16, 322. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Broders, K.; Munck, I.; Wyka, S.; Iriarte, G.; Beaudoin, E. Characterization of Fungal Pathogens Associated with White Pine Needle Damage (WPND) in Northeastern North America. Forests 2015, 6, 4088-4104. https://doi.org/10.3390/f6114088
Broders K, Munck I, Wyka S, Iriarte G, Beaudoin E. Characterization of Fungal Pathogens Associated with White Pine Needle Damage (WPND) in Northeastern North America. Forests. 2015; 6(11):4088-4104. https://doi.org/10.3390/f6114088
Chicago/Turabian StyleBroders, Kirk, Isabel Munck, Stephen Wyka, Gloria Iriarte, and Eric Beaudoin. 2015. "Characterization of Fungal Pathogens Associated with White Pine Needle Damage (WPND) in Northeastern North America" Forests 6, no. 11: 4088-4104. https://doi.org/10.3390/f6114088
APA StyleBroders, K., Munck, I., Wyka, S., Iriarte, G., & Beaudoin, E. (2015). Characterization of Fungal Pathogens Associated with White Pine Needle Damage (WPND) in Northeastern North America. Forests, 6(11), 4088-4104. https://doi.org/10.3390/f6114088