
If Long-Term Resistance to a Spruce Beetle Epidemic is Futile, Can Silvicultural Treatments Increase Resilience in Spruce-Fir Forests in the Central Rocky Mountains?

Abstract
:1. Introduction
2. Methods
2.1. Study Species
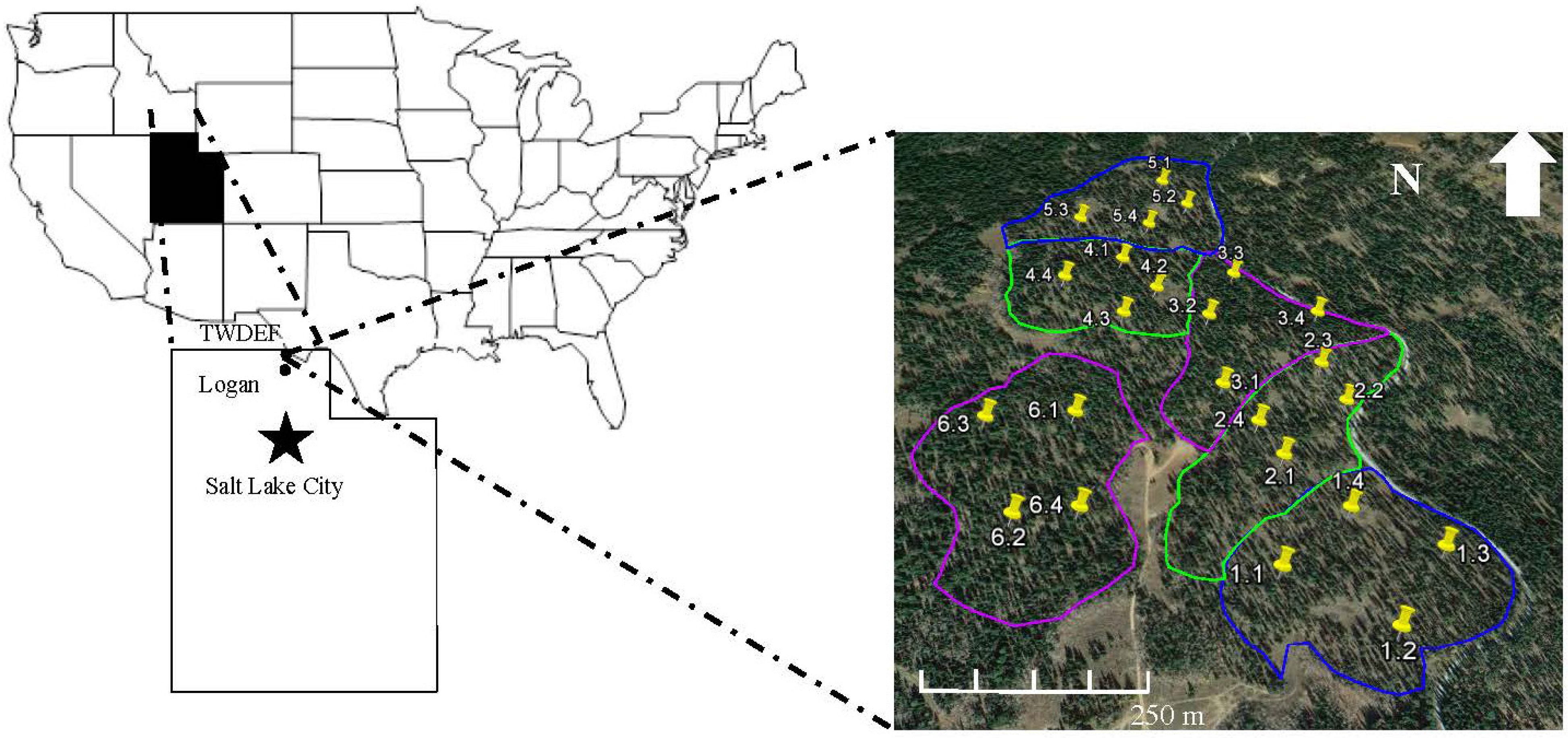
2.2. Site Description

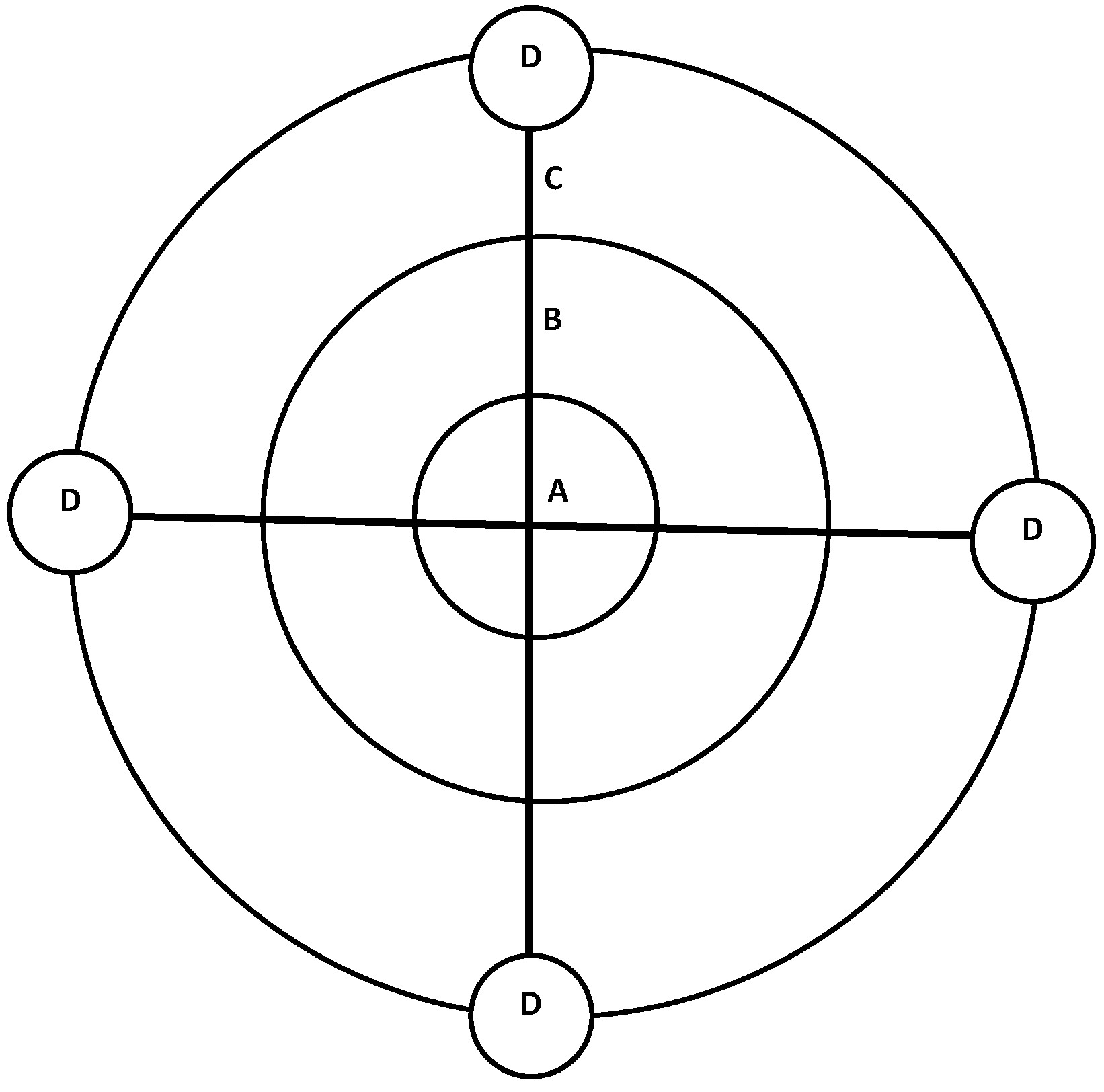
2.3. Silvicultural Treatments

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T.W. Daniel Experimental Forest Spruce Beetle Management Timeline | |
|---|---|
| Year | Event |
| 1996 |
|
| 1997 |
|
| 1998 |
|
| 1999 |
|
| 2000–2005 |
|
| 2006 |
|
| 2008 |
|
| 2013 | Remeasurement of permanent plots |

2.4. Defining Resistance and Resilience Metrics
2.4.1. Resistance
| Physiographic Location/Site Index | QMD of Spruce >25.4 cm dbh | Stand Basal Area (m2·ha−1) | Proportion of Stand That Is Spruce (%) | |
|---|---|---|---|---|
| High (3) | Spruce on well-drained sites in creek bottoms | >40.6 cm | >34. 44 | >65 |
| Medium (2) | Spruce on sites with site index of 24.4 to 36.6 m | 30.5–40.6 cm | 22.96–34.44 | 50–65 |
| Low (1) | Spruce on sites with site index of 12.2 to 24.4 m | <30.5 cm | <22.96 | <50 |
2.4.2. Resilience
2.5. Analysis
3. Results
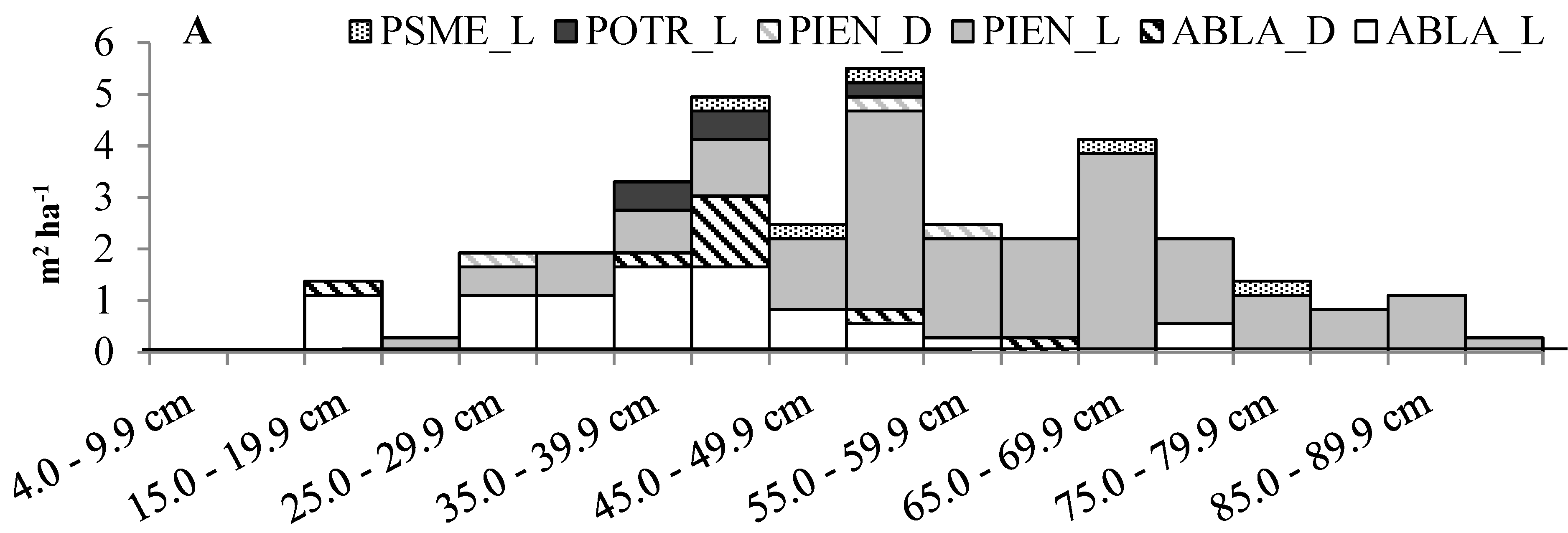
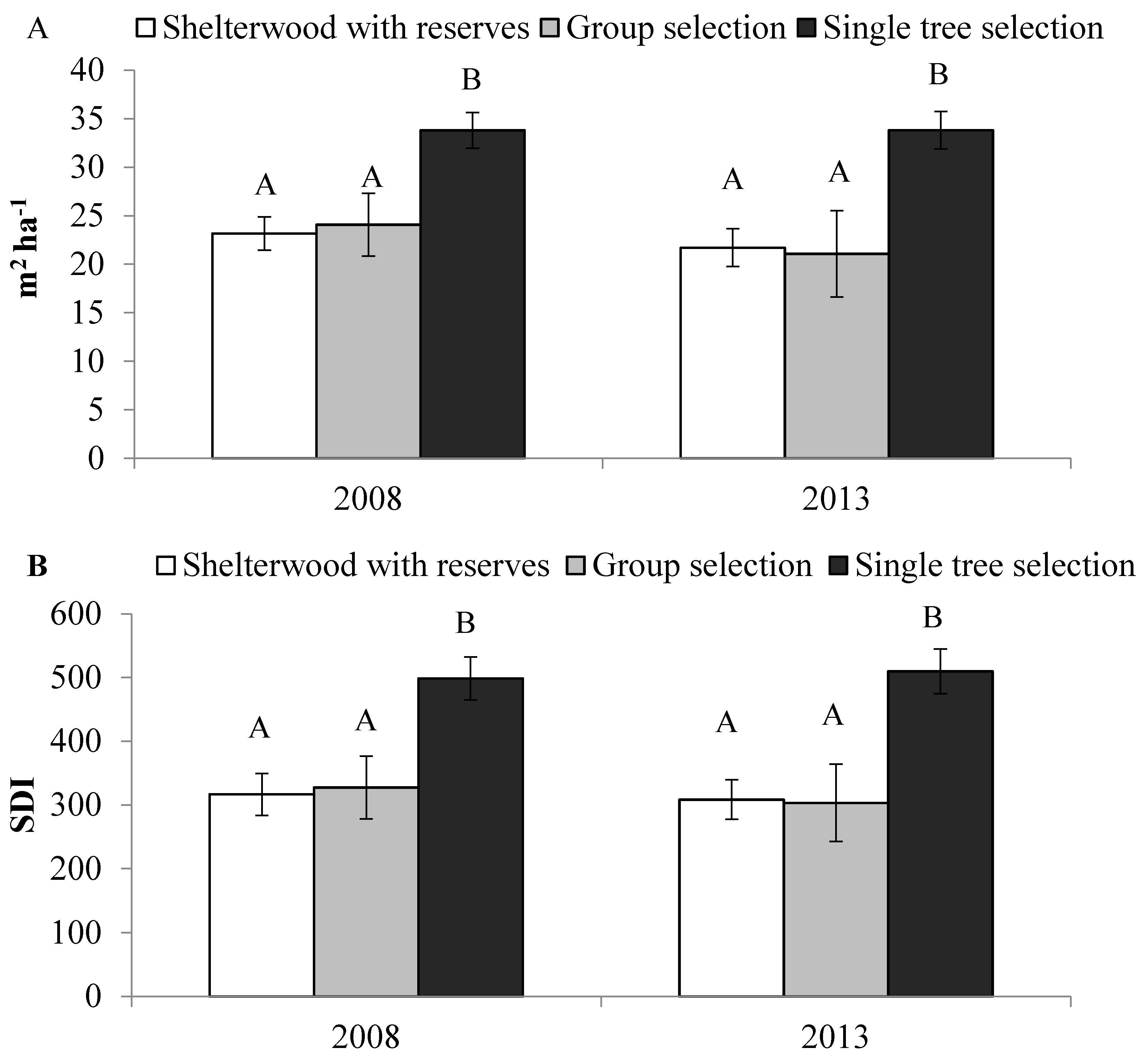
3.1. Overstory Composition, Structure, and Density



3.2. Spruce Beetle Risk Rating System
| Physiographic Location/Site Index | QMD of Spruce >25.4 cm dbh | Stand Basal Area (m2·ha−1) | Proportion of Stand That is Spruce (%) | Total Risk Rating | |||||
|---|---|---|---|---|---|---|---|---|---|
| Pre | <24.4 m | (1) | 55.1 | (3) | 33.0 | (2) | 65 | (2) | 8 |
| 2008 | |||||||||
| Shelterwood with reserves | <24.4 m | (1) | 67.0 | (3) | 23.2 | (2) | 64 | (2) | 8 |
| Group selection | <24.4 m | (1) | 64.4 | (3) | 24.1 | (2) | 76 | (3) | 9 |
| Single tree selection | <24.4 m | (1) | 62.1 | (3) | 33.8 | (2) | 84 | (3) | 9 |
| 2013 | |||||||||
| Shelterwood with reserves | <24.4 m | (1) | 69.6 | (3) | 21.7 | (1) | 65 | (2) | 7 |
| Group selection | <24.4 m | (1) | 66.3 | (3) | 21.1 | (1) | 81 | (3) | 8 |
| Single tree selection | <24.4 m | (1) | 62.9 | (3) | 33.8 | (2) | 84 | (3) | 9 |
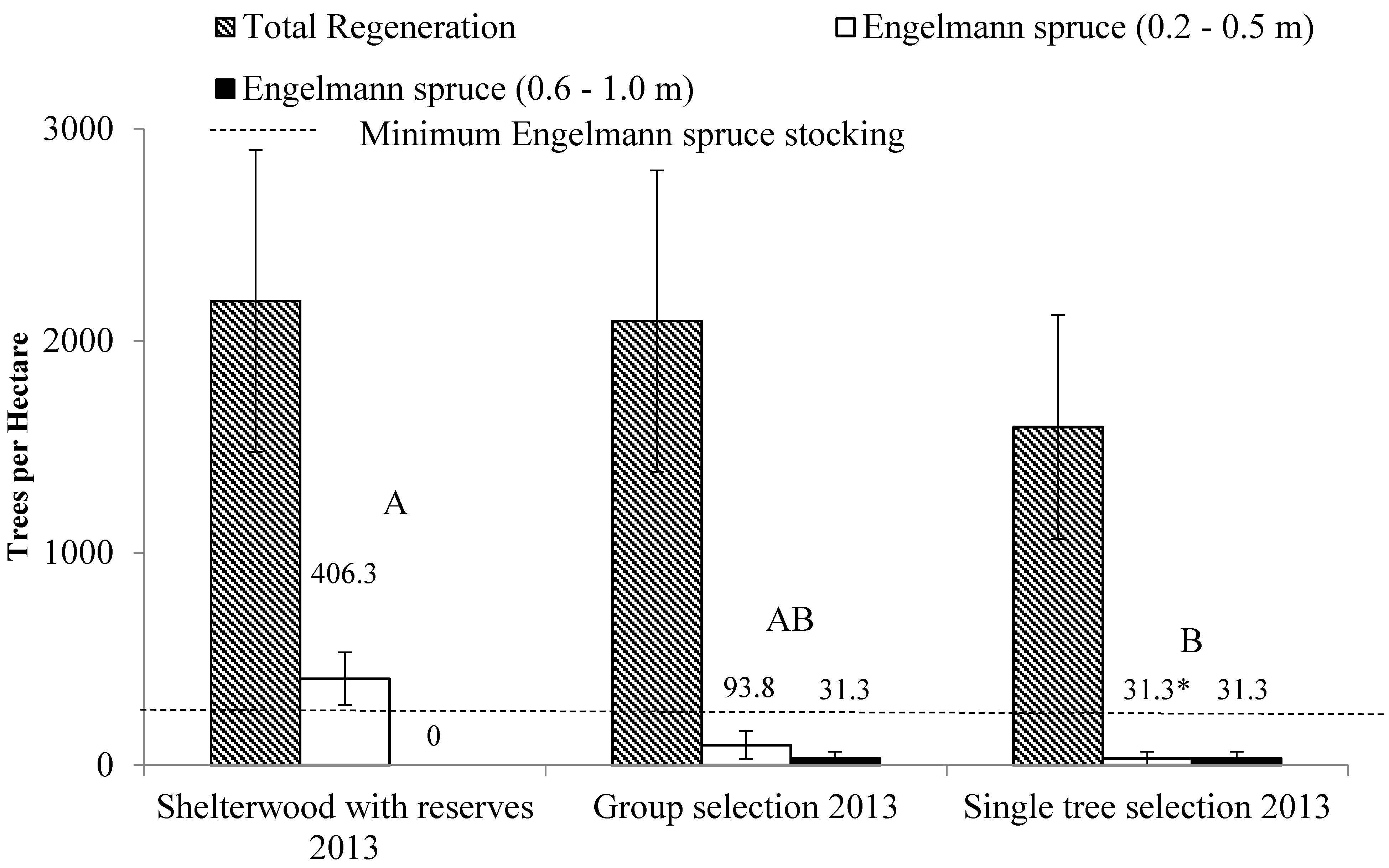
3.3. Regeneration
| Treatment | Subalpine Fir | Lodgepole Pine | Engelmann Spruce | Aspen | Grand Total | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2008 | ||||||||||
| Shelterwood with reserves | 812.5 | (187.5) | 0.0 | (0) | 0.0 | (0) | 750.0 | (566.9) | 1562.5 | (640.4) |
| Group selection | 437.5 | (147.5) | 0.0 | (0) | 0.0 | (0) | 250.0 | (182.9) | 687.5 | (181.5) |
| Single tree selection | 812.5 | (244.4) | 0.0 | (0) | 125.0 | (47.5) | 281.3 | (185.6) | 1218.8 | (264.9) |
| 2013 | ||||||||||
| Shelterwood with reserves | 812.5 | (220.3) | 218.8 | (218.8) | 406.3 | (124.4) | 750.0 | (592.0) | 2187.5 | (711.4) |
| Group selection | 1031.3 | (524.9) | 0.0 | (0) | 125.0 | (66.8) | 937.5 | (633.4) | 2093.8 | (710.2) |
| Single tree selection | 781.3 | (269.1) | 0.0 | (0) | 62.5 | (62.5) | 750.0 | (491.0) | 1593.8 | (528.1) |

4. Discussion
| Shelterwood with Reserves | Group Selection | Single Tree Selection | |
|---|---|---|---|
| Stand Level | |||
| Reduced Basal Area | X | X | |
| Retention of Groups & Gaps | X | X | |
| Diversity of Overstory Species | X | X | |
| Minimum Levels of Spruce Regeneration | X | ||
| Landscape Level | |||
| Ability to Treat Large Areas | X |
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Spittlehouse, D.; Stewart, R. Adaptation to climate change in forest management. J. Ecosyst. Manag. 2003, 4, 1–11. [Google Scholar]
- Millar, C.I.; Stephenson, N.L.; Stephens, S.L. Climate change and forests of the future: Managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; Mcdowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: IPCC Fifth Assessment Synthesis Report; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 1–115. [Google Scholar]
- Grossnickle, S.C. Ecophysiology of Northern Spruce Species: The Performance of Planted Seedlings; National Research Council of Canada: Ottawa, Canada, 2000; pp. 1–10. [Google Scholar]
- Faccoli, M.; Bernardinelli, I. Composition and elevation of spruce forests affect susceptibility of bark beetle attacks: Implications for forest management. Forests 2014, 5, 88–102. [Google Scholar] [CrossRef]
- Holsten, E.H.; Munson, R.W.; Munson, A.S.; Gibson, K.E. The Spruce Beetle, USDA Forest Service, Forest Insect and Disease Leaflet 127; USDA Forest Service: Washington, DC, USA, 1999; pp. 1–11. [Google Scholar]
- Berg, E.E.; Henry, J.D.; Fastie, C.L.; DeVolder, A.D.; Matsuoka, S.M. Spruce beetle outbreaks on the Kenai Peninusula, Alaska and Kluane National Park and Reserve, Yukon Territory: Relationship to summer temperature and regional differences in disturbance regimes. For. Ecol. Manag. 2006, 227, 219–232. [Google Scholar] [CrossRef]
- Fettig, C.J.; Klepzig, K.D.; Billings, R.F.; Munson, A.S.; Nebeker, T.E.; Negron, J.F.; Nowak, J.T. The effectiveness of vegetation management practices for prevention and control of bark beetle outbreaks in coniferous forests of the western and southern United States. For. Ecol. Manag. 2007, 238, 24–53. [Google Scholar] [CrossRef]
- Bentz, B.J.; Régnière, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negrón, J.F.; Seybold, S.J. Climate change and bark beetles of the western United States and Canada: Direct and indirect effects. BioScience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Juday, G.P. Spruce beetles, budworms, and climate warming. Available online: http://www.cgc.uaf.edu/Newsletter/gg6_1/beetles.html (accessed on 3 February 2015).
- Alexander, R.R. Ecology, Silviculture and Management of the Engelmann Spruce—Subalpine fir Type in the Central and Southern Rocky Mountains; Research Paper RM-121; USDA Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1987; p. 121. [Google Scholar]
- Kärvemo, S.; Rogell, B.; Schroeder, M. Dynamics of spruce bark beetle infestation spots: Importance of local population size and landscape characteristics after a storm disturbance. For. Ecol. Manag. 2014, 334, 232–240. [Google Scholar] [CrossRef]
- Alexander, R.R. Silviculture of Subalpine Forests in the Central and Southern Rocky Mountains: The Status of our Knowledge; Research Paper RM-121; USDA Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1973; p. 88. [Google Scholar]
- Schmid, J.M.; Hinds, T.E. Development of Spruce-Fir Stand Following Spruce Beetle Outbreaks; Research Note RM-131; USDA Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1974; p. 16. [Google Scholar]
- Mielke, J.L. Rate of deterioration of beetle-killed Engelmann spruce. J. For. 1950, 48, 882–888. [Google Scholar]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. BioScience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- DeRose, R.J.; Long, J.N. Factors influencing the spatial and temporal dynamics of Engelmann spruce mortality during a spruce beetle outbreak on the Markagunt Plateau, Utah. For. Sci. 2012, 58, 1–14. [Google Scholar]
- Forest Inventory and Analysis National Program. Available online: http://apps.fs.fed.us/fia/fido/index.html (accessed on 17 November 2014).
- Hart, S.J.; Veblen, T.T.; Eisenhart, K.S.; Jarvis, D.; Kulakowski, D. Drought induces spruce beetle (Dendroctonus rufipennis) outbreaks across northwestern Colorado. Ecology 2014, 95, 930–939. [Google Scholar] [CrossRef] [PubMed]
- McCambridge, W.F.; Knight, F.B. Factors affecting spruce beetles during a small outbreak. Ecology 1972, 53, 830–839. [Google Scholar] [CrossRef]
- Dymerski, A.D.; Anhold, J.A.; Munson, A.S. Spruce beetle (Dendroctonus rufipennis) outbreaks in Engelmann spruce (Picea engelmannii) in central Utah, 1986–1998. West. N. Am. Nat. 2001, 61, 19–24. [Google Scholar]
- Hansen, E.M.; Bents, B.J.; Turner, D.L. Temperature-based model for predicting univoltine brood proportions in spruce beetle (Coleoptera: Scolytidae). Can. Entomol. 2001, 133, 827–841. [Google Scholar] [CrossRef]
- Eaton, C.B. Influence of the mountain pine beetle on the composition of mixed pole stands of ponderosa pine and white fir. J. For. 1941, 39, 710–713. [Google Scholar]
- Schmid, J.M.; Frye, R.H. Stand Ratings for Spruce Beetles; Research Note RM-309; USDA Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1976; p. 4. [Google Scholar]
- Temperli, C.; Hart, S.J.; Veblen, T.T.; Kulakowski, D.; Hicks, J.J.; Andrus, R. Are density reduction treatments effective at managing for resistance or resilience to spruce beetle disturbance in the southern Rocky Mountains? For. Ecol. Manag. 2014, 334, 53–64. [Google Scholar] [CrossRef]
- Seidl, R. The shape of ecosystem management to come: Anticipating risks and fostering resilience. Bioscience 2014, 16, 1159–1169. [Google Scholar] [CrossRef]
- DeRose, R.J.; Long, J.N. Resistance and resilience: A conceptual framework for silviculture. For. Sci. 2014, 60, 1205–1212. [Google Scholar]
- Bentz, B.J.; Munson, A.S. Spruce beetle population suppression in northern Utah. West. J. Appl. For. 2000, 15, 122–128. [Google Scholar]
- Munson, S. Management Guide for Spruce Beetle: Dendroctonus Rufipennis Kirby. In Forest Health Protection and State Forestry Organizations; USDA Forest Service: Washington, DC, USA, 2010; p. 16. [Google Scholar]
- Hansen, E.M.; Negron, J.F.; Munson, A.S.; Anhold, J.A. A retrospective assessment of partial cutting to reduce spruce beetle-caused mortality in southern Rocky Mountains. West. J. Appl. For. 2010, 25, 81–87. [Google Scholar]
- DeRose, R.J.; Long, J.N. Disturbance, structure, and composition: Spruce beetle and Engelmann spruce forests on the Markagunt Plateau. For. Ecol. Manag. 2007, 244, 16–23. [Google Scholar] [CrossRef]
- Alexander, R.R. Silvicultural Systems and Cutting Methods for Old-Growth Spruce-Fir Forests in the Central and Southern Rocky Mountains; General Technical Report RM-126; USDA Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1986. [Google Scholar]
- Fiedler, C.E.; McCaughey, W.W.; Schmidt, W.C. Natural Regeneration in Intermountain Spruce-Fir Forests—A Gradual Process; USDA Forest Service Research Paper INT-343; USDA Forest Service, Intermountain Forest and Range Experiment Station: Odgen, UT, USA, 1985; p. 16. [Google Scholar]
- Peet, R.K. Forests and meadows of the Rocky Mountains. In North American Terrestrial Vegetation, 2nd ed.; Barbour, M.G., Billings, W.D., Eds.; Cambridge University Press: New York, NY, USA, 2000; pp. 75–121. [Google Scholar]
- Schmid, J.M.; Frye, R.H. Spruce Beetle in the Rockies; General Technical Report GTR RM-49; USDA Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1977; p. 38. [Google Scholar]
- Aplet, G.H.; Laven, R.D.; Smith, F.W. Patterns of community dynamics in Colorado Engelmann spruce-subalpine fir forests. Ecology 1988, 63, 312–319. [Google Scholar] [CrossRef]
- Veblen, T.T.; Hadley, K.S.; Reid, M.S.; Rebertus, A.J. Blowdown and stand development in a Colorado subalpine forest. Can. J. of For. Res. 1989, 19, 1218–1225. [Google Scholar] [CrossRef]
- Alexander, R.R.; Sheppard, W.D. Picea engelmanii Parry ex Engelm. In Silvics of North America, Volume 1, Conifers; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service, Agricultural Handbook 654, USDA Forest Service: Washington, DC, USA, 1990; pp. 187–203. [Google Scholar]
- Hart, S.J.; Veblen, T.T.; Kulakowski, D. Do tree and stand-level attributes determine susceptibility of spruce-fir forests to spruce beetle outbreaks in the 21st century? For. Ecol. Manag. 2014, 318, 44–53. [Google Scholar] [CrossRef]
- Alexander, R.R.; Shearer, R.C.; Shepperd, W.D. Abies lasiocarpa (Hook.) Nutt. In Silvics of North America, Volume 1, Conifers; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service, Agricultural Handbook 654, USDA Forest Service: Washington, DC, USA, 1990; pp. 60–70. [Google Scholar]
- Hobson, E.R.; Foster, J.H. Engelmann Spruce in the Rocky Mountains; USDA Forest Service Circular 170: Washington, DC, USA, 1910; p. 23. [Google Scholar]
- Stahelin, R. Factors influencing the natural restocking of high altitude burns by coniferous trees in the central Rocky Mountains. Ecology 1943, 24, 19–30. [Google Scholar] [CrossRef]
- Wadleigh, L.; Jenkins, M.J. Fire frequency and the vegetative mosaic of a spruce-fir forest in northern Utah. Gt. Basin Nat. 1996, 56, 28–37. [Google Scholar]
- Long, J.N. The Middle and Southern Rocky Mountain. In Regional Silviculture of the United States, 3rd ed.; Barrett, J.W., Ed.; John Wiley and Sons: New York, NY, USA, 1994; pp. 335–386. [Google Scholar]
- Alexander, R.R. Partial Cutting in Old Growth Spruce-Fir; Research Paper RM-110; USDA Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1973; p. 16. [Google Scholar]
- Alexander, F.F. Cutting methods in relation to resource use in central Rocky Mountain spruce-fir forests. J. For. 1977, 75, 395–400. [Google Scholar]
- Hart, G.E.; Lomas, D.A. Effects of clearcutting on soil water depletion in an Engelmann spruce stand. Water Resour. Res. 1979, 6, 1598–1602. [Google Scholar] [CrossRef]
- Schimpf, D.J.; Henderson, J.A.; MacMahon, J.A. Some aspects of succession in the spruce-fir forest zone of Northern Utah. Gt. Basin Nat. 1980, 40, 1–26. [Google Scholar]
- Van Miegroet, H.J.; Boettinger, J.L.; Baker, M.A.; Nielsen, J.; Evans, D.; Stum, A. Soil carbon distribution and quality in a montane rangeland-forest mosaic in northern Utah. For. Ecol. Manag. 2005, 220, 284–299. [Google Scholar] [CrossRef]
- Olsen, H.R.; van Miegroet, H. Factors affecting CO2 release from forest and rangeland soils in northern Utah. Soil Sci. Soc. Am. J. 2010, 74, 282–291. [Google Scholar] [CrossRef]
- Shaw, J.D. Application of stand density index to irregularly structured stands. West. J. Appl. For. 2000, 15, 40–42. [Google Scholar]
- Long, J.N.; Daniel, T.W. Assessment of growing stock in uneven-aged stands. West. J. Appl. For. 1990, 5, 93–96. [Google Scholar]
- Reineke, L.H. Perfecting a stand-density index for even-aged forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Long, J.N. A practical approach to density management. For. Chron. 1985, 61, 23–27. [Google Scholar] [CrossRef]
- Shaw, J.D. US Forest Service Rocky Mountain Research Station & Utah State University: Logan, UT, USA, Unpublished work; 2014.
- Curtis, R.O.; Marshall, D.D. Permanent-Plot Procedures for Silvicultural and Yield Research; General Technical Report PNW-GTR—634; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2005; p. 86. [Google Scholar]
- Alexander, R.R.; Edminster, C.B. Engelmann Spruce Seed Dispersal in the Central Rocky Mountains; Research Paper RM—217; USDA Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1986; p. 14. [Google Scholar]
- Alexander, R.R. Engelmann Spruce Seed Production and Dispersal, and Seedling Establishment in the Central Rocky Mountains; General Technical Report RM—134; USDA Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1986; p. 9. [Google Scholar]
- Ducey, M.J. The ratio of additive and traditional stand density indices. West. J. Appl. For. 2009, 24, 5–10. [Google Scholar]
- Curtis, R. Effect of diameter limits and stand structure on relative density indices: A case study. West. J. Appl. For. 2010, 25, 169–175. [Google Scholar]
- Clendenen, C.W. Base-Age Conversion and Site Index Equations for Engelmann Spruce Stands in the Central and Southern Rocky Mountains; Research Note INT-223; USDA Forest Service, Intermountain Forest and Range Experiment Station: Odgen, UT, USA, 1977; p. 6. [Google Scholar]
- DeRose, R.J.; Long, J.N. Regeneration response and seedling bank dynamics on a Dendroctonus rufipennis-killed Picea engelmannii landscape. J. Veg. Sci. 2010, 21, 377–387. [Google Scholar] [CrossRef]
- Noble, D.L.; Alexander, R.R. Environmental factors affecting natural regeneration of Engelmann spruce in the Central Rocky Mountains. For. Sci. 1977, 23, 420–429. [Google Scholar]
- DeRose, R.J.; Bentz, B.J.; Long, J.N.; Shaw, J.D. Effects of increasing temperatures on the distribution of spruce beetle in Engelmann spruce forests of the Interior West, USA. For. Ecol. Manag. 2013, 308, 198–206. [Google Scholar] [CrossRef]
- Mitchell, S.J. Wind as a natural disturbance agent in forests: A synthesis. Forestry 2013, 86, 147–157. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 867–953. [Google Scholar]
- Yousefpour, R.; Jacobsen, J.B.; Thorsen, B.J.; Mieilby, H.; Hanewinkel, M.; Oehler, K. A review of decision-making approaches to handle uncertainty and risk in adaptive forest management under climate change. Ann. For. Sci. 2012, 69, 1–15. [Google Scholar] [CrossRef]
- Volney, W.J.A.; Fleming, R.A. Climate change and impacts of boreal forest insects. Agric. Ecosyst. Environ. 2000, 82, 283–294. [Google Scholar] [CrossRef]
- Flower, C.E.; Gonzalez-Meler, M.A. Responses of temperate forest productivity to insects and pathogen disturbance. Ann. Rev. Plant Biol. 2015. [Google Scholar] [CrossRef]
- Seastedt, T.R.; Hobbs, R.J.; Suding, K.N. Management of novel ecosystems: Are novel approaches required? Front. Ecol. Environ. 2008, 6, 547–553. [Google Scholar] [CrossRef]
- Miller, L.K.; Werner, R.A. Cold-hardiness of adult and larval spruce beetles Dendroctonus rufipennis (Kirby) in interior Alaska. Can. J. Zool. 1987, 65, 2927–2930. [Google Scholar] [CrossRef]
- Rousseau, J.; Bauce, E.; Lavallee, R.; Guertin, C. Winter mortality and supercooling point of the spruce beetle (Coleoptera: Curculionidae) not affected by host tree vigor in Nova Scotia, Canada. J. Acadian Entomol. Soc. 2012, 8, 1–10. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Windmuller-Campione, M.A.; Long, J.N. If Long-Term Resistance to a Spruce Beetle Epidemic is Futile, Can Silvicultural Treatments Increase Resilience in Spruce-Fir Forests in the Central Rocky Mountains? Forests 2015, 6, 1157-1178. https://doi.org/10.3390/f6041157
Windmuller-Campione MA, Long JN. If Long-Term Resistance to a Spruce Beetle Epidemic is Futile, Can Silvicultural Treatments Increase Resilience in Spruce-Fir Forests in the Central Rocky Mountains? Forests. 2015; 6(4):1157-1178. https://doi.org/10.3390/f6041157
Chicago/Turabian StyleWindmuller-Campione, Marcella A., and James N. Long. 2015. "If Long-Term Resistance to a Spruce Beetle Epidemic is Futile, Can Silvicultural Treatments Increase Resilience in Spruce-Fir Forests in the Central Rocky Mountains?" Forests 6, no. 4: 1157-1178. https://doi.org/10.3390/f6041157
APA StyleWindmuller-Campione, M. A., & Long, J. N. (2015). If Long-Term Resistance to a Spruce Beetle Epidemic is Futile, Can Silvicultural Treatments Increase Resilience in Spruce-Fir Forests in the Central Rocky Mountains? Forests, 6(4), 1157-1178. https://doi.org/10.3390/f6041157