
A Process-Based Approach to Estimate Chinese Fir (Cunninghamia lanceolata) Distribution and Productivity in Southern China under Climate Change




Abstract
:1. Introduction
2. Experimental Section
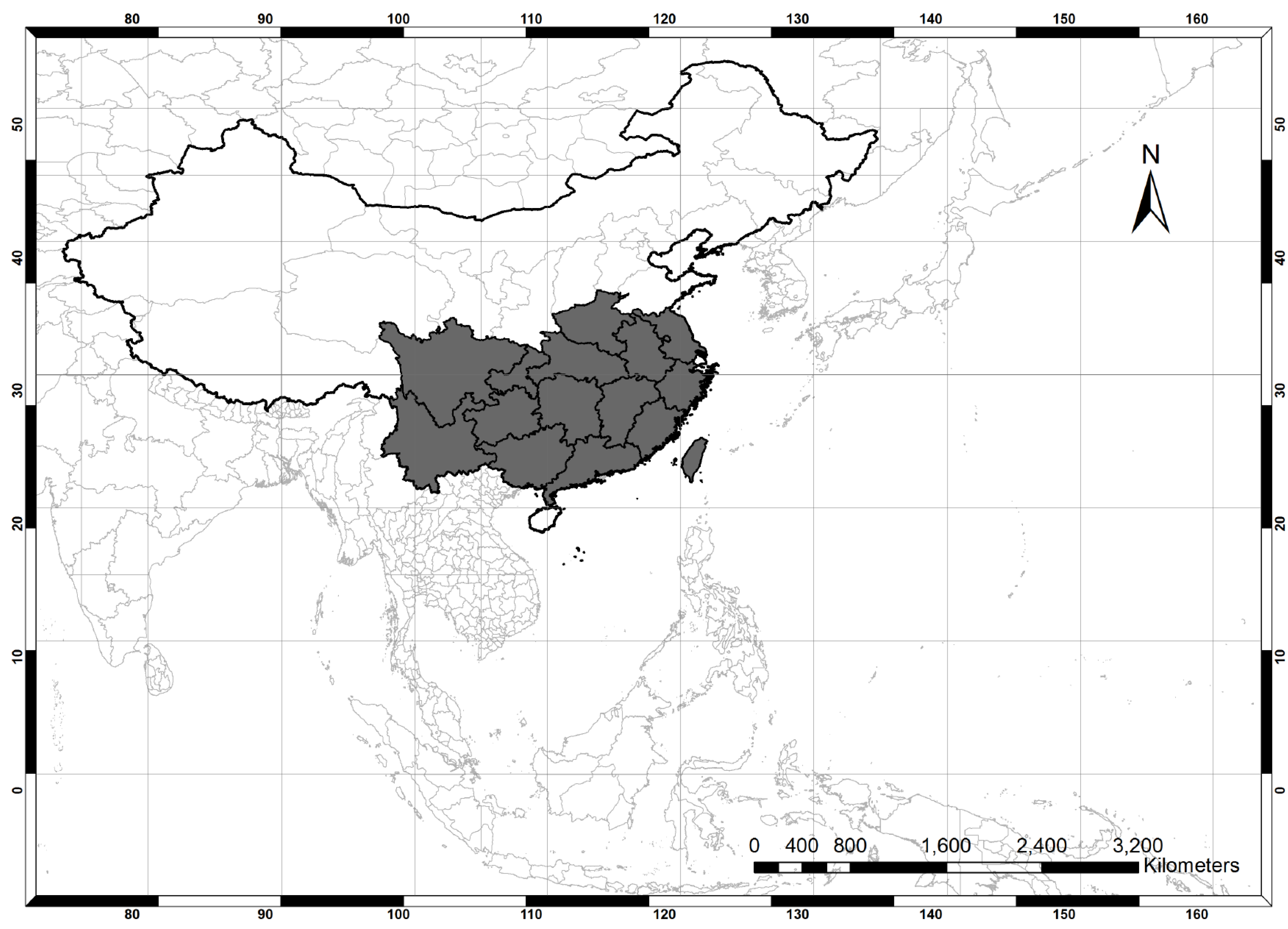
2.1. Study Area

2.2. Climate Data
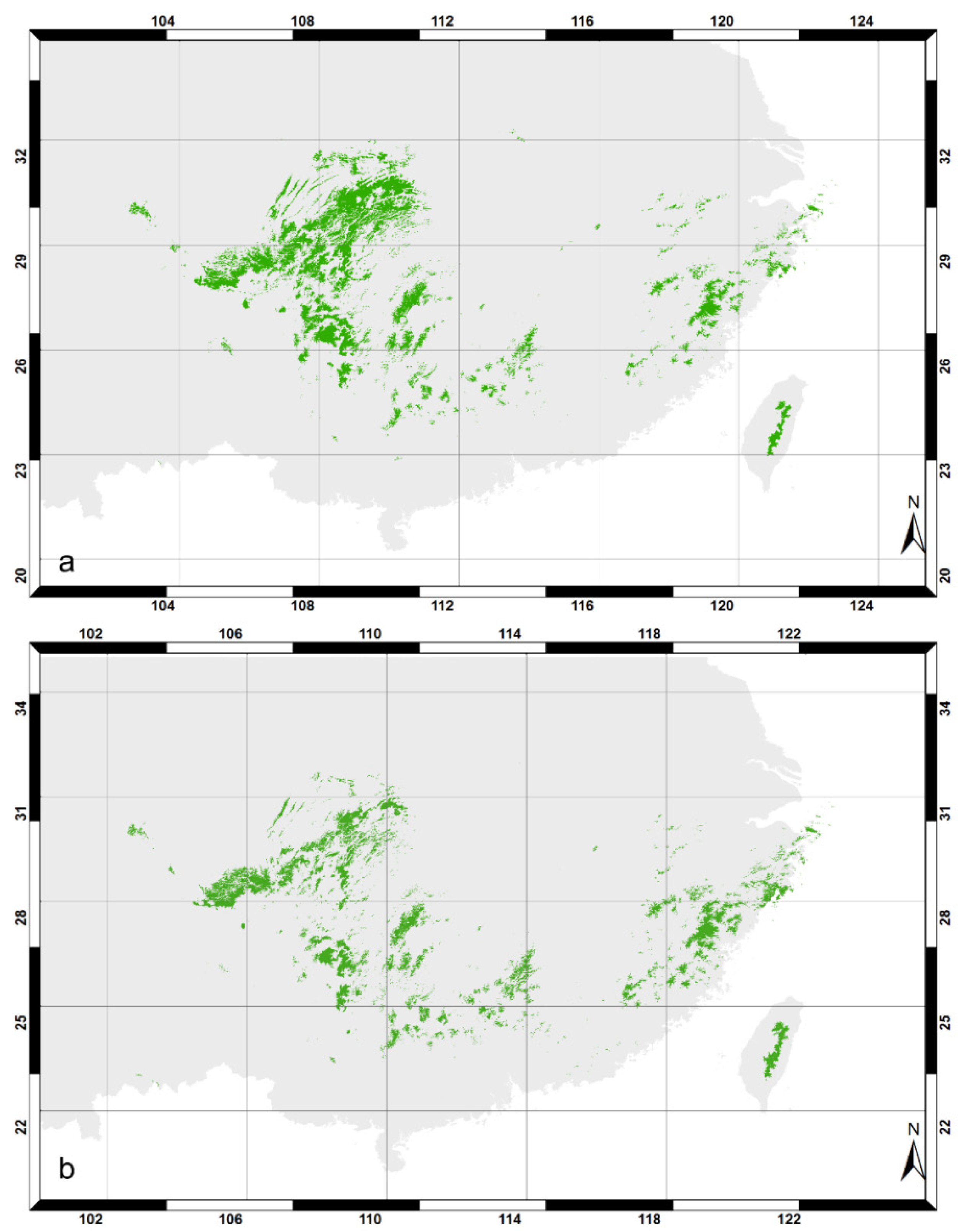
2.3. Chinese Fir Presence Data
2.4. 3-PG Model Description

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Meaning/Comments | Name | Unit | Value | Source |
|---|---|---|---|---|
| Allometric relationships & partitioning | ||||
| Foliage: Stem partitioning ratio @ D = 2 cm | pFS2 | 0.72 | This study | |
| Foliage: Stem partitioning ratio @ D = 20 cm | pFS20 | 0.38 | This study | |
| Constant in the stem mass v. diameter. relationship | aS | 0.0118 | [32] | |
| Power in the stem mass v. diameter. relationship | nS | 3.223 | [32] | |
| Maximum fraction of net primary production (NPP) to roots | pRx | 0.6 | [32] | |
| Minimum fraction of NPP to roots | pRn | 0.2 | [32] | |
| Litterfall & root turnover | ||||
| Maximum litterfall rate | gammaFx | 1/month | 0.0485 | [32] |
| Litterfall rate at t = 0 | gammaF0 | 1/month | 0.001 | [32] |
| Age at which litterfall rate has median value | tgammaF | months | 23 | This study |
| Average monthly root turnover rate | gammaR | 1/month | 0.02 | [32] |
| Temperature modifier (fT) | ||||
| Minimum temperature for growth | Tmin | degree °C | 0 | [32] |
| Optimum temperature for growth | Topt | degree °C | 17.5 | [32] |
| Maximum temperature for growth | Tmax | degree °C | 40 | [32] |
| Frost modifier (fFRost) | ||||
| Days production lost per frost day | kF | days | 1.0 | [32] |
| Soil water modifier (fSW) | ||||
| Moisture ratio deficit for fq = 0.5 | SWconst | 0.6 | This study | |
| Power of moisture ratio deficit | SWpower | 7 | This study | |
| Fertitlity effects | ||||
| Value of “m” when FR = 0 | m0 | 0 | Default | |
| Value of “fNutr” when FR = 0 | fN0 | 0.4 | This study | |
| Power of (1-FR) in “fNutr” | fNn | 1 | This study | |
| Age modifier (fAge) | ||||
| Maximum stand age used in age modifier | MaxAge | years | 150 | This study |
| Power of relative age in function for fAge | nAge | 4 | Default | |
| Relative age to give fAge = 0.5 | rAge | 0.95 | Default | |
| Stem mortality & self-thinning | ||||
| Max. stem mass per tree @ 1000 trees/hectare | wSx1000 | kg/tree | 175 | This study |
| Power in self-thinning rule | thinPower | 1.3 | This study | |
| Specific leaf area | ||||
| Specific leaf area at age 0 | SLA0 | m2/kg | 8 | This study |
| Specific leaf area for mature leaves | SLA1 | m2/kg | 4 | This study |
| Age at which specific leaf area = (SLA0+SLA1)/2 | tSLA | years | 3 | [32] |
| Light interception | ||||
| Extinction coefficient for absorption of PAR by canopy | K | 0.5 | Default | |
| Age at canopy cover | fullCanAge | years | 3 | [32] |
| Maximum proportion of rainfall evaporated from canopy | MaxIntcptn | 0.033 | This study | |
| LAI for maximum rainfall interception | LAImaxIntcptn | 5 | This study | |
| Production and respiration | ||||
| Canopy quantum efficiency | alpha | molC/molPAR | 0.033 | [32] |
| Ratio NPP/GPP | Y | 0.5 | [32] | |
| Conductance | ||||
| Maximum canopy conductance | MaxCond | m/s | 0.02 | [32] |
| LAI for maximum canopy conductance | LAIgcx | 3 | This study | |
| Defines stomatal response to VPD | CoeffCond | 1/mBar | 0.05 | Default |
| Canopy boundary layer conductance | BLcond | m/s | 0.2 | This study |
| Basic Density | ||||
| Minimum basic density—For young trees | rhoMin | t/m3 | 0.3 | This study |
| Maximum basic density—For older trees | rhoMax | t/m3 | 0.37 | This study |
2.5. Model Runs
2.5.1. Distribution Modeling
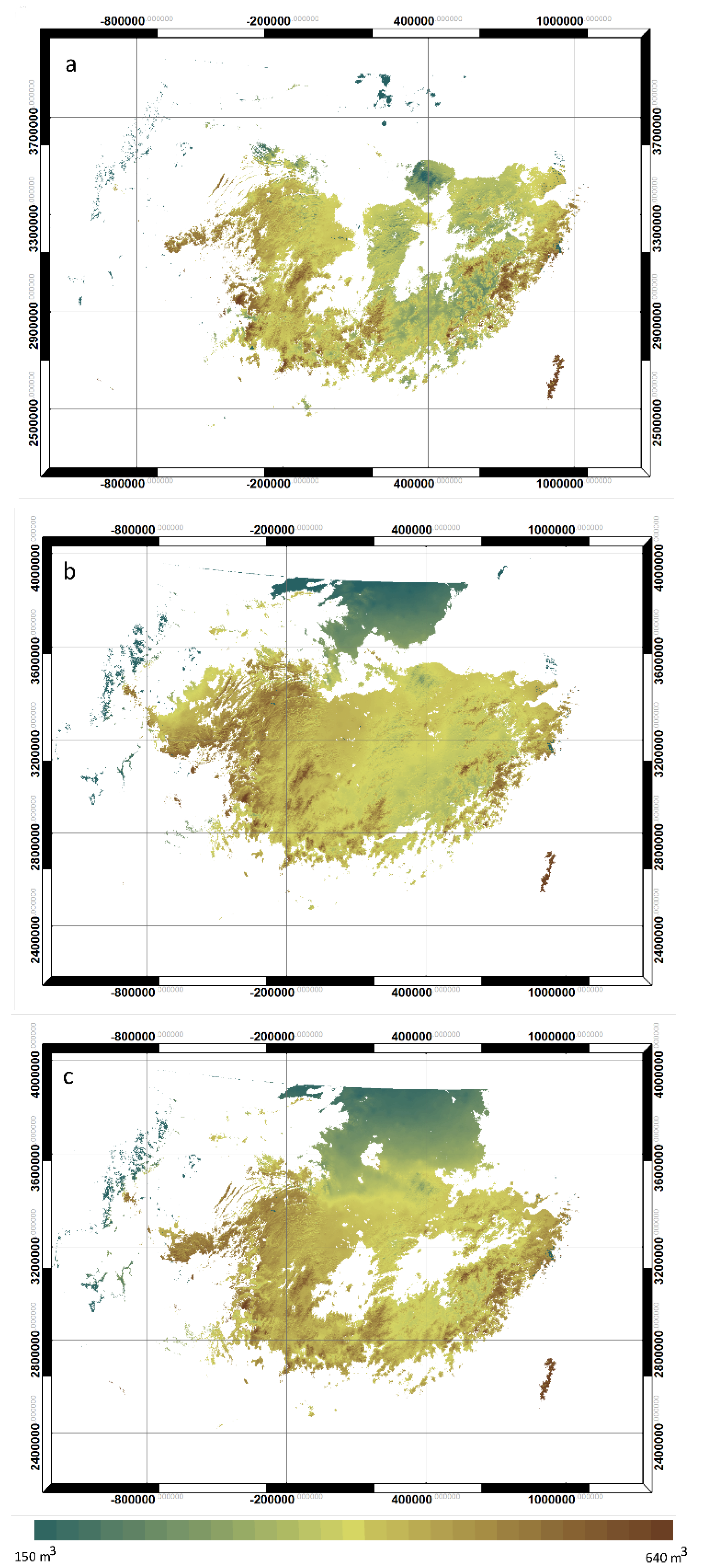
2.5.2. Productivity Modeling
3. Results and Discussion
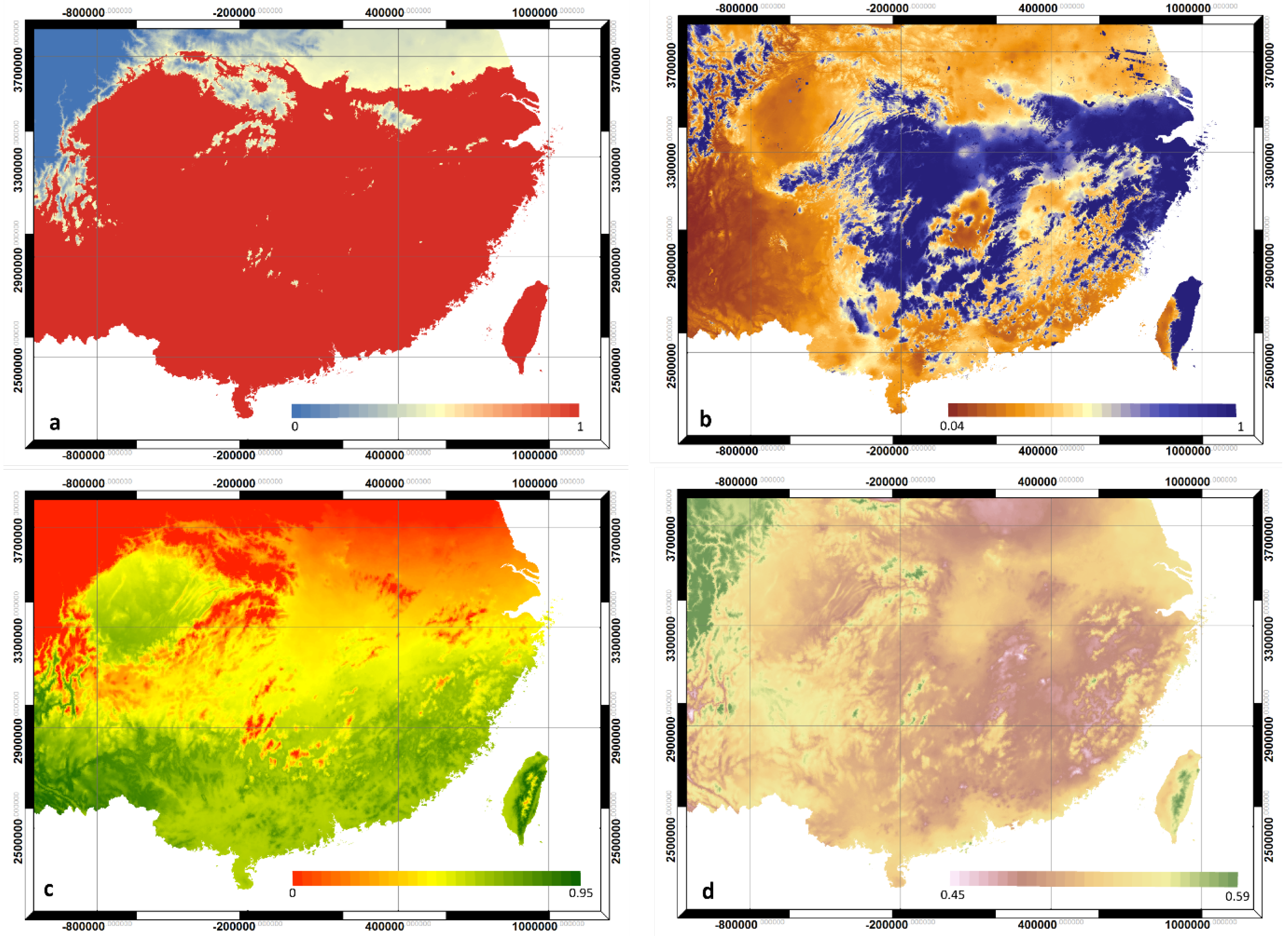
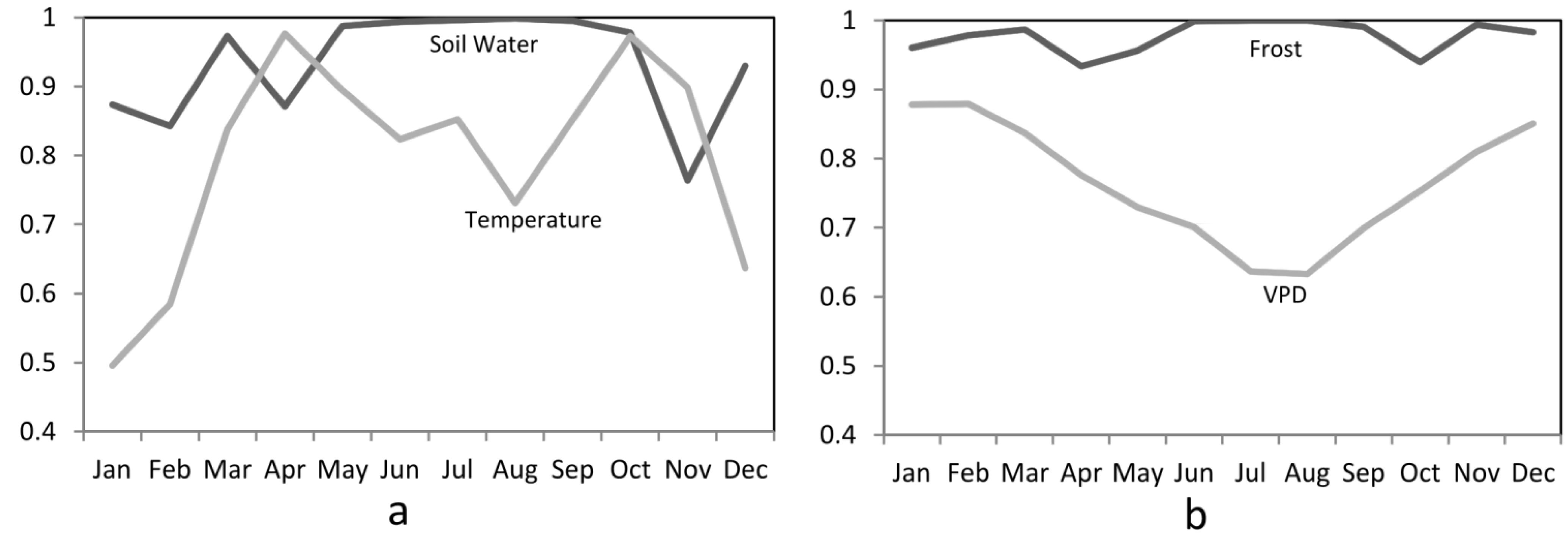
3.1. Climate Modifiers



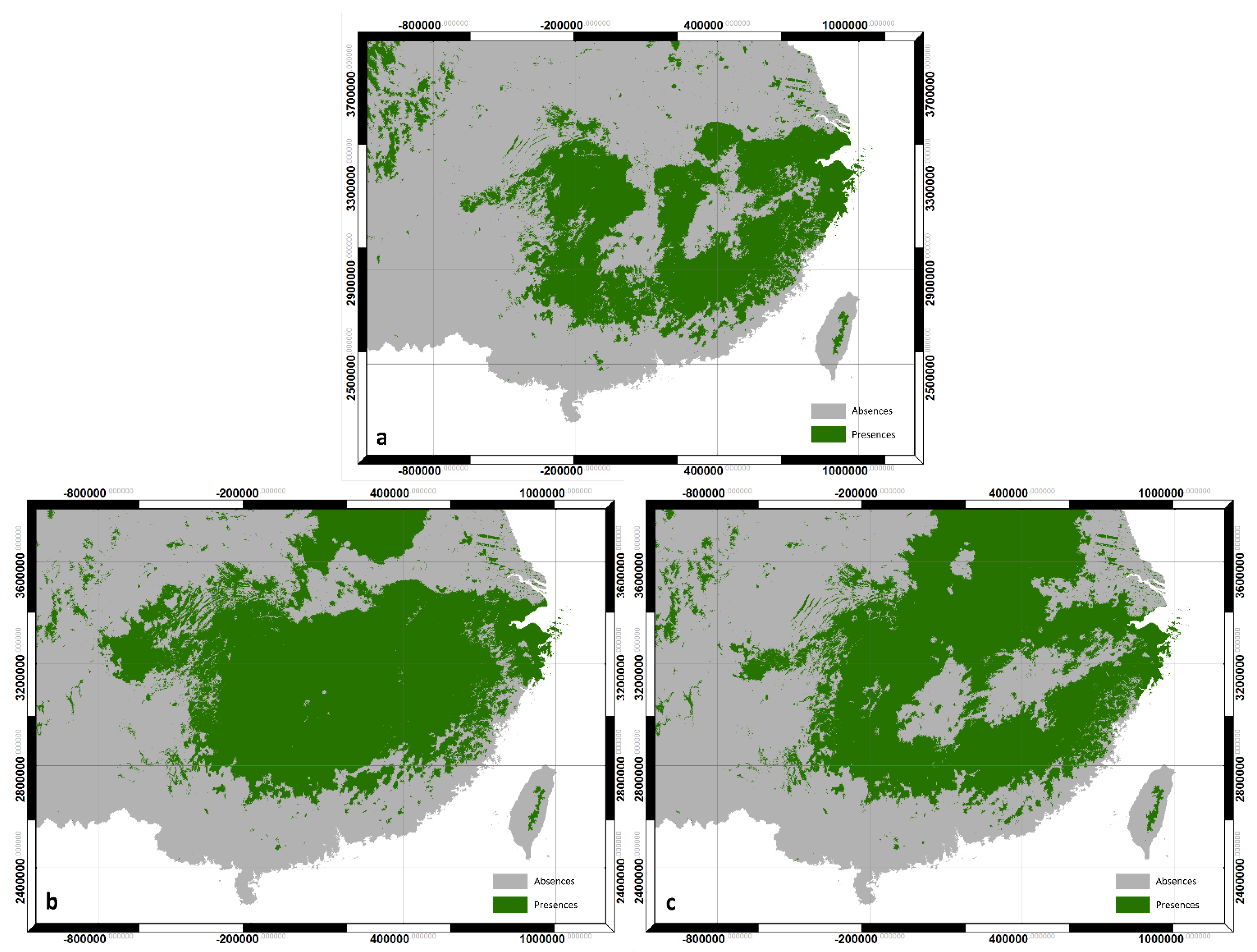
3.2. Decision Tree
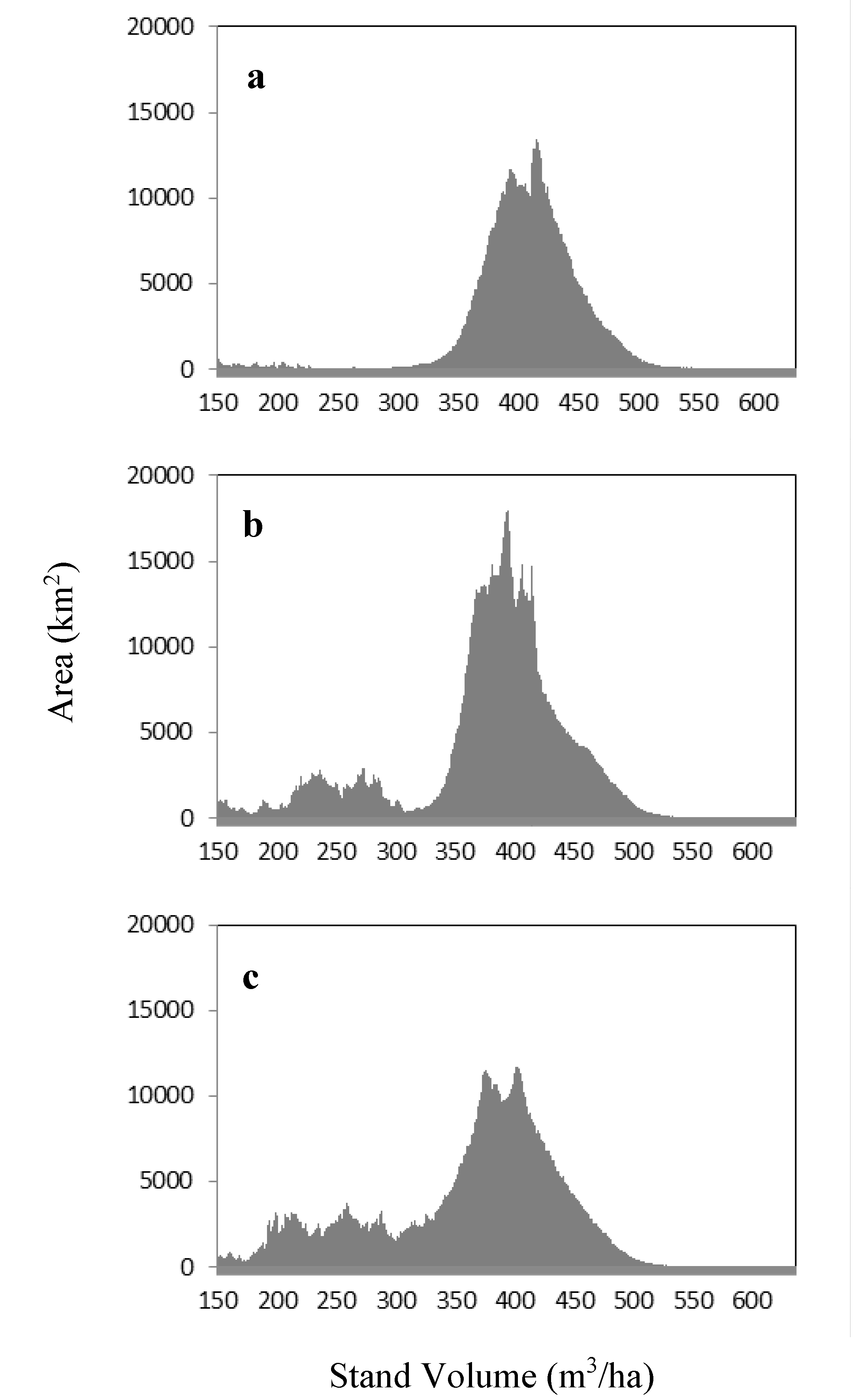
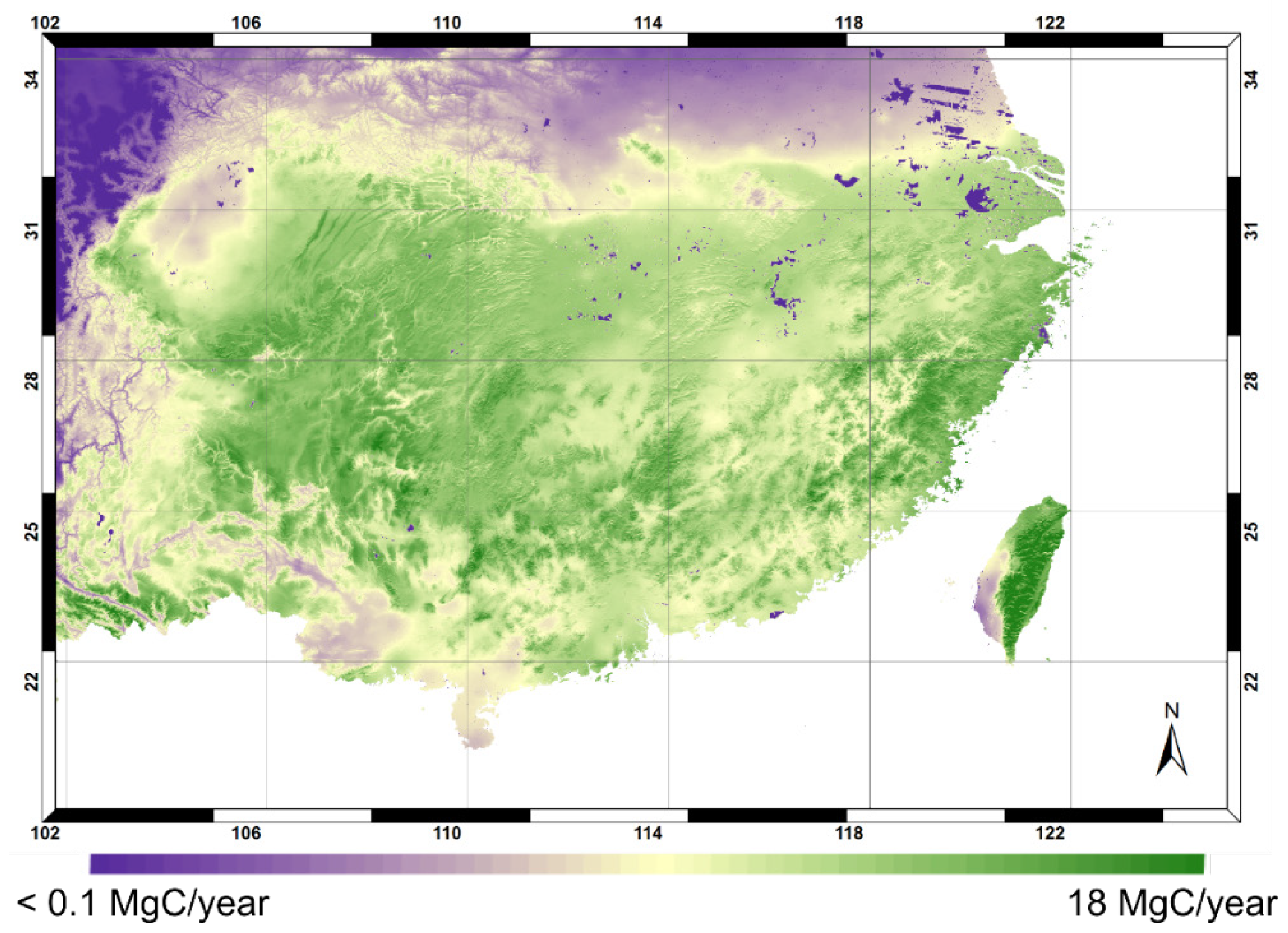
3.3. Productivity and NPP Estimates






3.4. Model Application and Limitation
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wu, Z.L. Chinese-fir; China Forestry Publishing House: Beijing, China, 1984. (In Chinese) [Google Scholar]
- Yu, X. Silviculture of Chinese fir; Fujian Sciences & Technology Press: Fuzhou, China, 1997. (In Chinese) [Google Scholar]
- Yang, Y.S. Studies on the Sustainable Management of Chinese Fir Plantations; Chinese Forestry Press: Beijing, China, 1998. [Google Scholar]
- Lei, J.F. Forest Resources in China; China Forestry Publish House: Beijing, China, 2005; p. 172. [Google Scholar]
- He, Z.Y. Study on problems of soil degradation in continuous cropping Chinese fir plantation. In Research on Site Degradation of Timber Plantation; Sheng, W.T., Ed.; Chinese Science & Technology Press: Beijing, China, 1992; pp. 243–250. (In Chinese) [Google Scholar]
- CMOF. Statistics of Forest Resource in China (1977–1981); Department of Forest Resource and Management, CMOF: Beijing, China, 1983. (In Chinese) [Google Scholar]
- CMOF. Statistics of Forest Resource in China (1984–1988); Department of Forest Resource and Management, CMOF: Beijing, China, 1989. (In Chinese) [Google Scholar]
- CMOF. Statistics of Forest Resource in China (1989–1993); Department of Forest Resource and Management, CMOF: Beijing, China, 1994. (In Chinese) [Google Scholar]
- CMOF. Statistics of Forest Resource in China (1994); Department of Forest Resource and Management, CMOF: Beijing, China, 1995. (In Chinese) [Google Scholar]
- SFA. State Forestry Administration (SFA) Statistics of Forest Resource in China (1994–1998); Department of Forest Resource and Management, SFA: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Wang, B.; Li, S.N.; Guo, H. The assessment of forest ecosystem services evaluation in Jiangxi Province. Jiangxi Sci. 2007, 5, 554–587. [Google Scholar]
- Huang, M.; Ji, J.; Li, K.; Liu, Y.; Yang, F.; Tao, B. The ecosystem carbon accumulation after conversion of grasslands to pine plantations in subtropical red soil of South China. Tellus 2007, 59, 439–448. [Google Scholar] [CrossRef]
- Fang, Q. Influences of continuous cropping on soil fertility and stand growth in Chinese fir plantation. Sci. Silv. Sin. 1987, 23, 389–397. (In Chinese) [Google Scholar]
- Yang, Y.; Zhang, R.; He, Z. Change on the stand productivity of 29-year-old Chinese fir plantation in different rotations. J. Fujian Col. For. 1997, 18, 202–206. (In Chinese) [Google Scholar]
- Ma, X.; Fan, S.; Chen, S.; Lin, S. Study on biomass productivity of Chinese fir plantations after successive planting. Sci. Silv. Sin. 2003, 39, 78–83. [Google Scholar]
- Andrewartha, H.G.; Birch, L.C. The Distribution and Abundance of Animals; University of Chicago Press: Chicago, IL, USA, 1954. [Google Scholar]
- McCarty, J.P. Ecological consequences of recent climate change. Conserv. Biol. 2001, 15, 320–331. [Google Scholar] [CrossRef]
- Qian, W.; Zhu, Y. Climate change in China from 1880 to 1998 and its impact on the environmental condition. Clim. Chang. 2001, 50, 419–444. [Google Scholar] [CrossRef]
- Wang, Z.; Ding, Y.; He, J.; Jun, Y. An updated analysis of the climate change in China in recent 50 years. Acta Meteorol. Sin. 2004, 62, 228–236. [Google Scholar]
- Ding, Y.; Ren, G.; Zhao, Z.; Xu, Y.; Luo, Y.; Li, Q.; Zhang, J. Detection, causes and projection of climate change over China: An overview of recent progress. Adv. Atmos. Sci. 2007, 24, 954–971. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yu, D.; Xun, B.; Sun, Y.; Hao, R. The potential effects of climate change on the distribution and productivity of Cunninghamia lanceolata in China. Environ. Monit. Assess. 2014, 186, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Hamann, A.; Spittlehouse, D.L.; Murdock, T. ClimateWNA—High-resolution spatial climate data for Western North America. J. Appl. Meteorol. 2012, 51, 16–29. [Google Scholar] [CrossRef]
- Daly, C. Variable Influence of Terrain on Precipitation Patterns: Delineation and Use of Effective Terrain Height in PRISM; Oregon State University: Corvallis, OR, USA, 2002. Available online: http://oldprism.nacse.org/pub/prism/docs/effectiveterrain-daly.pdf (accessed on 1 July 2014).
- Hijmans, R.J.; Cameron, S.; Parra, J.; Jones, P.G.; Jarvis, A. WorldClim, Version 1.3; University of California, Berkeley: Berkeley, CA, USA, 2005. [Google Scholar]
- IPCC Data Distribution Center. Available online: http://www.ipcc-data.org/sim/gcm_global/index.html (accessed on 1 March 2014).
- Landsberg, J.J.; Waring, R.H. A generalised model of forest productivity using simplified concepts of radiation-use efficiency, carbon balance and partitioning. For. Ecol. Manag. 1997, 95, 209–228. [Google Scholar] [CrossRef]
- Coops, N.C.; Hember, R.A.; Waring, R.H. Assessing the impact of current and projected climates on Douglas-Fir productivity in British Columbia, Canada, using a process-based model (3-PG). Can. J. For. Res. 2010, 40, 511–524. [Google Scholar] [CrossRef]
- Landsberg, J.; Coops, N.C. Modeling forest productivity across large areas and long periods. Nat. Res. Model. 1999, 12, 383–411. [Google Scholar] [CrossRef]
- Coops, N.C.; Waring, R.H.; Law, B.E. Assessing the past and future distribution and productivity of ponderosa pine in the Pacific Northwest using a process model, 3-PG. Ecol. Model. 2005, 183, 107–124. [Google Scholar] [CrossRef]
- Liu, X.Z.; Xiang, W.H.; Kang, W.X. Biomass dynamic pattern of Chinese fir plantation ecosystem. In Long-term Located Research on Forest Ecosystem; Liu, X.Z., Ed.; China Forestry Publish House: Beijing, China, 1993; pp. 18–22. [Google Scholar]
- Zhao, M.; Xiang, W.; Peng, C.; Tian, D. Simulating age-related changes in carbon storage and allocation in a Chinese fir plantation growing in southern China using the 3-PG model. For. Ecol. Manag. 2009, 257, 1520–1531. [Google Scholar] [CrossRef]
- Pearce, J.L.; Boyce, M.S. Modelling distribution and abundance with presence-only data. J. Appl. Ecol. 2006, 43, 405–412. [Google Scholar] [CrossRef]
- Fang, J.; Liu, G.; Xu, S. Biomass and net production of forest vegetation in China. Acta Ecol. Sin. 1996, 16, 497–508. [Google Scholar]
- De’Ath, G. Multivariate regression trees: A new technique for modeling species-environment relationships. Ecology 2002, 83, 1105–1117. [Google Scholar]
- Schwalm, C.R.; Black, T.A.; Amiro, B.D.; Arain, M.A.; Barr, A.G.; Bourque, C.P.A.; Dunn, A.L.; Flanagan, L.B.; Giasson, M.A.; Lafleur, P.M.; et al. Photosynthetic light use efficiency of three biomes across an east-west continental-scale transect in Canada. Agric. Meteorol. 2006, 140, 269–286. [Google Scholar] [CrossRef]
- Breiman, L.; Friedman, J.; Stone, C.J.; Olshen, R.A. Classification and Regression Trees; CRC Press: Bota Raton, FL, USA, 1984. [Google Scholar]
- Melendez, K.V.; Jones, D.L.; Feng, A.S. Classification of communication signals of the little brown bat. J. Acoust. Soc. Am. 2006, 120, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Zaniewski, A.E.; Lehmann, A.; Overton, J.M. Predicting species spatial distributions using presence-only data: A case study of native New Zealand ferns. Ecol. Model. 2002, 157, 261–280. [Google Scholar] [CrossRef]
- Congalton, R.G. A review of assessing the accuracy of classifications of remotely sensed data. Remote Sens. Environ. 1991, 37, 35–46. [Google Scholar] [CrossRef]
- Yang, J.; Lin, T.; Kung, F. Geographic variation of China fir (Cunninghamia lanceolata (Lamb.) Hook) in Taiwan: Analysis of provenance division by ecotype and by province. Taiwan J. For. Sci. 2000, 15, 303–315. [Google Scholar]
- Chung, J.D.; Lin, T.P.; Tan, Y.C.; Lin, M.Y.; Hwang, S.Y. Genetic diversity and biogeography of Cunninghamia konishii (Cupressaceae), an island species in Taiwan: A comparison with Cunninghamia lanceolata, a mainland species in China. Mol. Phylogenet. Evol. 2004, 33, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Yu, X. Chinese fir; Fujian Sciences & Technology Press: Fuzhou, China, 1982. (In Chinese) [Google Scholar]
- Coops, N.C.; Hember, R.A. Physiologically derived predictions of Douglas-fir site index in British Columbia. For. Chron. 2009, 85, 733–744. [Google Scholar] [CrossRef]
- Coops, N.C.; Waring, R.H. A process-based approach to estimate lodgepole pine (Pinus contorta Dougl.) distribution in the Pacific Northwest under climate change. Clim. Ch. 2011, 105, 313–328. [Google Scholar] [CrossRef]
- Woodward, F.I. The impact of low temperatures in control- ling the geographical distribution of plants. Philos. Trans. B 1990, 326, 585–593. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef]
- Woodward, F.I.; Beerling, D.J. The dynamics of vegetation change: Health warnings for equilibrium “dodo” models. Glob. Ecol. Biogeogr. Lett. 1997, 6, 413–418. [Google Scholar] [CrossRef]
- Cramer, W.; Bondeau, A.; Woodward, F.I.; Prentice, I.C.; Betts, R.A.; Brovkin, V.; Cox, P.M.; Fisher, V.; Foley, J.A.; Friend, A.D.; et al. Global response of terres-trial ecosystem structure and function to CO2 and climate change: Results from six dynamic global vegetation models. Glob. Chang. Biol. 2001, 7, 357–373. [Google Scholar] [CrossRef]
- Sykes, M.T.; Prentice, I.C.; Smith, B.; Cramer, W.; Venevsky, S. An introduction to the European terrestrial ecosystem modelling activity. Glob. Ecol. Biogeogr. 2001, 10, 581–593. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Coops, N.C.; Wang, T.; Wang, G. A Process-Based Approach to Estimate Chinese Fir (Cunninghamia lanceolata) Distribution and Productivity in Southern China under Climate Change. Forests 2015, 6, 360-379. https://doi.org/10.3390/f6020360
Lu Y, Coops NC, Wang T, Wang G. A Process-Based Approach to Estimate Chinese Fir (Cunninghamia lanceolata) Distribution and Productivity in Southern China under Climate Change. Forests. 2015; 6(2):360-379. https://doi.org/10.3390/f6020360
Chicago/Turabian StyleLu, Yuhao, Nicholas C. Coops, Tongli Wang, and Guangyu Wang. 2015. "A Process-Based Approach to Estimate Chinese Fir (Cunninghamia lanceolata) Distribution and Productivity in Southern China under Climate Change" Forests 6, no. 2: 360-379. https://doi.org/10.3390/f6020360
APA StyleLu, Y., Coops, N. C., Wang, T., & Wang, G. (2015). A Process-Based Approach to Estimate Chinese Fir (Cunninghamia lanceolata) Distribution and Productivity in Southern China under Climate Change. Forests, 6(2), 360-379. https://doi.org/10.3390/f6020360