
Variability of Stand Structures and Development in Old-Growth Forests in the Pacific Northwest, USA


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sample Plots

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Identification | Mean Altitude (m) | Aspect (°) | Slope (%) | Physiography | Oldest Tree Age (Year) | Age Distribution | Fire | Windthrow |
|---|---|---|---|---|---|---|---|---|
| 1 | 701.0 | 218 | 47 | Bench/terrace | 132 | Single cohort | - | - |
| 2 | 1,066.8 | 170 | 65 | Sidehill/middle 1/3 | 132 | Single cohort | Yes | - |
| 3 | 701.0 | 256 | 30 | Sidehill/middle 1/3 | 155 | Single cohort | Yes | Small |
| 4 | 670.6 | 30 | 50 | Sidehill/upper1/3 | 120 | Single cohort | Yes | Small |
| 5 | 548.6 | 340 | 25 | Sidehill/lower 1/3 | 252 | Continuous DF-WH | Yes | Small |
| 6 | 487.7 | 80 | 20 | Sidehill/lower 1/3 | 240 | Continuous DF-WH | - | Small |
| 7 | 609.6 | 355 | 60 | Sidehill/middle | 294 | Continuous DF-WH | Yes | Large |
| 8 | 487.7 | 1 | 0 | Broad flat | 258 | Aggregate | - | Some |
| 9 | 1,127.8 | 199 | 9 | Bench/terrace | 482 | Continuous DF-WH | - | Some |
| 10 | 457.2 | 360 | 30 | Sidehill/lower 1/3 | 426 | Continuous | - | Some |
| 11 | 548.6 | 90 | 65 | Narrow ridgetop/peak | 490 | Aggregate | - | Small |
| 12 | 1,005.8 | 270 | 30 | Sidehill/lower 1/3 | 549 | Continuous/aggregate | - | - |
| 13 | 731.5 | 340 | 30 | Sidehill/middle 1/3 | 426 | Continuous/aggregate | - | Small |
| 14 | 1,097.3 | 1 | 1 | Broad flat | 999 | Aggregate | - | - |
| 15 | 792.5 | 30 | 50 | Sidehill/lower 1/3 | 998 | Continuous/aggregate | - | Large |
| 16 | 609.6 | 360 | 90 | Sidehill/lower 1/3 | 600 | Continuous/aggregate | Yes | Small |
2.3. Data Collection
2.4. Data Analysis
| Abbreviation forTable 3 and Table 4 | Structural Elements | Source of Information and Relevant References |
|---|---|---|
| # Tree species | Number of tree species | |
| R BA DF | Ratio of Douglas-fir basal area to total stand basal area | Shade intolerant Douglas-fir [37] |
| R BA ST | Ratio of shade tolerant tree basal to total stand basal area | Ratio of shade tolerant species [37] |
| D trees ≥100 cm | Number of trees ≥100 cm DBH ha−1 | 40 (ca. 100 cm) to 60 inches in diameter [36] |
| D DF ≥100 cm | Number of Douglas-fir ≥100 cm DBH ha−1 | Douglas-fir older than 200 years [23,36], Douglas-fir trees with DBH of 1 to 2 m [38] |
| D ST ≥50 cm | Number of shade tolerant trees ≥50 cm DBH ha−1 | Shade tolerant species ≥50 cm DBH |
| D snag ≥50 cm | Number of snags ≥50 cm DBH ha−1 | Density of snags >50 cm DBH and >15 m tall [10], ≥50 cm DBH [38] |
| D log ≥50 cm | Number of logs ≥50 cm diameter ha−1 | Density of logs >60 cm diameter [23] |
| DBH classes | Number of 10-cm DBH classes containing trees | Structural heterogeneity [39] |
| Max DBH | Maximum DBH class | Large tree size |
| Missing DBH ≥ 50 cm | Number of 10-cm DBH classes without trees between 50 cm & max DBH: “missing upper DBH classes” | Broken or continuous canopy of medium to large [24] |
| Missing DBH < 50 cm | Number of 10-cm DBH classes without trees <50 cm DBH: “missing lower DBH classes” | Understory absent or consisting of some seedlings [24] |
2.5. Stand Development Reconstruction
3. Results
3.1. Relationships among Structural Variables
| Variable (See Table 2) | # Tree Species | R BA DF | R BA ST | D Trees ≥100 cm | D DF ≥100 cm | D ST ≥50 cm | D Snag ≥50 cm | D Log ≥50 cm | DBH Classes | DF DBH Classes | Max DBH Class | Missing DBH ≥ 50 cm |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R BA DF | −0.291 | |||||||||||
| R BA ST | 0.203 | −0.900 *** | ||||||||||
| D trees ≥100 cm | −0.515 * | 0.546 * | −0.577 * | |||||||||
| D DF ≥100 cm | −0.415 | 0.715 ** | −0.821 *** | 0.867 *** | ||||||||
| D ST ≥50 cm | 0.060 | −0.681 ** | 0.762 ** | −0.227 | −0.487 | |||||||
| D snag ≥50 cm | −0.243 | 0.103 | −0.094 | 0.214 | 0.303 | 0.213 | ||||||
| D log ≥50 cm | 0.403 | −0.275 | 0.284 | −0.140 | −0.244 | 0.360 | −0.113 | |||||
| DBH classes | −0.097 | 0.373 | −0.298 | 0.126 | 0.331 | 0.095 | 0.296 | −0.146 | ||||
| DF DBH classes | −0.220 | 0.920 *** | −0.852 *** | 0.395 | 0.622 * | −0.760 ** | −0.018 | −0.144 | 0.243 | |||
| Max DBH class | −0.245 | 0.409 | −0.458 | 0.869 *** | 0.753 ** | −0.018 | 0.349 | −0.048 | 0.162 | 0.268 | ||
| Missing DBH ≥50 cm | −0.067 | 0.229 | −0.332 | 0.786 *** | 0.629 ** | 0.055 | 0.304 | 0.090 | 0.020 | 0.110 | 0.958 *** | |
| Missing DBH <50 cm | −0.756 ** | 0.468 | −0.313 | 0.603 * | 0.415 | −0.155 | 0.045 | −0.368 | −0.048 | 0.344 | 0.449 | 0.306 |
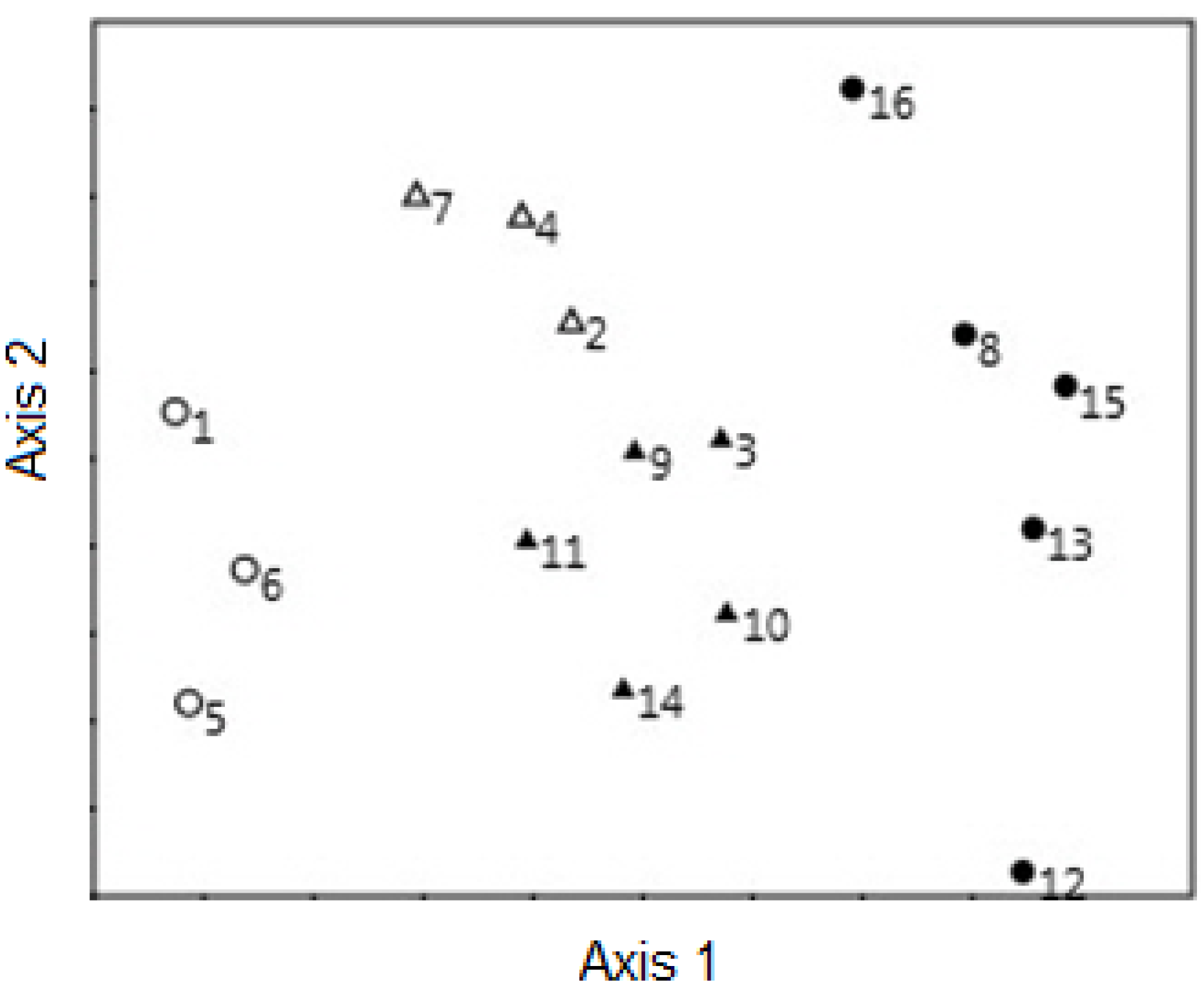
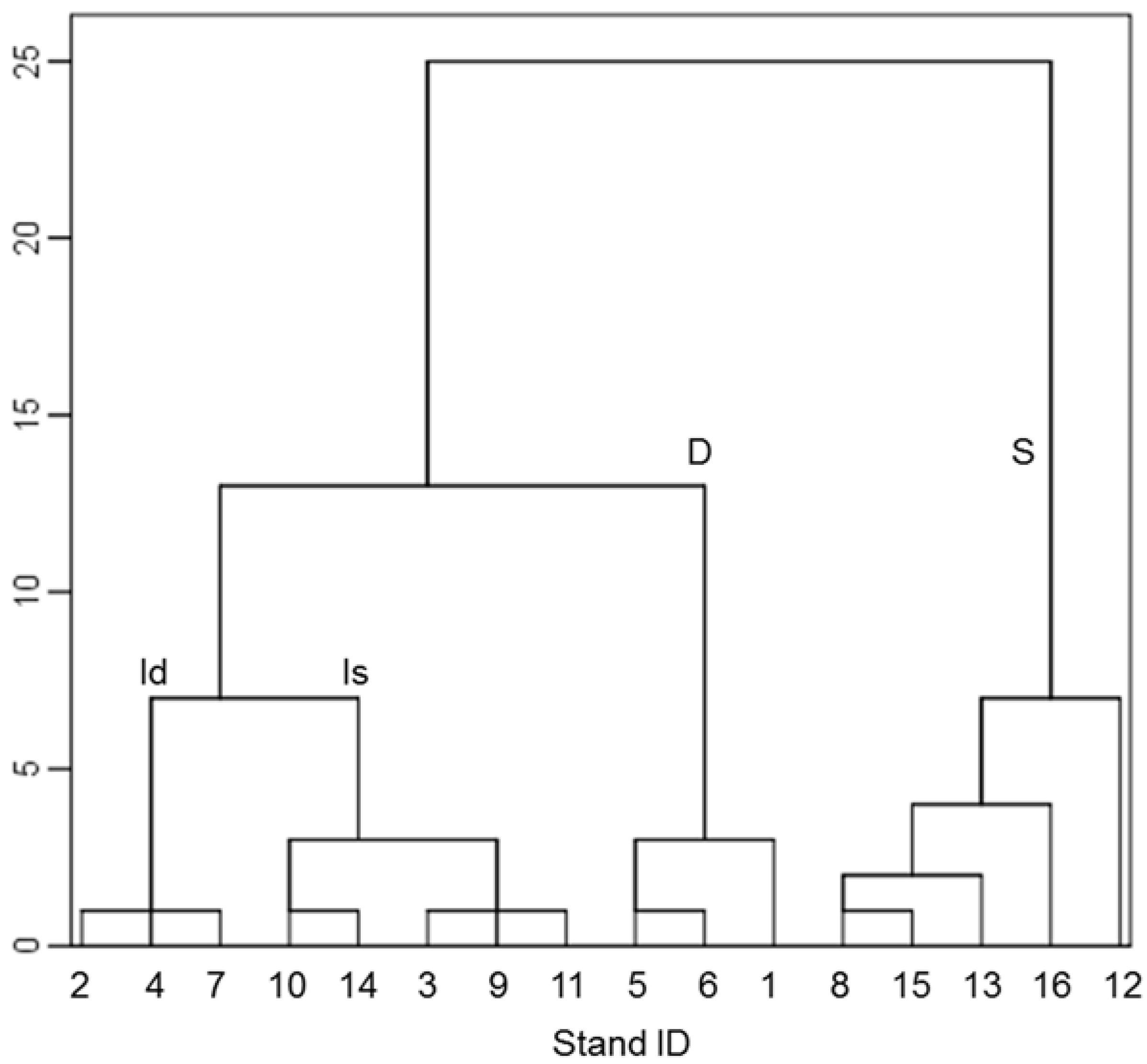
3.2. Non-Metric Multidimensional Scaling and Grouping of Stands


3.3. Structural Characteristics of Grouped Stands


3.4. Differences and Similarities in Structural Features of Grouped Stands
| Abbreviation (Table 2) | Group D | Group Id | Group Is | Group S | Mean |
|---|---|---|---|---|---|
| R BA DF | 91.0 (1.7) a | 57.9 (4.8) b | 57.0 (3.2) b | 2.8 (2.5) c | 46.6 (8.4) |
| R BA ST | 7.6 (2.9) a | 36.8 (3.6) b | 42.5 (4.1) b | 86.3 (4.3) c | 48.6 (7.5) |
| D trees ≥100 cm | 44 (4.4) a | 13 (7.6) b | 24 (7.7) ab | 8 (5.3) b | 21 (4.5) |
| D DF ≥100 cm | 44 (4.7) a | 13 (7.5) b | 18 (6.6) b | 0 b | 17 (4.6) |
| D ST ≥50 cm | 0 b | 17 (4.3) ab | 66 (9.3) a | 66 (14.4) a | 44 (8.8) |
| D snag ≥50 cm | 44 (23.2) ns | 17 (4.3) ns | 42 (7.8) ns | 39 (21.6) ns | 37 (8.0) |
| D log ≥50 cm | 0.0 b | 61 (8.8) a | 21 (6.6) b | 21 (6.6) b | 25 (5.9) |
| DBH classes | 8.7 (0.7) | 8.0 (0.6) | 9.2 (0.5) | 7.8 (0.6) | 8.4 (0.3) |
| DF DBH classes | 6.0 (1.0) a | 4.3 (0.3) ab | 2.6 (0.5) bc | 0.4 (0.2) c | 2.9 (0.6) |
| Max DBH class | 16 (2.1) ns | 11 (2.0) ns | 16 (2.7) ns | 10 (1.0) ns | 13 (1.2) |
| Missing DBH ≥50 cm | 5.3 (2.0) ns | 3.3 (2.4) ns | 6.2 (2.6) ns | 1.8 (0.7) ns | 4.1 (1.0) |
| Missing DBH <50 cm | 2.0 (0.0) a | 0.0 (0.0) b | 1.0 (0.5) ab | 0.2 (0.2) b | 0.8 (0.3) |
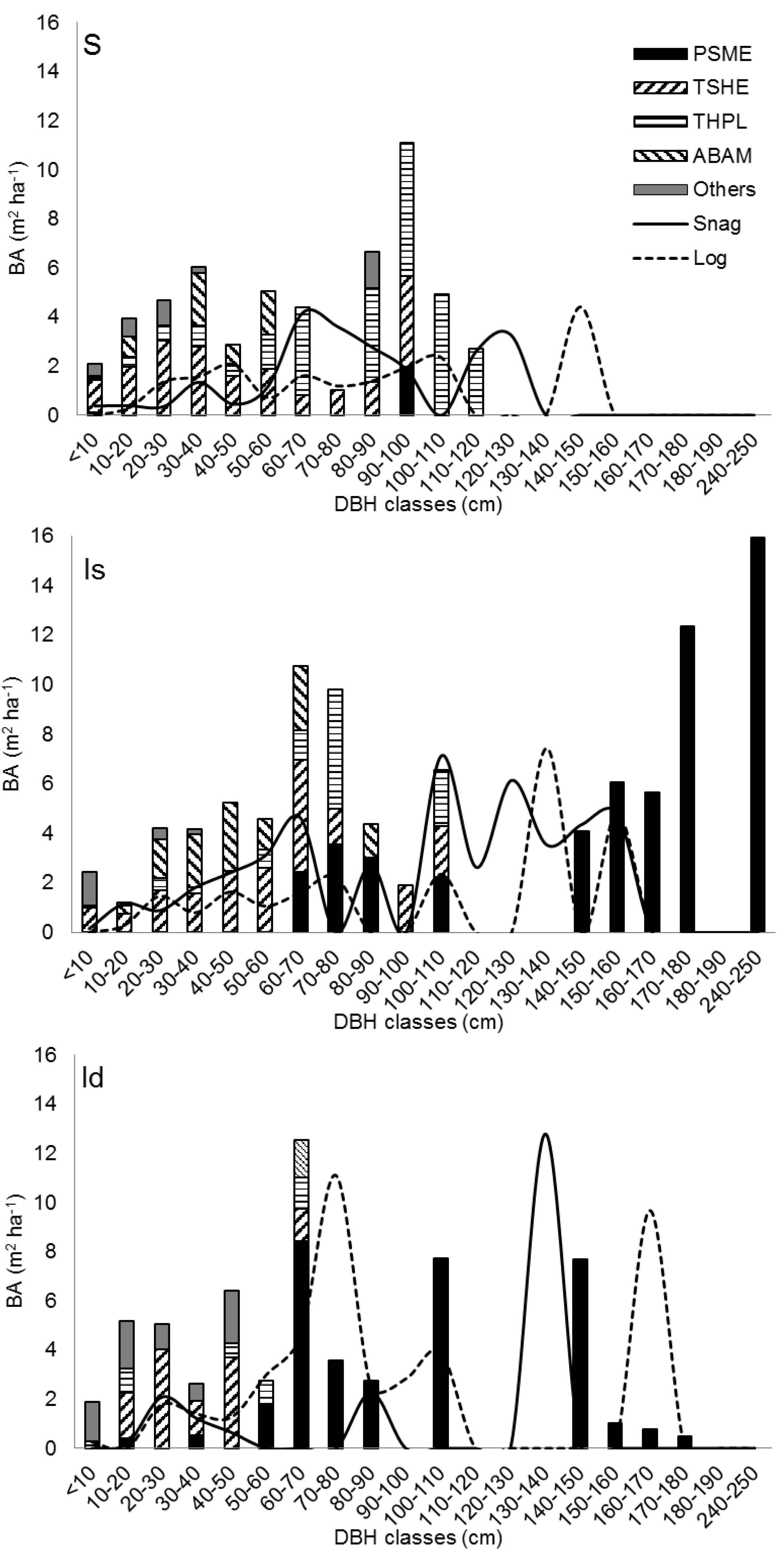
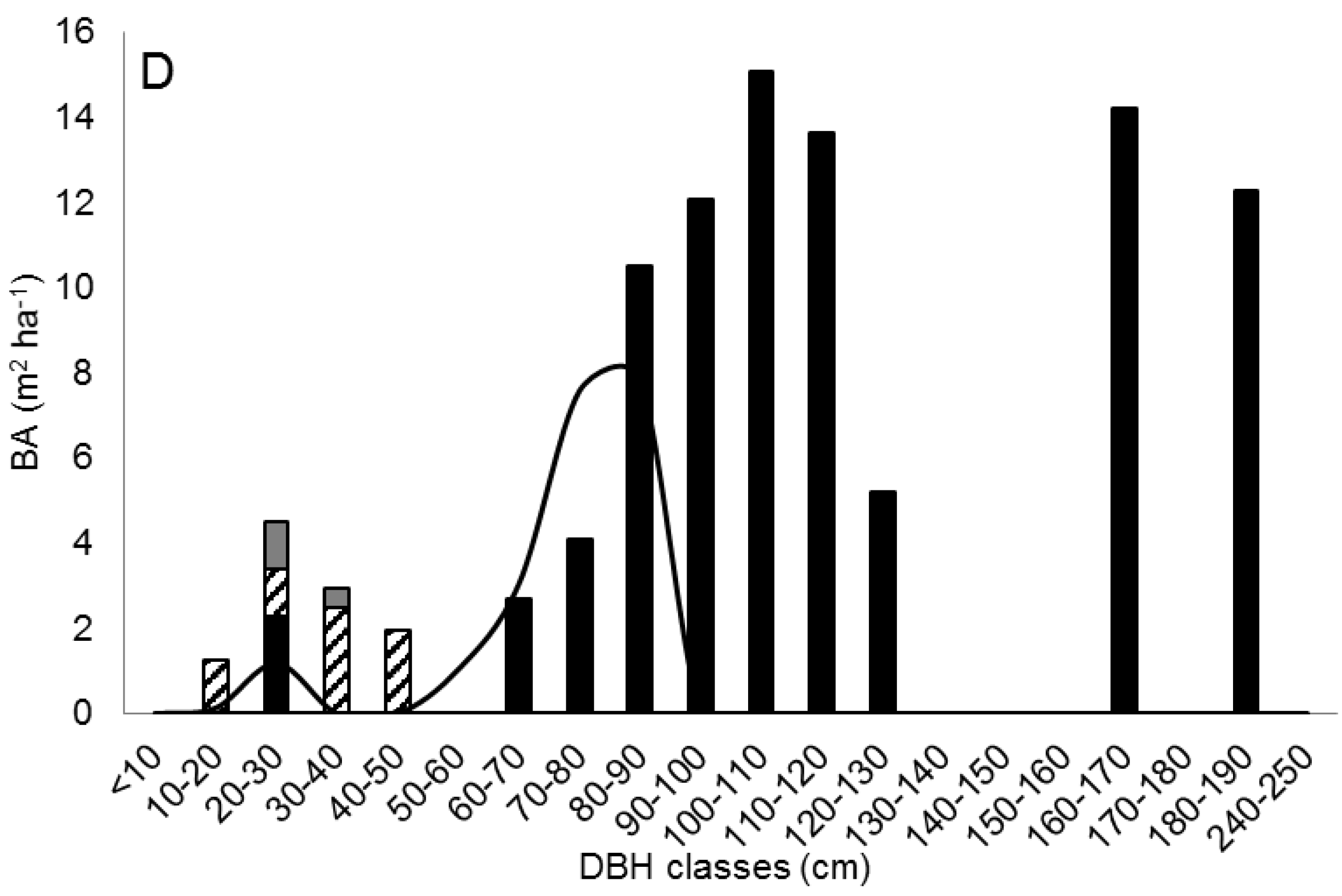
3.5. Reconstruction of a Sequential Change in Stand Structures
- In all plots with Douglas-firs, the largest western hemlocks and western redcedars are noticeably smaller than the largest Douglas-firs, suggesting that the three species were or had been growing together and/or the hemlocks and redcedars were younger.
- In S, the hemlocks and redcedars are small for the age of the stand but there are no larger trees of any species, suggesting they are either young or are/were suppressed—in either case by larger trees that are no longer alive. This indicates a sequence of “S occurring later than D, Id, and Is.”
- The order “D, Id, and Is” shows increasingly fewer numbers of Douglas-firs but increasingly greater diameters of the largest Douglas-firs and indicates a possible sequence in that order.
- The order “D, Id, and Is” shows increasingly fewer numbers of large Douglas-firs, but increasing numbers of snags and logs of large sizes, indicating a possible sequence of the large Douglas-firs dying.
- The order “D, Id, and Is” showed increasing gaps in the diameter distribution of large Douglas-firs, indicating a possible sequence with the overstory developing gaps.
- The near absence of large Douglas-firs in S, but the presence of snags and logs much larger than present shade tolerant species indicates a sequence of “S occurring later than D, Id, and Is.”
| Franklin et al. [5] | Oliver and Larson [4] | Sub-structure categories in this paper: |
|---|---|---|
| Vertical Diversification | Transition Old-growth | D |
| Horizontal Diversification | Transition Old-growth | Id & Is |
| Pioneer Cohort Loss | True Old-growth | S |
4. Discussion

5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Booth, D.E. Estimating prelogging old-growth in the Pacific Northwest. J. For. 1991, 89, 25–29. [Google Scholar]
- Forest Ecosystem Management Assessment Team (FEMAT). Forest Ecosystem Management: An Ecological, Economic, and Social Assessment. Draft Supplemental Environmental Impact Statement: On Management of Habitat for Late-Successional and Old-Growth Forest Related Species within the Range of the Northern Spotted Owl; FEMAT: Washington, DC, USA, 1993. [Google Scholar]
- Hedman, C.W.; van Lear, D.H.; Swank, W.T. In-stream large woody debris loading and riparian forest seral stage associations in the southern Appalachian Mountains. Can. J. For. Res. 1996, 26, 1218–1227. [Google Scholar] [CrossRef]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; John Wiley and Sons: New York, NY, USA, 1996. [Google Scholar]
- Franklin, J.F.; Spies, T.A.; Pelt, R.V.; Carey, A.B.; Thornburgh, D.A.; Berg, D.R.; Lindenmayer, D.B.; Harmong, M.E.; Keeton, W.S.; Shawh, D.C.; et al. Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. For. Ecol. Manag. 2002, 155, 399–423. [Google Scholar] [CrossRef]
- Carey, A.B.; Curtis, R.O. Conservation of biodiversity: A useful paradigm for forest ecosystem management. Wildl. Soc. Bull. 1996, 24, 610–620. [Google Scholar]
- Spies, T.A.; Duncan, S.L. Old Growth in a New World; Island Press: Washington, DC, USA, 2009. [Google Scholar]
- Spies, T.A.; Franklin, J.F. The structure of natural young, mature, and old-growth Douglas-fir forests in Oregon and Washington. In Wildlife and Vegetation of Unmanaged Douglas-fir Forests; Ruggiero, L.F., Aubry, K.B., Eds.; General Technical Report PNW-GTR-285; USDA Forest Service: Portland, OR, USA, 1991; pp. 91–110. [Google Scholar]
- Nystrom, M.N.; DeBell, D.S.; Oliver, C.D. Development of Young Growth Western Red Cedar Stands; USDA Forest Service Research Paper PNW-324; USDA Forest Service: Portland, OR, USA, 1984; p. 9. [Google Scholar]
- Zenner, E.K. Development of tree size distributions in Douglas-fir forests under differing disturbance regimes. Ecol. Appl. 2005, 15, 701–714. [Google Scholar] [CrossRef]
- Donato, D.C.; Campbell, J.L.; Franklin, J.F. Multiple successional pathways and precocity in forest development: Can some forests be born complex? J. Veg. Sci. 2012, 23, 576–584. [Google Scholar] [CrossRef]
- Wierman, C.A.; Oliver, C.D. Crown stratification by species in even-aged mixed stands of Douglas-fir/western hemlock. Can. J. For. Res. 1979, 9, 1–9. [Google Scholar] [CrossRef]
- Spies, T.A. Ecological concepts and diversity of old-growth forests. J. For. 2004, 102, 14–20. [Google Scholar]
- Tsuga heterophylla. Available online: http://www.fs.fed.us/database/feis/ (accessed on 1 August 2011).
- Minore, D. Thuja plicata Donn ex D. Don. In Silvics of North America, Volume 1, Conifers; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service Agriculture Handbook 654; USDA Forest Service: Washington, DC, USA, 1990; p. 675. [Google Scholar]
- Oliver, C.D. Forest development in North America following major disturbances. For. Ecol. Manag. 1981, 3, 153–168. [Google Scholar] [CrossRef]
- Tappeiner, J.C.; Huffman, D.; Marshall, D.; Spies, T.A.; Bailey, J.D. Density, ages, and growth rates in old-growth and young-growth forests in coastal Oregon. Can. J. For. Res. 1997, 27, 638–648. [Google Scholar] [CrossRef]
- Winter, L.E.; Brubaker, L.B.; Franklin, J.F.; Miller, E.A.; DeWitt, D.Q. Canopy disturbances over the five-century lifetime of an old-growth Douglas-fir stand in the Pacific Northwest. Can. J. For. Res. 2002, 32, 1057–1070. [Google Scholar] [CrossRef]
- Freund, J.A.; Franklin, J.F.; Lutz, J.A. Structure of early old-growth Douglas-fir forests in the Pacific Northwest. For. Ecol. Manag. 2015, 335, 11–25. [Google Scholar] [CrossRef]
- Franklin, J.F.; Cromack, K., Jr.; Denison, W.; McKee, A.; Maser, C.; Sedell, J.; Swanson, F.; Juday, G. Ecological Characteristics of Old-growth Douglas-fir Forests; General Technical Report PNW-GTR-118; USDA Forest Service: Portland, OR, USA, 1981. [Google Scholar]
- Brown, E.R. (Ed.) Management of Wildlife and Fish Habitats in Forests of Western Oregon and Washington. Part I-Chapter Narratives; USDA Forest Service Publication No. R6-F&WL-192-1985; USDA Forest Service: Portland, OR, USA, 1985.
- Spies, T.A.; Franklin, J.F.; Thomas, T.B. Coarse woody debris in Douglas-fir forests of western Oregon and Washington. Ecology 1988, 69, 1689–1702. [Google Scholar] [CrossRef]
- Hansen, A.J.; Spies, T.A.; Swanson, F.J.; Omann, J.L. Conserving biodiversity in managed forests. Bioscience 1991, 41, 382–292. [Google Scholar] [CrossRef]
- O’Hara, K.L.; Latham, P.A.; Hessburg, P.; Smith, B.G. A structural classification for Inland Northwest forest vegetation. West. J. Appl. For. 1996, 11, 97–102. [Google Scholar]
- Burrascano, S.; Keeton, W.S.; Sabatini, F.M.; Blasi, C. Commonality and variability in the structural attributes of moist temperate old-growth forests: A global review. For. Ecol. Manag. 2013, 291, 458–479. [Google Scholar] [CrossRef]
- Franklin, J.F.; Dyrness, C.T. Natural Vegetation of Oregon and Washington; Oregon State University Press: Corvalis, OR, USA, 1988. [Google Scholar]
- Cispus. Available online: http://www.reo.gov/ama/locations/cispus.htm (accessed on 1 August 2011).
- Randle 1 E, Washington, NCDC 1981–2010 Monthly Normals. Available online: http://www.wrcc.dri.edu/cgi-bin/cliMAIN.pl?wa6909 (accessed on 9 August 2015).
- Packwood, Washington, NCDC 1981–2010 Monthly Normals. Available online: http://www.wrcc.dri.edu/cgi-bin/cliMAIN.pl?wa6262 (accessed on 9 August 2015).
- Soil Survey of Lewis County Area, Washington. Available online: http://www.nrcs.usda.gov/Internet/FSE_MANUSCRIPTS/washington/WA641/0/wa641_text.pdf (accessed on 14 July 2015).
- A Soil Survey of Skamania County Area, Washington. Available online: http://www.nrcs.usda.gov/Internet/FSE_MANUSCRIPTS/washington/WA659/0/wa659_text.pdf (accessed on 14 July 2015).
- Burns, R.M.; Honkala, B.H. Silvics of North America, Volume 1, Conifer; USDA Forest Service Agriculture Handbook 654; USDA Forest Service: Washington, DC, USA, 1990. [Google Scholar]
- Jiang, H.; Strittholt, J.R.; Frost, P.A.; Slosser, N.C. The classification of late seral forests in the Pacific Northwest, USA using Landsat ETM+ imagery. Remote Sens. Environ. 2004, 91, 320–331. [Google Scholar] [CrossRef]
- USDA Forest Service. Current Vegetation Survey, Version 1.5; Pacific Northwest Region, USDA Forest Service: Portland, OR, USA, 1995. [Google Scholar]
- Smith, D.; Larson, B.C.; Kelty, M.J.; Ashton, P.M.S. The Practice of Silviculture: Applied Forest Ecology, 9th ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1997. [Google Scholar]
- Society of American Foresters (SAF). Scheduling the Harvest of Old-growth; Society of American Foresters: Bethesda, MD, USA, 1984. [Google Scholar]
- North, M.; Chen, J.; Oakley, B.; Song, B.; Rudnicki, M.; Gray, A.; Innes, J. Forest stand structure and pattern of old-growth western hemlock/Douglas-fir and mixed-conifer forests. For. Sci. 2004, 50, 299–311. [Google Scholar]
- Spies, T.A.; Franklin, J.F. The diversity and maintenance of old-growth forests. In Biodiversity in Managed Landscapes: Theory and Practice; Szaro, R.C., Johnson, D.W., Eds.; Oxford University Press: New York, NY, USA, 1996; pp. 296–314. [Google Scholar]
- Franklin, J.F.; van Pelt, R. Spatial aspects of complexity in old-growth forests. J. For. 2004, 102, 22–28. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson Prentice-Hall: Upper Saddle River, NJ, USA, 2010. [Google Scholar]
- Kruskal, J.B. Nonmetric multidimensional scaling: A numerical method. Psychometrika 1964, 29, 115–129. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, p. 9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 11 February 2015).
- Lutz, H. Vegetation of Heart’s Content, a virgin forest in northwestern Pennsylvania. Ecology 1930, 11, 1–29. [Google Scholar] [CrossRef]
- Henry, J.D.; Swan, J.M.A. Reconstructing forest history from live and dead plant material—An approach to the study of forest succession in southwest New Hampshire. Ecology 1974, 55, 772–783. [Google Scholar] [CrossRef]
- Oliver, C.D.; Stephens, E.P. Reconstruction of a mixed species forest in central New England. Ecology 1977, 58, 562–572. [Google Scholar] [CrossRef]
- Huff, M.H. Forest age structure and development following wildfires in the western Olympic Mountains, Washington. Ecol. Appl. 1995, 5, 471–483. [Google Scholar] [CrossRef]
- Swetnam, T.W. Fire history and climate change in Giant Sequoia groves. Science 1993, 262, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D.; Mckane, R.B.; Tingey, D.T.; Beedlow, P.A. Vertical gradients in photosynthetic light response within an old-growth Douglas-fir and western hemlock canopy. Tree Physiol. 2000, 20, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.J.; Garman, S.L.; Weigand, J.F.; Urban, D.L.; McComb, W.C.; Raphael, M.G. Alternative silvicultural regimes in the Pacific Northwest: Simulations of ecological and economic effects. Ecol. Appl. 1995, 5, 535–554. [Google Scholar] [CrossRef]
- Coates, K.D. Conifer seedling response to northern temperate forest gaps. For. Ecol. Manag. 2000, 127, 249–269. [Google Scholar] [CrossRef]
- Kohyama, T. Size-structured tree populations in gap-dynamic forest—The forest architecture hypothesis for the stable coexistence of species. J. Ecol. 1993, 81, 131–143. [Google Scholar] [CrossRef]
- Knowles, P.; Grant, M.C. Age and size structure analyses of Engelmann spruce, ponderosa pine, lodgepole pine, and limber pine in Colorado. Ecology 1983, 64, 1–9. [Google Scholar] [CrossRef]
- Mailly, D.; Kimmins, J.P. Growth of Pseudotsuga menziesii and Tsuga heterophylla seedlings along a light gradient: Resource allocation and morphological acclimation. Can. J. For. Res. 1997, 75, 1424–1435. [Google Scholar]
- Pojar, J.; MacKinnon, A.; Alaback, P.B. Plants of the Pacific Northwest Coast: Washington, Oregon, British Columbia & Alaska; Lone Pine Publishing: Vancouver, BC, Canada, 1994. [Google Scholar]
- Jang, W.; Park, P.S. Stand structure and maintenance of Picea jezoensis in a northern temperate forest, South Korea. J. Plant Biol. 2010, 53, 180–189. [Google Scholar] [CrossRef]
- Nilsson, S.G.; Niklasson, M.; Hedin, J.; Aronsson, G.; Gutowski, J.M.; Linder, P.; Ljungberg, H.; Mikusiński, G.; Ranius, T. Erratum to “Densities of large living and dead trees in old-growth temperate and boreal forests”. For. Ecol. Manag. 2003, 178, 355–370. [Google Scholar] [CrossRef]
- Cline, S.P.; Berg, A.B.; Wight, H.M. Snag characteristics and dynamics in Douglas-fir forests, western Oregon. J. Wildl. Manag. 1980, 44, 773–786. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, P.S.; Oliver, C.D. Variability of Stand Structures and Development in Old-Growth Forests in the Pacific Northwest, USA. Forests 2015, 6, 3177-3196. https://doi.org/10.3390/f6093177
Park PS, Oliver CD. Variability of Stand Structures and Development in Old-Growth Forests in the Pacific Northwest, USA. Forests. 2015; 6(9):3177-3196. https://doi.org/10.3390/f6093177
Chicago/Turabian StylePark, Pil Sun, and Chadwick D. Oliver. 2015. "Variability of Stand Structures and Development in Old-Growth Forests in the Pacific Northwest, USA" Forests 6, no. 9: 3177-3196. https://doi.org/10.3390/f6093177
APA StylePark, P. S., & Oliver, C. D. (2015). Variability of Stand Structures and Development in Old-Growth Forests in the Pacific Northwest, USA. Forests, 6(9), 3177-3196. https://doi.org/10.3390/f6093177