
Soil Nitrogen Transformations and Availability in Upland Pine and Bottomland Alder Forests


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites

{kind=link}
{kind=link}
{kind=link}
| Upland Pine Forest (UPF) | Bottomland Alder Forest (BAF) | ||||
|---|---|---|---|---|---|
| Low Density (LD) | High Density (HD) | Low Moisture (LM) | Middle Moisture (MM) | High Moisture (HM) | |
| Site (1980–2010 average for climate data) | |||||
| Location | 37°47′01″ N, 127°10′37″ E Rural area 30 km north of Seoul | 37°27′52″ N, 127°04′53″ E Southern suburb of Seoul | |||
| Altitude (m) | 410–440 | 40 | |||
| Aspect, slope (°) | West, 13–22 | South, <1 | |||
| Temperature (°C) | 11.4 (monthly range: −5.0–26.5) | 13.5 (monthly range: −1.9–27.2) | |||
| Annual precipitation (mm) | 1471 | 1473 | |||
| Vegetation (in 2007 for the UPF and in 2009 for the BAF) | |||||
| Canopy species | Pinus densiflora | Alnus japonica | |||
| Density (trees·ha−1) | 983 | 1517 | 900 | 575 | 550 |
| Diameter at breast height (cm) | 20.5 | 17.6 | 32.6 | 27.1 | 24.2 |
| Aboveground biomass (Mg C·ha−1) | 88.1 | 86.7 | 187.9 | 78.9 | 61.8 |
| Needle or leaf litter C:N ratio | 72.7 (2.0) | 73.2 (2.2) | 21.8 (0.3) | 20.8 (0.2) | 18.6 (0.2) |
| Total N storage (Mg N·ha−1) | 0.78 | 0.78 | 1.65 | 0.74 | 0.60 |
| Soil (0–15 cm; during the study period) | |||||
| Soil texture | Silt loam | Silt loam | Sandy loam | Sandy loam | Silt loam |
| Bulk density (g·cm−3) | 1.05 (0.04) | 1.02 (0.03) | 0.86 (0.06) | 0.81 (0.03) | 0.50 (0.14) |
| Soil water content (kg·kg−1) | 0.25 (0.02) | 0.20 (0.03) | 0.35 (0.06) | 0.72 (0.17) | 1.23 (0.10) |
| pH | 4.54 (0.03) | 4.56 (0.01) | 4.45 (0.07) | 5.04 (0.10) | 5.14 (0.04) |
| Total C concentration (g·kg−1) | 33.4 (2.3) | 29.7 (2.1) | 20.8 (2.0) | 31.2 (1.8) | 61.7 (2.3) |
| C:N ratio | 15.8 (0.4) | 17.8 (0.4) | 9.6 (0.3) | 12.4 (0.2) | 12.3 (0.1) |
| Total N storage (0–30 cm; Mg N·ha−1) | 3.62 (0.24) | 2.98 (0.12) | 4.64 (0.52) | 5.06 (0.70) | 7.14 (0.20) |
2.2. Field Experiment and Laboratory Analysis
2.3. Statistical Analysis
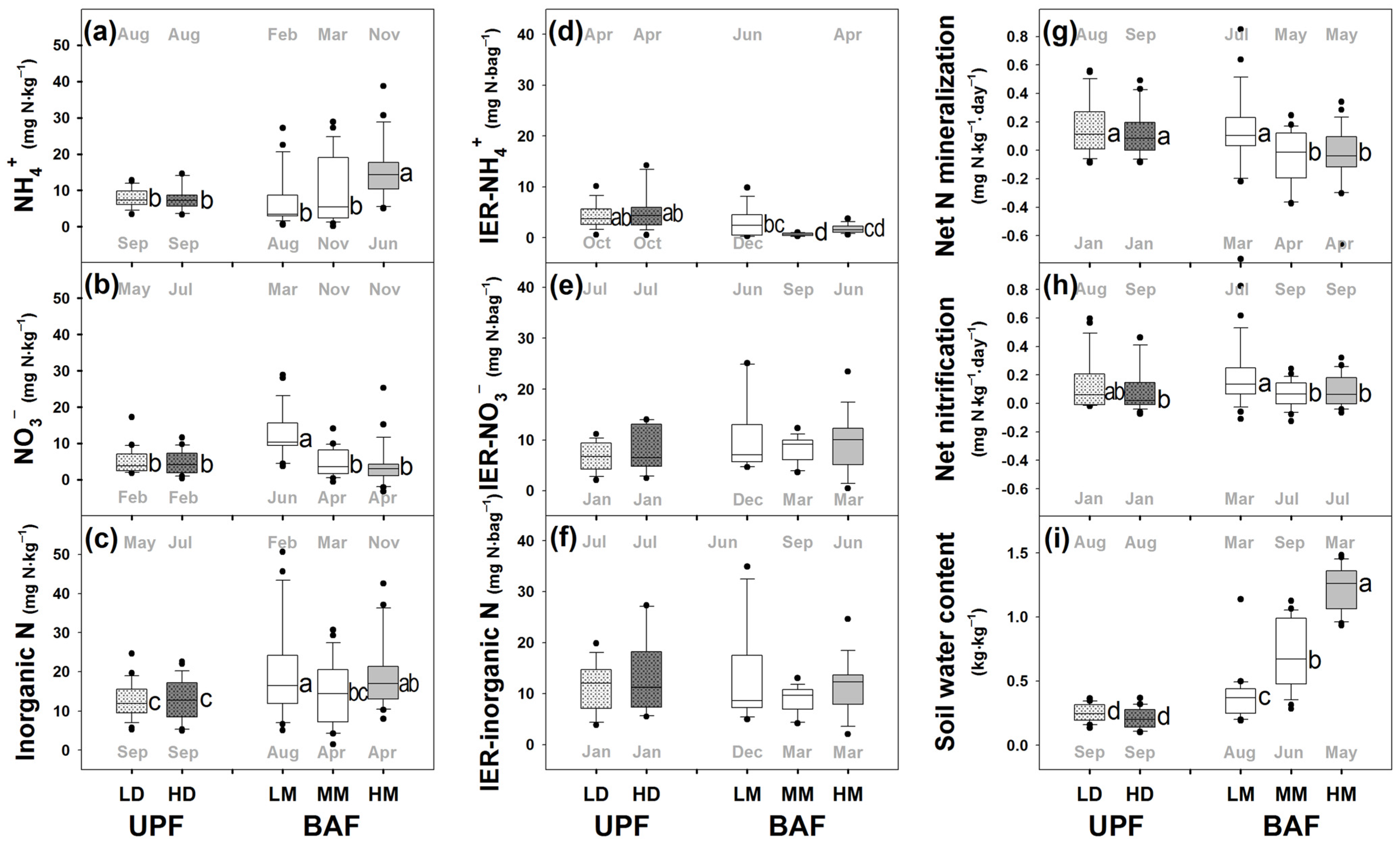
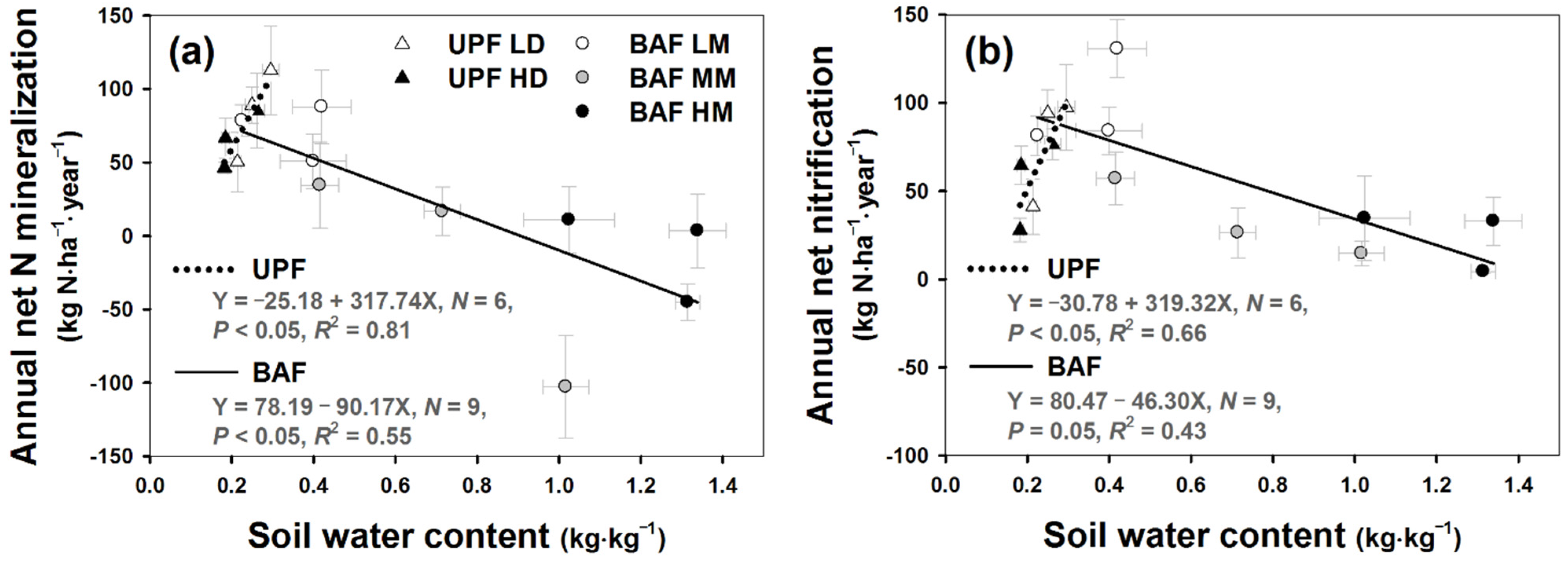
3. Results

| Soil Temperature | Soil Water Content | |||
|---|---|---|---|---|
| UPF | BAF | UPF | BAF | |
| NH4+ | 0.31 | – | 0.54 | 0.57 |
| NO3− | – | – | – | −0.48 |
| Inorganic N | 0.34 | – | 0.43 | – |
| IER-NH4+ | – | 0.37 | – | – |
| IER-NO3− | 0.71 | 0.46 | – | – |
| IER-inorganic N | 0.57 | 0.47 | – | – |
| Net N mineralization | 0.58 | 0.24 | 0.56 | −0.37 |
| Net nitrification | 0.67 | 0.32 | 0.60 | −0.29 |

| Annual Net N Mineralization Rate † | Annual Net Nitrification Rate † | IER-NH4+ ‡ | IER-NO3− ‡ | IER-Total Inorganic N ‡ | Annual N Uptake † | ANPP § | |
|---|---|---|---|---|---|---|---|
| UPF | |||||||
| LD | 84.01 ± 18.18 | 77.61 ± 18.26 | 4.29 ± 0.49 | 6.81 ± 0.58 | 11.10 ± 0.97 | 73.19 | 6.17 |
| HD | 66.05 ± 11.25 | 56.26 ± 14.56 | 5.17 ± 0.79 | 8.17 ± 0.88 | 13.34 ± 1.53 | 60.18 | 5.03 |
| BAF | |||||||
| LM | 72.24 ± 11.13 | 98.74 ± 16.01 | 3.03 ± 0.59 | 10.73 ± 1.43 | 13.76 ± 1.98 | 191.77 | 5.01 |
| MM | −17.22 ± 43.10 | 32.69 ± 12.67 | 0.66 ± 0.05 | 8.14 ± 0.52 | 8.80 ± 0.54 | 141.21 | 6.06 |
| HM | −10.26 ± 17.60 | 23.98 ± 9.80 | 1.75 ± 0.17 | 9.62 ± 1.13 | 11.37 ± 1.07 | 128.76 | 4.51 |

4. Discussion
4.1. Restricted Net N Mineralization under NH4+ Rich Condition
4.2. Discrepancy among Inorganic N in Bulk Soil and IER, and Net N Transformation Rates: Which N Indices Could Describe the Real Status of N Availability?
4.3. Stand Density Effects on Soil N Transformation and Availability
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Binkley, D.; Vitousek, P. Soil nutrient availability. In Plant Physiological Ecology; Pearcy, R., Ehleringer, J., Mooney, H., Rundel, P., Eds.; Chapman and Hall Ltd.: New York, NY, USA, 1989; pp. 75–96. [Google Scholar]
- Vitousek, P.M.; Howarth, R.W. Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry 1991, 13, 87–115. [Google Scholar] [CrossRef]
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Saunders, D.L.; Kalff, J. Nitrogen retention in wetlands, lakes and rivers. Hydrobiologia 2001, 443, 205–212. [Google Scholar] [CrossRef]
- Cameron, K.C.; Di, H.J.; Moir, J.L. Nitrogen losses from the soil/plant system: A review. Ann. Appl. Biol. 2013, 162, 145–173. [Google Scholar] [CrossRef]
- Binkley, D.; Hart, S.C. The components of nitrogen availability assessments in forest soils. In Advances in Soil Science; Stewart, B.A., Ed.; Springer: New York, NY, USA, 1989; Volume 10, pp. 57–112. [Google Scholar]
- Schimel, J.P.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Frank, D.A.; Groffman, P.M. Plant rhizospheric N processes: What we don’t know and why we should care. Ecology 2009, 90, 1512–1519. [Google Scholar] [CrossRef] [PubMed]
- Ros, G.H.; Temminghoff, E.J.M.; Hoffland, E. Nitrogen mineralization: A review and meta-analysis of the predictive value of soil tests. Eur. J. Soil Sci. 2011, 62, 162–173. [Google Scholar] [CrossRef]
- Zhu, B.; Gutknecht, J.L.M.; Herman, D.J.; Keck, D.C.; Firestone, M.K.; Cheng, W. Rhizosphere priming effects on soil carbon and nitrogen mineralization. Soil Biol. Biochem. 2014, 76, 183–192. [Google Scholar] [CrossRef]
- Manzoni, S.; Porporato, A. Soil carbon and nitrogen mineralization: Theory and models across scales. Soil Biol. Biochem. 2009, 41, 1355–1379. [Google Scholar] [CrossRef]
- Paul, K.I.; Polglase, P.J.; O'Connell, A.M.; Carlyle, J.C.; Smethurst, P.J.; Khanna, P.K. Defining the relation between soil water content and net nitrogen mineralization. Eur. J. Soil Sci. 2003, 54, 39–48. [Google Scholar] [CrossRef]
- Sierra, J. Temperature and soil moisture dependence of n mineralization in intact soil cores. Soil Biol. Biochem. 1997, 29, 1557–1563. [Google Scholar] [CrossRef]
- Guntiñas, M.E.; Leirós, M.C.; Trasar-Cepeda, C.; Gil-Sotres, F. Effects of moisture and temperature on net soil nitrogen mineralization: A laboratory study. Eur. J. Soil Biol. 2012, 48, 73–80. [Google Scholar] [CrossRef]
- Sleutel, S.; Moeskops, B.; Huybrechts, W.; Vandenbossche, A.; Salomez, J.; De Bolle, S.; Buchan, D.; de Neve, S. Modeling soil moisture effects on net nitrogen mineralization in loamy wetland soils. Wetlands 2008, 28, 724–734. [Google Scholar] [CrossRef]
- Zak, D.R.; Grigal, D.F. Nitrogen mineralization, nitrification and denitrification in upland and wetland ecosystems. Oecologia 1991, 88, 189–196. [Google Scholar] [CrossRef]
- Cartaxana, P.; Caçador, I.; Vale, C.; Falcão, M.; Catarino, F. Seasonal variation of inorganic nitrogen and net mineralization in a salt marsh ecosystem. Mangroves Salt Marshes 1999, 3, 127–134. [Google Scholar] [CrossRef]
- Uri, V.; Lõhmus, K.; Tullus, H. Annual net nitrogen mineralization in a grey alder (Alnus incana (L.) Moench) plantation on abandoned agricultural land. For. Ecol. Manag. 2003, 184, 167–176. [Google Scholar] [CrossRef]
- Hefting, M.; Clément, J.C.; Dowrick, D.; Cosandey, A.C.; Bernal, S.; Cimpian, C.; Tatur, A.; Burt, T.P.; Pinay, G. Water table elevation controls on soil nitrogen cycling in riparian wetlands along a European climatic gradient. Biogeochemistry 2004, 67, 113–134. [Google Scholar] [CrossRef]
- Western, A.W.; Grayson, R.B.; Bloschl, G. Scaling of soil moisture: A hydrologic perspective. Annu. Rev. Earth Pl. Sc. 2002, 30, 149–180. [Google Scholar] [CrossRef]
- Porporato, A.; D’Odorico, P.; Laio, F.; Rodriguez-Iturbe, I. Hydrologic controls on soil carbon and nitrogen cycles. I. Modeling scheme. Adv. Water Resour. 2003, 26, 45–58. [Google Scholar] [CrossRef]
- Robinson, D.A.; Campbell, C.S.; Hopmans, J.W.; Hornbuckle, B.K.; Jones, S.B.; Knight, R.; Ogden, F.; Selker, J.; Wendroth, O. Soil moisture measurement for ecological and hydrological watershed-scale observatories: A review. Vadose Zone J. 2008, 7, 358–389. [Google Scholar] [CrossRef]
- Updegraff, K.; Pastor, J.; Bridgham, S.D.; Johnston, C.A. Environmental and substrate controls over carbon and nitrogen mineralization in northern wetlands. Ecol. Appl. 1995, 5, 151–163. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G.; Han, X.; Parsons, W.F.J.; Zhu, W. On the nature of environmental gradients: Temporal and spatial variability of soils and vegetation in the New Jersey pinelands. J. Ecol. 1997, 85, 785–798. [Google Scholar] [CrossRef]
- Yoon, T.K.; Noh, N.J.; Han, S.; Kwak, H.; Lee, W.-K.; Son, Y. Small-scale spatial variability of soil properties in a Korean swamp. Landsc. Ecol. Eng. 2015, 11, 723–734. [Google Scholar] [CrossRef]
- Khanna, P.K.; Raison, R.J. In situ core methods for estimating soil mineral-N fluxes: Re-evaluation based on 25 years of application and experience. Soil Biol. Biochem. 2013, 64, 203–210. [Google Scholar] [CrossRef]
- Friedel, J.K.; Herrmann, A.; Kleber, M. Ion exchange resin–soil mixtures as a tool in net nitrogen mineralisation studies. Soil Biol. Biochem. 2000, 32, 1529–1536. [Google Scholar] [CrossRef]
- Johnson, D.W.; Verburg, P.S.J.; Arnone, J.A. Soil extraction, ion exchange resin, and ion exchange membrane measures of soil mineral nitrogen during incubation of a tallgrass prairie soil. Soil Sci. Soc. Am. J. 2005, 69, 260–265. [Google Scholar] [CrossRef]
- Vitousek, P. Nutrient cycling and nutrient use efficiency. Am. Nat. 1982, 119, 553–572. [Google Scholar] [CrossRef]
- Gower, S.T.; Son, Y. Differences in soil and leaf litterfall nitrogen dynamics for five forest plantations. Soil Sci. Soc. Am. J. 1992, 56, 1959–1966. [Google Scholar] [CrossRef]
- Aerts, R. The advantages of being evergreen. Trends Ecol. Evol. 1995, 10, 402–407. [Google Scholar] [CrossRef]
- Reich, P.B.; Grigal, D.F.; Aber, J.D.; Gower, S.T. Nitrogen mineralization and productivity in 50 hardwood and conifer stands on diverse soils. Ecology 1997, 78, 335–347. [Google Scholar] [CrossRef]
- Mueller, K.E.; Hobbie, S.E.; Oleksyn, J.; Reich, P.B.; Eissenstat, D.M. Do evergreen and deciduous trees have different effects on net N mineralization in soil? Ecology 2012, 93, 1463–1472. [Google Scholar] [CrossRef] [PubMed]
- Binkley, D.; Giardina, C. Why do tree species affect soils? The warp and woof of tree-soil interactions. Biogeochemistry 1998, 42, 89–106. [Google Scholar] [CrossRef]
- Fujita, H.; Fujimura, Y. Distribution pattern and regeneration of swamp forest species with respect to site conditions. In Ecology of Riparian Forests in Japan; Sakio, H., Tamura, T., Eds.; Springer: Tokyo, Japan, 2008; pp. 225–236. [Google Scholar]
- Eom, H.J.; Chang, K.S.; Kim, H.; Chang, C.-S. Notes on a new overlooked taxon of Alnus (Betulaceae) in Korea. For. Sci. Technol. 2011, 7, 42–46. [Google Scholar]
- Whang, B.-C.; Lee, M.-W. Landscape ecology planning principles in korean Feng-Shui, Bi-Bo woodlands and ponds. Landsc. Ecol. Eng. 2006, 2, 147–162. [Google Scholar] [CrossRef]
- Yoon, T.K.; Noh, N.J.; Kim, R.-H.; Seo, K.W.; Lee, S.K.; Yi, K.; Lee, I.K.; Lim, J.-H.; Son, Y. Mass dynamics of coarse woody debris in an old-growth deciduous forest of Gwangneung, Korea. For. Sci. Technol. 2011, 7, 145–150. [Google Scholar] [CrossRef]
- Noh, N.J.; Son, Y.; Lee, S.K.; Yoon, T.K.; Seo, K.W.; Kim, C.; Lee, W.-K.; Bae, S.W.; Hwang, J. Influence of stand density on soil CO2 efflux for a Pinus densiflora forest in Korea. J. Plant Res. 2010, 123, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Noh, N.J.; Son, Y.; Jo, W.; Yi, K.; Park, C.W.; Han, S. Preliminary study on estimating fine root growth in a natural Pinus densiflora forest using a minirhizotron technique. For. Sci. Technol. 2012, 8, 47–50. [Google Scholar]
- Noh, N.J.; Kim, C.; Bae, S.W.; Lee, W.K.; Yoon, T.K.; Muraoka, H.; Son, Y. Carbon and nitrogen dynamics in a Pinus densiflora forest with low and high stand densities. J. Plant Ecol. 2013, 6, 368–379. [Google Scholar] [CrossRef]
- Yoon, T.K.; Noh, N.J.; Han, S.; Lee, J.; Son, Y. Soil moisture effects on leaf litter decomposition and soil carbon dioxide efflux in wetland and upland forests. Soil Sci. Soc. Am. J. 2014, 78, 1804–1816. [Google Scholar] [CrossRef]
- Vicca, S.; Luyssaert, S.; Peñuelas, J.; Campioli, M.; Chapin, F.S.; Ciais, P.; Heinemeyer, A.; Högberg, P.; Kutsch, W.L.; Law, B.E.; et al. Fertile forests produce biomass more efficiently. Ecol. Lett. 2012, 15, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Eno, C.F. Nitrate production in the field by incubating the soil in polyethylene bags. Soil Sci. Soc. Am. J. 1960, 24, 277–279. [Google Scholar] [CrossRef]
- Son, Y.; Lee, I.K. Soil nitrogen mineralization in adjacent stands of larch, pine and oak in central Korea. Ann. For. Sci. 1997, 54, 1–8. [Google Scholar] [CrossRef]
- Johansson, T. Biomass equations for determining fractions of common and grey alders growing on abandoned farmland and some practical implications. Biomass Bioenerg. 2000, 18, 147–159. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011; Available online: http://www.r-project.org (accessed on 15 June 2015).
- Craine, J.M.; Morrow, C.; Fierer, N. Microbial nitrogen limitation increases decomposition. Ecology 2007, 88, 2105–2113. [Google Scholar] [CrossRef] [PubMed]
- Shi, W. Agricultural and ecological significance of soil enzymes: Soil carbon sequestration and nutrient cycling. In Soil Enzymology; Shukla, G., Varma, A., Eds.; Springer: Berlin-Heidelberg, Germany, 2011; Volume 22, pp. 43–60. [Google Scholar]
- Regina, K.; Nykänen, H.; Maljanen, M.; Silvola, J.; Martikainen, P.J. Emissions of N2O and NO and net nitrogen mineralization in a boreal forested peatland treated with different nitrogen compounds. Can. J. For. Res. 1998, 28, 132–140. [Google Scholar] [CrossRef]
- Höfferle, Š.; Nicol, G.W.; Pal, L.; Hacin, J.; Prosser, J.I.; Mandić-Mulec, I. Ammonium supply rate influences archaeal and bacterial ammonia oxidizers in a wetland soil vertical profile. FEMS Microbiol. Ecol. 2010, 74, 302–315. [Google Scholar]
- Carpenter-Boggs, L.; Pikul, J.L.; Vigil, M.F.; Riedell, W.E. Soil nitrogen mineralization influenced by crop rotation and nitrogen fertilization. Soil Sci. Soc. Am. J. 2000, 64, 2038–2045. [Google Scholar] [CrossRef]
- Fisk, M.; Fahey, T. Microbial biomass and nitrogen cycling responses to fertilization and litter removal in young northern hardwood forests. Biogeochemistry 2001, 53, 201–223. [Google Scholar] [CrossRef]
- Bowden, R.D.; Davidson, E.; Savage, K.; Arabia, C.; Steudler, P. Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard forest. For. Ecol. Manag. 2004, 196, 43–56. [Google Scholar] [CrossRef]
- Treseder, K.K. Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies. Ecol. Lett. 2008, 11, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Matson, P.; Lohse, K.A.; Hall, S.J. The globalization of nitrogen deposition: Consequences for terrestrial ecosystems. AMBIO 2002, 31, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Shin, A.Y.; Sung, M.Y.; Choi, J.S.; On, J.S.; Roh, S.A.; Ahn, J.Y.; Han, J.S.; Lee, G.W. A study on the acid deposition of inorganic ions and flux in Seoul, Gangwha, Icheon, 2008. J. Korean Soc. Urban Environ. 2012, 12, 14–22. (In Korean) [Google Scholar]
- Jacks, G.; Joelsson, A.; Fleischer, S. Nitrogen retention in forest wetlands. AMBIO 1994, 23, 358–362. [Google Scholar]
- Son, Y. Non-symbiotic nitrogen fixation in forest ecosystems. Ecol. Res. 2001, 16, 183–196. [Google Scholar] [CrossRef]
- Vitousek, P.; Cassman, K.; Cleveland, C.; Crews, T.; Field, C.; Grimm, N.; Howarth, R.; Marino, R.; Martinelli, L.; Rastetter, E.; et al. Towards an ecological understanding of biological nitrogen fixation. Biogeochemistry 2002, 57–58, 1–45. [Google Scholar] [CrossRef]
- Han, J.S.; Hong, Y.D.; Ahn, J.Y.; Chung, I.R.; Shin, A.Y.; Lim, B.K.; Noh, S.A.; Son, J.S.; Park, J.H. Acid Deposition Monitoring and Impact Assessment (V): Centered on Forest Ecosystem; National Institute of Environmental Research: Incheon, Korea, 2008. [Google Scholar]
- Lee, Y.; Son, Y. Diurnal and seasonal patterns of nitrogen fixation in an Alnus hirsuta plantation of central Korea. J. Plant Biol. 2005, 48, 332–337. [Google Scholar] [CrossRef]
- Kim, D.K. Comparisons of Physico-Chemical Properties of Soil between the Alnus japonica Wetlands and Adjacent Slope. Master’s Thesis, Kongju National University, Gongju, Korea, Febuary 2014. [Google Scholar]
- Bedard-Haughn, A.; Matson, A.L.; Pennock, D.J. Land use effects on gross nitrogen mineralization, nitrification, and N2O emissions in ephemeral wetlands. Soil Biol. Biochem. 2006, 38, 3398–3406. [Google Scholar] [CrossRef]
- Durán, J.; Delgado-Baquerizo, M.; Rodríguez, A.; Covelo, F.; Gallardo, A. Ionic exchange membranes (IEMs): A good indicator of soil inorganic N production. Soil Biol. Biochem. 2013, 57, 964–968. [Google Scholar] [CrossRef]
- Qian, P.; Schoenau, J.J. Practical applications of ion exchange resins in agricultural and environmental soil research. Canad. J. Soil Sci. 2002, 82, 9–21. [Google Scholar] [CrossRef]
- Van Raij, B. Bioavailable tests: Alternatives to standard soil extractions. Commun. Soil Sci. Plant Anal. 1998, 29, 1553–1570. [Google Scholar] [CrossRef]
- Meason, D.F.; Idol, T.W. Nutrient sorption dynamics of resin membranes and resin bags in a tropical forest. Soil. Sci. Soc. Am. J. 2008, 72, 1806–1814. [Google Scholar] [CrossRef]
- Nadelhoffer, K.J.; Aber, J.D.; Melillo, J.M. Leaf-litter production and soil organic matter dynamics along a nitrogen-availability gradient in southern Wisconsin (U.S.A.). Can. J. For. Res. 1983, 13, 12–21. [Google Scholar] [CrossRef]
- Binkley, D.; Valentine, D. Fifty-year biogeochemical effects of green ash, white pine, and Nrway spruce in a replicated experiment. For. Ecol. Manag. 1991, 40, 13–25. [Google Scholar] [CrossRef]
- Kim, C.; Son, Y.; Lee, W.-K.; Jeong, J.; Noh, N.-J.; Kim, S.-R.; Yang, A.R.; Ju, N.-G. Influence of forest tending (Soopkakkugi) works on litterfall and nutrient inputs in a Pinus densiflora stand. For. Sci. Technol. 2012, 8, 83–88. [Google Scholar]
- Kang, J.-S.; Shibuya, M.; Shin, C.-S. The effect of forest-thinning works on tree growth and forest environment. For. Sci. Technol. 2014, 10, 33–39. [Google Scholar] [CrossRef]
- Franklin, J.F.; Spies, T.A.; van Pelt, R.; Carey, A.B.; Thornburgh, D.A.; Berg, D.R.; Lindenmayer, D.B.; Harmon, M.E.; Keeton, W.S.; Shaw, D.C.; et al. Disturbances and structural development of natural forest ecosystsems with silvicultural implications, using Douglas-fir forests as an example. For. Ecol. Manag. 2002, 1–3, 399–423. [Google Scholar] [CrossRef]
- Zak, D.R.; Groffman, P.M.; Pregitzer, K.S.; Christensen, S.; Tiedje, J.M. The vernal dam: Plant-microbe competition for nitrogen in northern hardwood forests. Ecology 1990, 71, 651–656. [Google Scholar] [CrossRef]
- Aulakh, M.S.; Singh, K.; Singh, B.; Doran, J.W. Kinetics of nitrification under upland and flooded soils of varying texture. Commun. Soil Sci. Plant Anal. 1996, 27, 2079–2089. [Google Scholar] [CrossRef]
- Yoo, J. Hydrological Properties and Nutrient Budgets in a Coniferous Forest Catchment in Gwangneung, Gyeonggi Province, Korea. Ph.D. Thesis, Korea University, Seoul, Korea, Febuary 2010. [Google Scholar]
- Fisher, J.; Acreman, M.C. Wetland nutrient removal: A review of the evidence. Hydrol. Earth Syst. Sci. 2004, 8, 673–685. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, T.K.; Noh, N.J.; Chung, H.; Yang, A.-R.; Son, Y. Soil Nitrogen Transformations and Availability in Upland Pine and Bottomland Alder Forests. Forests 2015, 6, 2941-2958. https://doi.org/10.3390/f6092941
Yoon TK, Noh NJ, Chung H, Yang A-R, Son Y. Soil Nitrogen Transformations and Availability in Upland Pine and Bottomland Alder Forests. Forests. 2015; 6(9):2941-2958. https://doi.org/10.3390/f6092941
Chicago/Turabian StyleYoon, Tae Kyung, Nam Jin Noh, Haegeun Chung, A-Ram Yang, and Yowhan Son. 2015. "Soil Nitrogen Transformations and Availability in Upland Pine and Bottomland Alder Forests" Forests 6, no. 9: 2941-2958. https://doi.org/10.3390/f6092941
APA StyleYoon, T. K., Noh, N. J., Chung, H., Yang, A. -R., & Son, Y. (2015). Soil Nitrogen Transformations and Availability in Upland Pine and Bottomland Alder Forests. Forests, 6(9), 2941-2958. https://doi.org/10.3390/f6092941