
Response of Boreal Jack Pine (Pinus banksiana Lamb.) Stands to a Gradient of Commercial Thinning Intensities, with and without N Fertilization



Abstract
:1. Introduction
2. Experimental Section
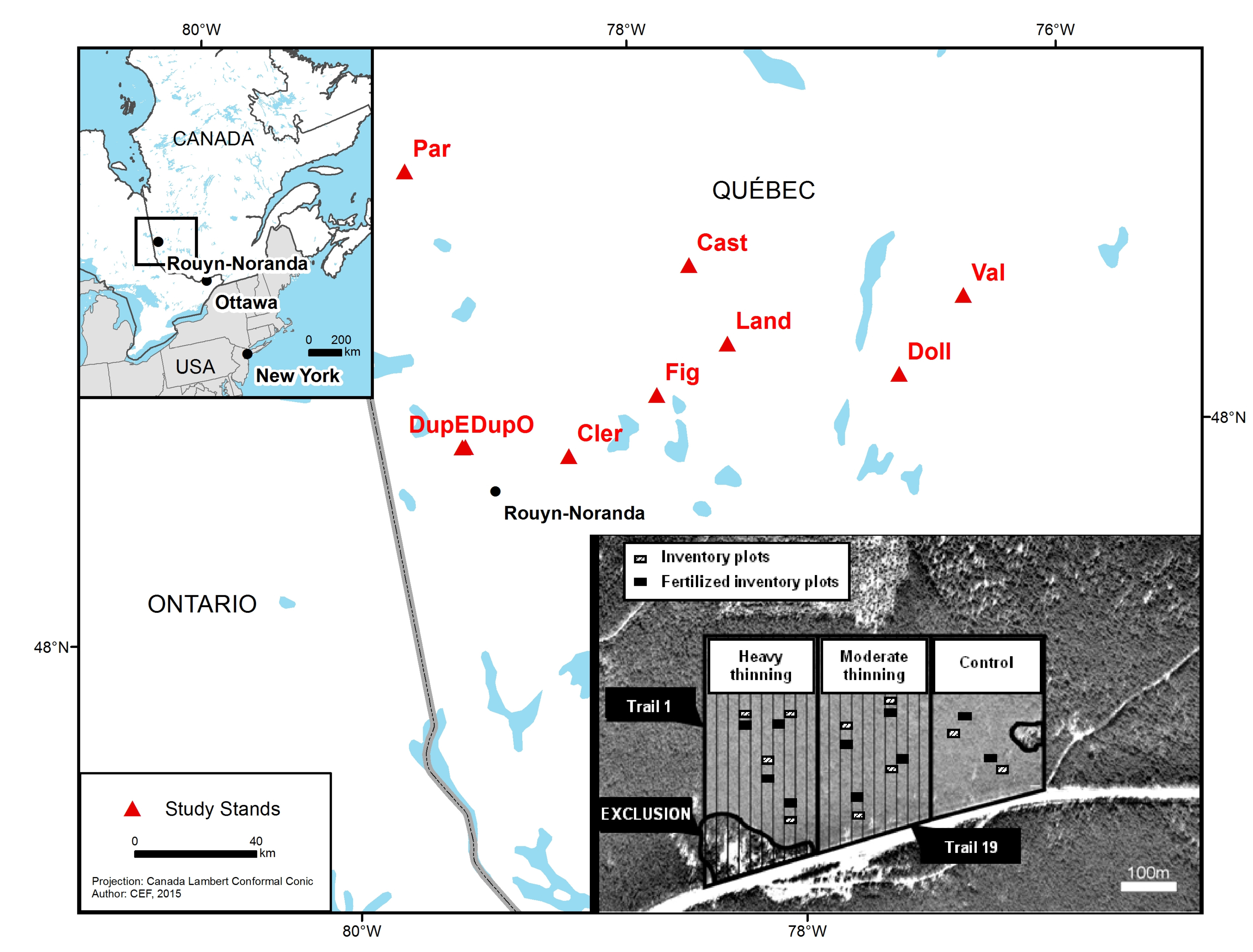
2.1. Study Area

2.2. Experimental Design

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand | Age 1 | Stand Basal Area (m2 ha−1) | Stand Density (Stems ha−1) | Thinning Intensity (%) | ||||
|---|---|---|---|---|---|---|---|---|
| Control Plots | Thinned Plots | Control Plots | Thinned Plots | Min | Mean | Max | ||
| Cast * | 48 | 24 ± 1 | 25 ± 3 | 2025 ± 177 | 2275 ± 740 | 6 | 21 | 45 |
| Cler * | 69 | 29 ± 8 | 23 ± 4 | 2125 ± 1096 | 2137 ± 226 | 37 | 49 | 64 |
| Doll * | 53 | 27 ± 4 | 23 ± 3 | 3125 ± 1025 | 3050 ± 436 | 17 | 28 | 37 |
| DupE * | 53 | 36 ± 1 | 29 ± 4 | 4125 ± 177 | 3487 ± 1122 | 26 | 41 | 52 |
| DupO | 48 | 31 ± 1 | 31 ± 4 | 1725 ± 247 | 2106 ± 607 | 16 | 29 | 40 |
| Fig * | 81 | 28 ± 1 | 28 ± 4 | 1575 ± 177 | 2106 ± 729 | 26 | 38 | 53 |
| Land | 42 | 26 ± 3 | 21 ± 4 | 1900 ± 212 | 1868 ± 356 | 21 | 27 | 35 |
| Par | 68 | 33 ± 8 | 24 ± 4 | 2750 ± 566 | 1668 ± 413 | 2 | 12 | 21 |
| Val | NA | 24 ± 3 | 29 ± 4 | 1550 ± 71 | 2081 ± 488 | 3 | 14 | 29 |
2.3. Permanent Sampling Plot Measurements
2.4. Calculations
- DBH = diameter at breast height (cm),
- SD = stump diameter (cm),
- SH = stump height (cm),
- a and b = species specific coefficients locally calibrated.
- H = height (m),
- D = diameter at breast height (cm),
- β0, β1 and β2 = specific species coefficients locally calibrated.
- V = volume (dm3),
- H = height (m),
- D = diameter at breast height (cm),
- β0, β1, β2 and β3 = species specific coefficients locally calibrated.
2.5. Statistical Analyses
3. Results
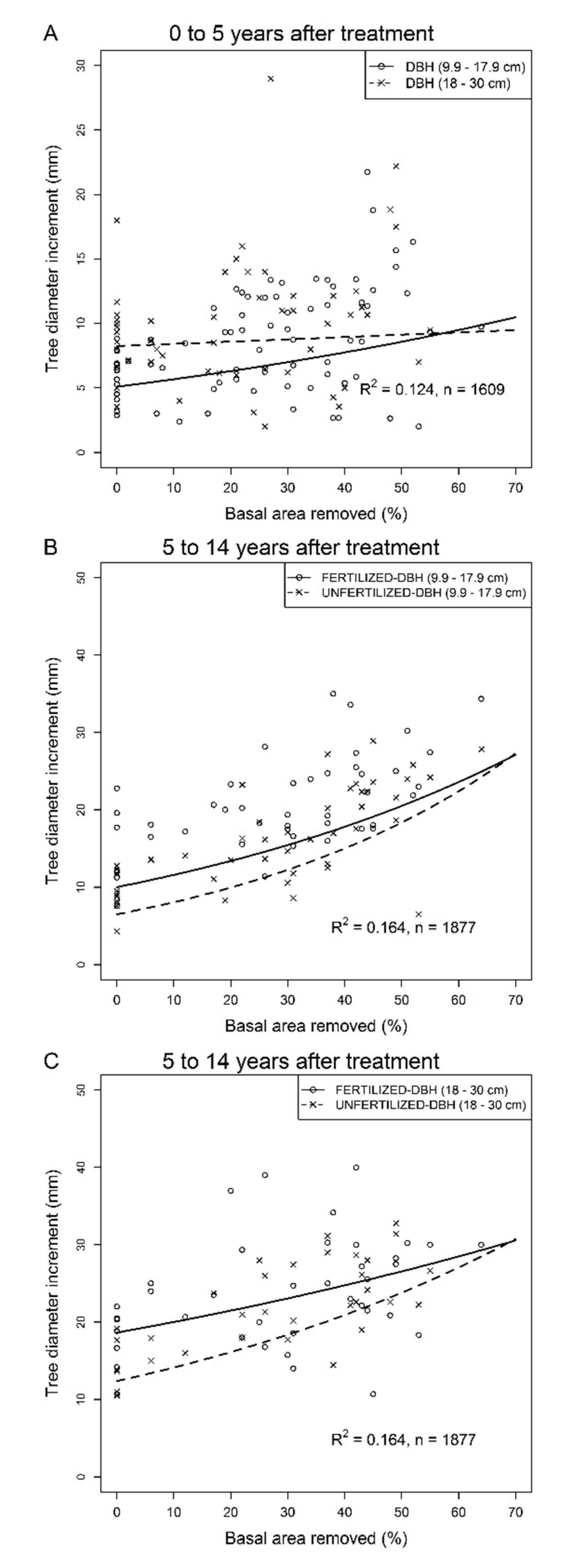
3.1. Tree Response 0 to 5 Years Following Thinning
| Response Variables | Absolute Tree Diameter Increment * | Absolute Tree Diameter Increment * | Relative Tree Volume Increment * | Mean Tree Volume | Absolute Tree Volume Increment * | Relative Tree Volume Increment * | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Period After Thinning | 5 Years | 5 to 14 Years | 5 Years | 14 Years | 5 to 14 Years | 5 to 14 Years | ||||||
| Fixed factors | E | p | E | p | E | p | E | p | E | p | E | p |
| Intercept | 2.72 | <0.001 | 3.64 | <0.001 | 2.56 | <0.001 | 112.4 | 0.043 | 4.41 | <0.001 | 3.68 | <0.001 |
| BA initial | −0.02 | 0.014 | −0.01 | 0.181 | −0.02 | 0.007 | 1.37 | 0.442 | −0.02 | 0.108 | −0.02 | 0.131 |
| BA removed | 0.01 | 0.505 | 0.01 | 0.034 | 0.01 | 0.402 | 1.25 | 0.003 | 0.01 | 0.074 | 0.01 | 0.260 |
| Size (DBH ≤ 17.9 cm) | 0.29 | 0.184 | −0.43 | 0.089 | 0.13 | 0.083 | - | −0.93 | <0.001 | 0.08 | 0.559 | |
| Age | −0.01 | 0.832 | 0.01 | 0.270 | - | - | - | |||||
| BA removed × Size | 0.01 | 0.009 | 0.01 | 0.021 | 0.01 | 0.027 | - | 0.01 | 0.018 | 0.01 | 0.012 | |
| Size × Age | −0.01 | <0.001 | −0.01 | 0.542 | - | - | - | |||||
| Fertilization (UnF) | - | −0.38 | <0.001 | - | −11.8 | 0.074 | −0.28 | 0.030 | −0.29 | 0.024 | ||
| BA removed × Fertilization | - | 0.01 | 0.007 | - | −0.33 | 0.350 | 0.01 | 0.016 | 0.01 | 0.013 | ||
| Fertilization × Size | - | - | - | - | −0.18 | 0.032 | −0.24 | 0.046 | ||||

3.2. Tree Response 5 to 14 Years Following Thinning in Unfertilized and Fertilized Plots


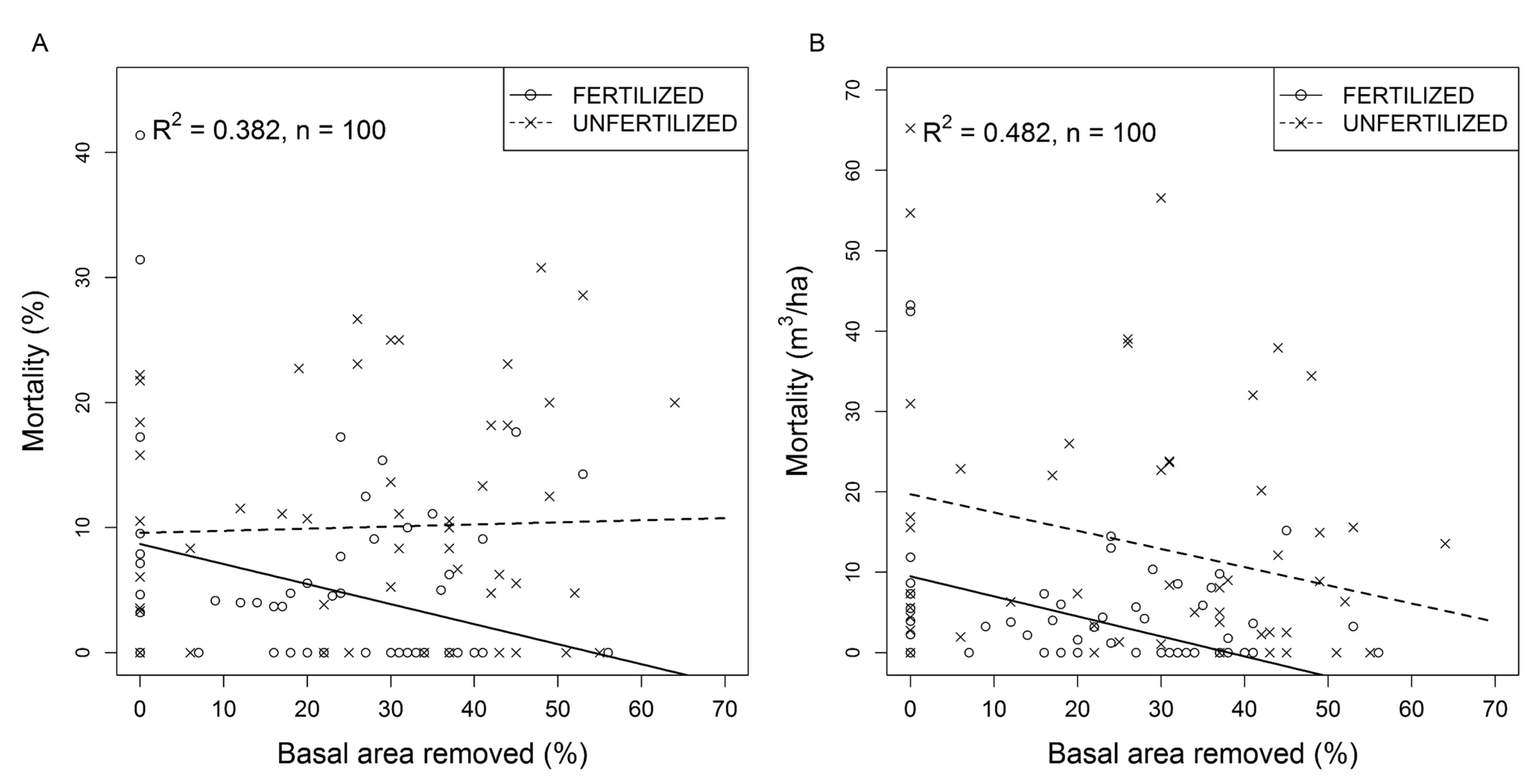
3.3. Tree Mortality in Response to Thinning and Fertilization
| Response Variable | Tree Mortality (% of Residual Stems) | Tree Mortality (% of Residual Stems) | Volume Lost to Mortality | Volume Lost to Mortality | ||||
|---|---|---|---|---|---|---|---|---|
| Period After Thinning | 5 Years | 5 to 14 Years | 5 Years | 5 to 14 Years | ||||
| Fixed factors | E | p | E | p | E | p | E | p |
| Intercept | −10.8 | 0.008 | −19.9 | <0.001 | −9.9 | 0.136 | −35.9 | <0.001 |
| BA initial | 0.001 | 0.993 | 0.35 | 0.022 | 0.31 | 0.130 | 0.49 | 0.022 |
| BA removed | 0.08 | 0.016 | −0.16 | 0.027 | −0.01 | 0.879 | −0.24 | 0.001 |
| Age | 0.21 | 0.004 | 0.33 | 0.015 | 2.3 | 0.029 | 0.56 | 0.010 |
| Fertilization (UnF) | - | 0.89 | 0.722 | - | 10.8 | <0.001 | ||
| BA removed × Fertilization (UnF) | - | 0.17 | 0.037 | - | 0.02 | 0.838 | ||

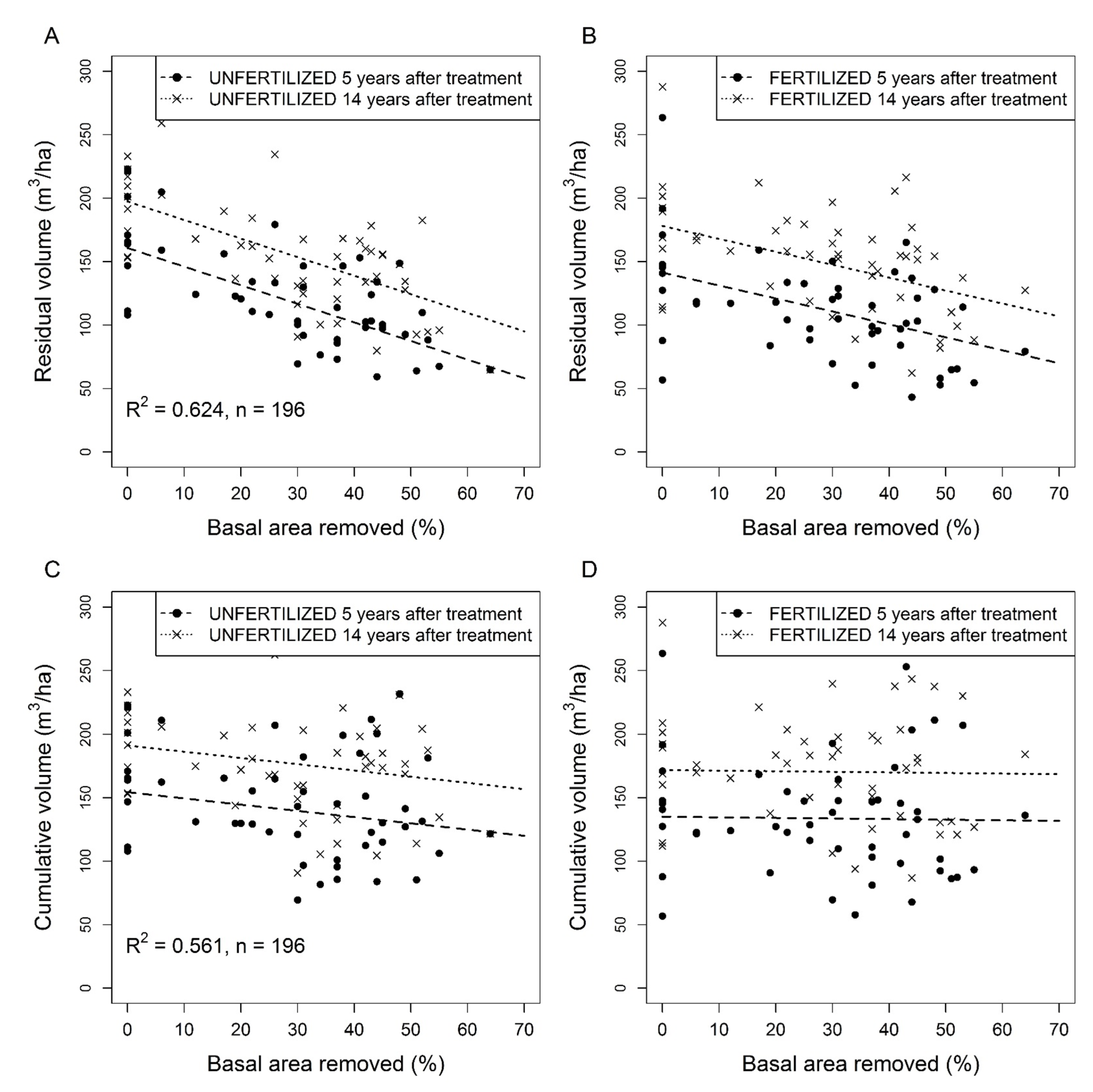
3.4. Stand Basal Area and Volume Increment Following Thinning with and without Fertilization
| Response Variables | Absolute Basal Area Increment | Absolute Basal Area Increment | Absolute Volume Increment | Absolute Volume Increment | Relative Volume Increment | Relative Volume Increment | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Period After Thinning | 5 Years | 5 to 14 Years | 5 Years | 5 to 14 Years | 5 Years | 5 to 14 Years | ||||||
| Fixed factors | E | p | E | p | E | p | E | p | E | p | E | p |
| Intercept | 1.82 | 0.103 | 3.91 | <0.001 | 17.79 | 0.101 | 62.1 | <0.001 | 38.5 | 0.006 | 85.6 | <0.001 |
| BA initial | 0.05 | 0.001 | 0.12 | <0.001 | 0.42 | 0.061 | 0.94 | 0.008 | −0.30 | 0.234 | −0.16 | 0.672 |
| BA removed | −0.01 | 0.134 | 0.01 | 0.837 | −0.04 | 0.505 | 0.20 | 0.060 | 0.22 | 0.001 | 0.41 | 0.001 |
| Age | −0.02 | 0.378 | −0.04 | 0.024 | −0.17 | 0.319 | −0.83 | 0.006 | −0.29 | 0.199 | −0.83 | 0.018 |
| Fertilization (UnF) | - | −0.79 | 0.050 | −8.33 | 0.096 | - | −11.5 | 0.024 | ||||
| BA removed × Fertilization | - | −0.01 | 0.338 | −0.05 | 0.729 | - | −0.06 | 0.666 | ||||


| Response Variables | Residual Live Volume | Cumulative Volume | ||
|---|---|---|---|---|
| Fixed factors | E | p | E | p |
| Intercept | 14.3 | 0.646 | −75.8 | 0.073 |
| BA initial | 3.33 | <0.001 | 5.20 | <0.001 |
| BA removed | −1.02 | <0.001 | −0.05 | 0.830 |
| Age | 0.68 | 0.187 | 1.28 | 0.115 |
| Time since thinning (14 years) | 36.8 | <0.001 | 36.8 | <0.001 |
| BA removed × time (14 years) | 0.01 | 0.990 | 0.01 | 0.986 |
| Fertilization (UnF) | 19.3 | <0.001 | 19.3 | <0.001 |
| BA removed × Fertilization | −0.44 | 0.008 | −0.44 | 0.008 |
4. Discussion
4.1. Tree Growth
4.2. Mortality
4.3. Fertilization Effect on Tree Growth
4.4. Fertilization and Tree Mortality
4.5. Stand Volume Increment and Cumulative Volume
5. Conclusions
Supplementary Material

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pothier, D.; Margolis, A. Analysis of growth and light interception of balsam fir and white birch saplings following precommercial thinning. Ann. For. Sci. 1991, 48, 123–132. [Google Scholar] [CrossRef]
- Karsh, M.B.; Lavigne, M.B.; Donnelly, J.G. Growth Responses of the Balsam Fir and Black Spruce Spacing Trials; Information Report N-X-291; Natural Resources Canada, Atlantic Forestry Centre: St-John’s, NF, Canada, 1994.
- Smith, D.M.; Larson, B.C.; Kelty, M.J.; Ashton, P.M.S. The Practice of Silviculture: Applied Forest Ecology, 9th ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1997. [Google Scholar]
- Schneider, R.; Zhang, S.Y.; Swift, D.E.; Bégin, J.; Lussier, J.M. Predicting selected wood properties of jack pine following commercial thinning. Can. J. For. Res. 2008, 38, 2030–2043. [Google Scholar] [CrossRef]
- Zeide, B. Thinning and growth: A full turn around. J. For. 2001, 99, 20–25. [Google Scholar]
- Schneider, R. Effet de L’éclaircie Commerciale sur la Croissance et la Qualité du bois du pin Gris. Ph.D. Thesis, Université Laval, Québec, QC, Canada, 2007. [Google Scholar]
- Mäkinen, H.; Isomäki, A. Thinning intensity and long-term changes in increment and stem form of Scots pine trees. For. Ecol. Manag. 2004, 203, 21–34. [Google Scholar] [CrossRef]
- Mäkinen, H.; Isomäki, A. Thinning intensity and growth of Scots pine stands in Finland. For. Ecol. Manag. 2004, 201, 311–325. [Google Scholar] [CrossRef]
- Allen, H.L. Silvicultural treatments to enhance productivity. In The Forests Handbook: Volume II; Evans, J., Ed.; Blackwell Science Ltd: Oxford, UK, 2001; p. 382. [Google Scholar]
- Curtis, R.O.; Marshall, D.D. Levels-of-Growing-Stock Cooperative Study in Douglas Fir: Report No. 14-Stampede Creek: 30 Year Results; Research Paper PNW-RP-543; USDA Forest Service: Portland, OR, USA, 2002; p. 77.
- Ontario Ministry of Natural Resources (OMNR). Provincial Wood Supply Strategy; Queen’s Printer for Ontario: Toronto, ON, Canada, 2004. [Google Scholar]
- Boulay, E. Ressources et industries forestières. Portrait statistique. Édition 2013. Available online: http://www.mffp.gouv.qc.ca/publications/forets/connaissances/portrait-statistique-2013.pdf (accessed on 10 June 2014).
- Cameron, A.D. Importance of early thinning in the development of long-term stand stability and improved log quality: A review. Forestry 2002, 75, 25–35. [Google Scholar] [CrossRef]
- Bella, I.E.; de Franceschi, J.P. Commercial Thinning Improves Growth of Jack Pine; Information Report NOR-X-112; Natural Resources Canada, Northern Forest Research Centre: Edmonton, AB, Canada, 1974; p. 23.
- Smith, C.R.; Oerlemans, W.J.A.M. Ten-Year Growth Response and Financial Evaluation of Commercial Strip Thinning of Jack Pine: A Case Study; Information Report O-X-396; Canadian Forestry Service, Great Lakes Forestry Centre: Sault Ste. Marie, ON, Canada, 1988; p. 25.
- Barbour, R.J.; Fayle, D.C.F.; Chauret, G.; Cook, J.; Karsh, M.B.; Ran, S. Breast-heigth relative density and radial growth in mature jack pine (Pinus bansksiana) for 38-years after thinning. Can. J. For. Res. 1994, 24, 2439–2447. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Chauret, G.; Swift, D.E.; Duchesne, L. Effects of pre-commercial thinning on tree growth and lumber quality in a jack pine stand in New Brunswick, Canada. Can. J. For. Res. 2006, 36, 945–952. [Google Scholar] [CrossRef]
- Yang, R.C. Foliage and stand growth responses of semimature lodgepole pine to thinning and fertilization. Can. J. For. Res. 1998, 28, 1794–1804. [Google Scholar] [CrossRef]
- Tremblay, S.; Laflèche, V. Résultats Obtenus 5 ans après Traitement dans les Placettes du Réseau de la Mesure des Effets Réels de L’éclaircie Commerciale en Peuplements Résineux. Mémoire de Recherche Forestière n°166. Direction de la Recherche Forestière; Ministère des ressources naturelles du Québec: Québec, QC, Canada, 2012; p. 52. (In French)
- Moorhead, D.J.; Dickens, E.D. Fertilization Pine Plantations. A County Agent’s Guide for Making Fertilization Recommendations; Warnell School of Forest Ressources, the University of Georgia: Athens, GA, USA, 2002. Available online: http://bugwood.org/fertilization/csoillab.html (accessed on 10 March 2015).
- Weetman, G.F.; Krause, H.H.; Koller, E.; Veilleux, J.M. Interprovincial Fertilization Program, 1968–1983: Standardized Fertilization Installations in 81 Unmanaged Middle-Aged Stands in Seven Provinces; Final Report: Results of Ten-Year Growth Measurements; Information Report DPC-X-21; Canadian Forestry Service: Ottawa, ON, Canada, 1987.
- Weetman, G.F.; Aigar, D. Jack pine nitrogen fertilization and nutrition studies: Three year results. Can. J. For. Res. 1974, 4, 381–398. [Google Scholar] [CrossRef]
- Morrison, I.K.; Swan, H.S.D.; Foster, N.W.; Winston, D.A. Ten-year growth in two fertilization experiments in a semimature jack pine stand in northwestern Ontario. For. Chron. 1997, 53, 142–146. [Google Scholar] [CrossRef]
- Bernier, C.; Camiré, B. Fertilization azotée en forêt de pin gris (Pinus banksiana) II. Variations de la masse foliaire et des éléments nutritifs du feuillage de pin gris et de quelques espèces du sous-bois. Can. J. For. Res. 1981, 11, 423–432. (In French) [Google Scholar]
- Newton, P.F.; Amponsah, I.G. Systematic review of short-term growth responses of semi-mature black spruce and jack pine stands to nitrogen-based fertilization treatments. For. Ecol. Manag. 2006, 237, 1–14. [Google Scholar] [CrossRef]
- Groot, A.; Brown, K.M.; Morrison, I.K.; Barker, J.E. A 10-year tree and stand response of jack pine to urea fertilization and low thinning. Can. J. For. Res. 1984, 14, 44–50. [Google Scholar] [CrossRef]
- Valinger, E. Effect of thinning and nitrogen fertilization on stem growth and stem form of Pinus sylvestris trees. Can. J. For. Res. 1992, 7, 219–228. [Google Scholar]
- Valinger, E.; Elfving, B.; Morling, T. Twelve-year growth response of Scots pine to thinning and nitrogen fertilization. For. Ecol. Manag. 2000, 134, 45–53. [Google Scholar] [CrossRef]
- Eriksson, E. Thinning operation and their impact on biomass production in stands of Norway spruce and Scots pine. Biomass Bioenergy 2006, 30, 848–854. [Google Scholar] [CrossRef]
- Brix, H. Effects of thinning and fertilization on growth of Douglas-fir: Relative contribution of foliage quantity and efficiency. Can. J. For. Res. 1983, 13, 167–175. [Google Scholar] [CrossRef]
- Brix, H.; Mitchell, A.K. Effects of thinning and nitrogen fertilization on xylem development in Douglas-fir. Can. J. For. Res. 1980, 10, 121–128. [Google Scholar] [CrossRef]
- DeBell, D.S.; Harrington, C.A.; Shumway, J. Thinning Shock and Response to Fertilizer Less than Expected in Young Douglas-fir Stand at Wind River Experimental Forest; USDA Forest Service Res. Paper PNW-RP-547; Pacific Northwest Research Station: Portland, OR, USA, 2002; p. 20. [Google Scholar]
- Cown, D.J.; McConchie, D.L. Effects of thinning and fertilizer application on wood properties of Pinus radiala. N. Z. For. Sci. 1981, 11, 79–91. [Google Scholar]
- Van Cleve, K.; Zasada, J. Response of 70-yr-old white spruce to thinning and fertilization in interior Alaska. Can. J. For. Res. 1976, 6, 145–152. [Google Scholar] [CrossRef]
- Weetman, G.F.; Roberge, M.R.; Meng, C.H. Black spruce: 15-year growth and microbiological response to thinning and fertilization. Can. J. For. Res. 1980, 10, 502–509. [Google Scholar] [CrossRef]
- Hale, S.E. The effect of thinning intensity on the below-canopy light environment in a Sitka spruce plantation. For. Ecol. Manag. 2003, 179, 341–349. [Google Scholar] [CrossRef]
- Latham, P.; Tappeiner, J. Response of old growth conifers to reduction in stand density in western Oregon forests. Tree Physiol. 2002, 22, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.F.; Coates, K.D.; Canham, C.D.; Bartemucci, P. Species variability in growth response to light across climatic regions in northwestern British Columbia. Can. J. For. Res. 1998, 28, 871–886. [Google Scholar] [CrossRef]
- Weiner, J. Size hierarchies in experimental populations of annual plants. Ecology 1985, 66, 743–752. [Google Scholar] [CrossRef]
- Weiner, J. How competition for light and nutrients affects size variability in Ipomea tricolor populations. Ecology 1986, 67, 1425–1427. [Google Scholar] [CrossRef]
- Del Río, M.; Condés, S.; Pretzsch, H. Analyzing size-symmetric vs. size-asymmetric and intra- vs. inter-specific competition in beech (Fagus sylvatica L.) mixed stands. For. Ecol. Manag. 2014, 325, 90–98. [Google Scholar] [CrossRef]
- Zhang, J.; Oliver, W.W.; Powers, R.F. Long-term effects of thinning and fertilization on growth of red fir in northeastern California. Can. J. For. Res. 2005, 35, 1285–1293. [Google Scholar] [CrossRef]
- Veillette, J.J. Evolution and paleohydrology of glacial Lakes Barlow and Ojibway. Quat. Sci. Rev. 1994, 13, 945–971. [Google Scholar] [CrossRef]
- Environment Canada. Available online: http://climat.meteo.gc.ca/climate_normals/index_f.html (accessed on 18 February 2014).
- Rowe, J.S. Forest Regions of Canada Publication No. 1300; Environment Canada, Canadian Forest Service: Ottawa, ON, Canada, 1972; p. 172.
- Saucier, J.-P.; Robitaille, A.; Grondin, P.; Bergeron, J.-F.; Gosselin, J. Les Régions Écologiques du Québec Méridional (4ème version). Carte à L’échelle de 1/1250000; Ministère des Ressources naturelles et de la Faune du Québec: Québec, Canada, 2011. (In French)
- Goudiaby, V.; Brais, S.; Berninger, F.; Schneider, R. Vertical patterns in specific volume increment along stems of dominant jack pine (Pinus banksiana) and black spruce (Picea mariana) after thinning. Can. J. For. Res. 2012, 42, 733–748. [Google Scholar] [CrossRef]
- Alemdag, I.S.; Honer, T.G. Metric Relationship between Breast-Height and Stump Diameters for Eleven Tree Species from Eastern and Central Canada; Canadian Forestry Service Information report FMR-X-49M; Forest Management Institute: Ottawa, ON, Canada, 1977; p. 26. [Google Scholar]
- Perron, J.-Y. Tarif de Cubage Général—Volume Marchand Brut, 3rd ed.Gouvernement du Québec, Ministère des Ressources Naturelles de la Faune et des Parcs: Québec, QC, Canada, 2003; p. 60. (In French)
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; the R Core team. The Nlme Package. 2008. Available online: http://cran.stat.sfu.ca/ (accessed on 10 October 2014).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Sprugel, D.G. Correcting for bias in log-transformed allometric equations. Ecology 1983, 64, 209–210. [Google Scholar] [CrossRef]
- Tarroux, E.; DesRochers, A.; Krause, C. Effect of natural root grafting on growth response of jack pine (Pinus banksiana) after commercial thinning. For. Ecol. Manag. 2010, 260, 526–535. [Google Scholar] [CrossRef]
- Pape, R. Influence of thinning and tree diameter class on the development of basic density and annual ring width in Picea abies. Scand. J. For. Res. 1999, 14, 27–37. [Google Scholar] [CrossRef]
- Aussenac, G.; Granier, A. Effects of thinning on water stress and growth in Douglas-fir. Can. J. For. Res. 1998, 18, 100–105. [Google Scholar] [CrossRef]
- Vincent, M.; Krause, C.; Zhang, S.Y. Radial growth response of black spruce roots and stems to commercial thinning in the boreal forest. Forestry 2009, 82, 557–571. [Google Scholar] [CrossRef]
- Mäkinen, H.; Isomäki, A. Thinning intensity and growth of Norway spruce stands in Finland. Forestry 2004, 77, 349–364. [Google Scholar] [CrossRef]
- Mäkinen, H.; Isomäki, A. Thinning intensity and long-term changes in increment and stem form of Norway spruce trees. For. Ecol. Manag. 2004, 201, 295–309. [Google Scholar] [CrossRef]
- Moore, J.A.; Zhang, L.; Newberry, J.D. Effects of intermediate silvicultural treatments on the distribution of within-stand growth. Can. J. For. Res. 1994, 24, 398–404. [Google Scholar] [CrossRef]
- Pukkala, T.; Miina, J.; Kellomäki, S. Response to different thinning intensities in young Pinus sylvestris. Scand. J. For. Res. 1998, 13, 141–150. [Google Scholar] [CrossRef]
- Mäkinen, H.; Hynynen, J.; Isomäki, A. Intensive management of Scots pine stands in southern Finland: First empirical results and simulated further development. For. Ecol. Manag. 2005, 215, 37–50. [Google Scholar] [CrossRef]
- Kuusipalo, J. On the use of tree stand parameters in estimating light conditions below the canopy. Silva Fenn. 1985, 19, 185–196. [Google Scholar] [CrossRef]
- Comeau, P. Measuring Light in the Forest; Extension Note 42; Ministry of Forests Research Program: Victoria, BC, Canada, 2000; p. 7.
- Hale, S.E. Light regime beneath Sitka spruce plantations in northern Britain: Preliminary results. For. Ecol. Manag. 2001, 151, 61–66. [Google Scholar] [CrossRef]
- Carlyle, J.C. Nutrient management in a Pinus radiata plantation after thinning: The effect of thinning and residues on nutrient distribution, mineral nitrogen fluxes, and extractable phosphorus. Can. J. For. Res. 1995, 25, 1278–1291. [Google Scholar] [CrossRef]
- Thibodeau, L.; Raymond, P.; Camiré, C.; Munson, A.D. Impact of precommercial thinning in balsam fir stands on soil nitrogen dynamics, microbial biomass, decomposition, and foliar nutrition. Can. J. For. Res. 2000, 30, 229–238. [Google Scholar] [CrossRef]
- Pothier, D. Twenty-year results of precommercial thinning in balsam fir stand. For. Ecol. Manag. 2002, 168, 177–186. [Google Scholar] [CrossRef]
- Ryan, M.G.; Binkley, D.; Fownes, J.H. Age-related decline in forest productivity: Pattern and process. Adv. Ecol. Res. 1997, 27, 213–262. [Google Scholar]
- Skovsgaard, J.P.; Vanclay, J.K. Forest site productivity: A review of the evolution of dendrometric concepts for even-aged stands. Forestry 2008, 81, 13–31. [Google Scholar] [CrossRef]
- Kenkel, N.C.; Hoskins, J.A.; Hoskins, W.D. Local competition in a naturally established jack pine stand. Can. J. Bot. 1989, 67, 2630–2635. [Google Scholar] [CrossRef]
- Peet, R.K.; Christensen, N.L. Competition and tree death. BioScience 1987, 37, 586–595. [Google Scholar] [CrossRef]
- Elfving, B. Natural mortality in thinning and fertilization experiments with pine and spruce in Sweden. For. Ecol. Manag. 2010, 260, 353–360. [Google Scholar] [CrossRef]
- Ruel, J.C. Understanding windthrow: Silvicultural implications. For. Chron. 1995, 71, 434–445. [Google Scholar] [CrossRef]
- Scott, R.E.; Mitchell, S.J. Empirical modelling of windthrow risk in partially harvested stands using tree, neighbourhood, and stand attributes. For. Ecol. Manag. 2005, 218, 193–209. [Google Scholar] [CrossRef]
- Mason, W.L.; Kerr, G. Transforming Even-Aged Conifer Stands to Continuous Cover Management; Information note 40; Forestry Commission: Edinburgh, UK, 2001.
- Helmisaari, H.S.; Hanssen, K.H.; Jacobson, S.; Kukkola, M.; Luiro, J.; Saarsalmi, A.; Tamminen, P.; Tveite, B. Logging residue removal after thinning in Nordic boreal forests: Long-term impact on tree growth. For. Ecol. Manag. 2011, 261, 1919–1927. [Google Scholar] [CrossRef]
- Del Rio, M.; Calama, R.; Canellas, I.; Roig, S.; Montero, G. Thinning intensity and growth response in SW-European Scots pine stands. Ann. For. Sci. 2008, 65, 308. [Google Scholar] [CrossRef]
- Assmann, E. The Principles of Forest Yield Study; Pergamon Press: Oxford, UK, 1970. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moulinier, J.; Brais, S.; Harvey, B.D.; Koubaa, A. Response of Boreal Jack Pine (Pinus banksiana Lamb.) Stands to a Gradient of Commercial Thinning Intensities, with and without N Fertilization. Forests 2015, 6, 2678-2702. https://doi.org/10.3390/f6082678
Moulinier J, Brais S, Harvey BD, Koubaa A. Response of Boreal Jack Pine (Pinus banksiana Lamb.) Stands to a Gradient of Commercial Thinning Intensities, with and without N Fertilization. Forests. 2015; 6(8):2678-2702. https://doi.org/10.3390/f6082678
Chicago/Turabian StyleMoulinier, Julien, Suzanne Brais, Brian D. Harvey, and Ahmed Koubaa. 2015. "Response of Boreal Jack Pine (Pinus banksiana Lamb.) Stands to a Gradient of Commercial Thinning Intensities, with and without N Fertilization" Forests 6, no. 8: 2678-2702. https://doi.org/10.3390/f6082678
APA StyleMoulinier, J., Brais, S., Harvey, B. D., & Koubaa, A. (2015). Response of Boreal Jack Pine (Pinus banksiana Lamb.) Stands to a Gradient of Commercial Thinning Intensities, with and without N Fertilization. Forests, 6(8), 2678-2702. https://doi.org/10.3390/f6082678