
Effects of Hurricane-Felled Tree Trunks on Soil Carbon, Nitrogen, Microbial Biomass, and Root Length in a Wet Tropical Forest



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling Design
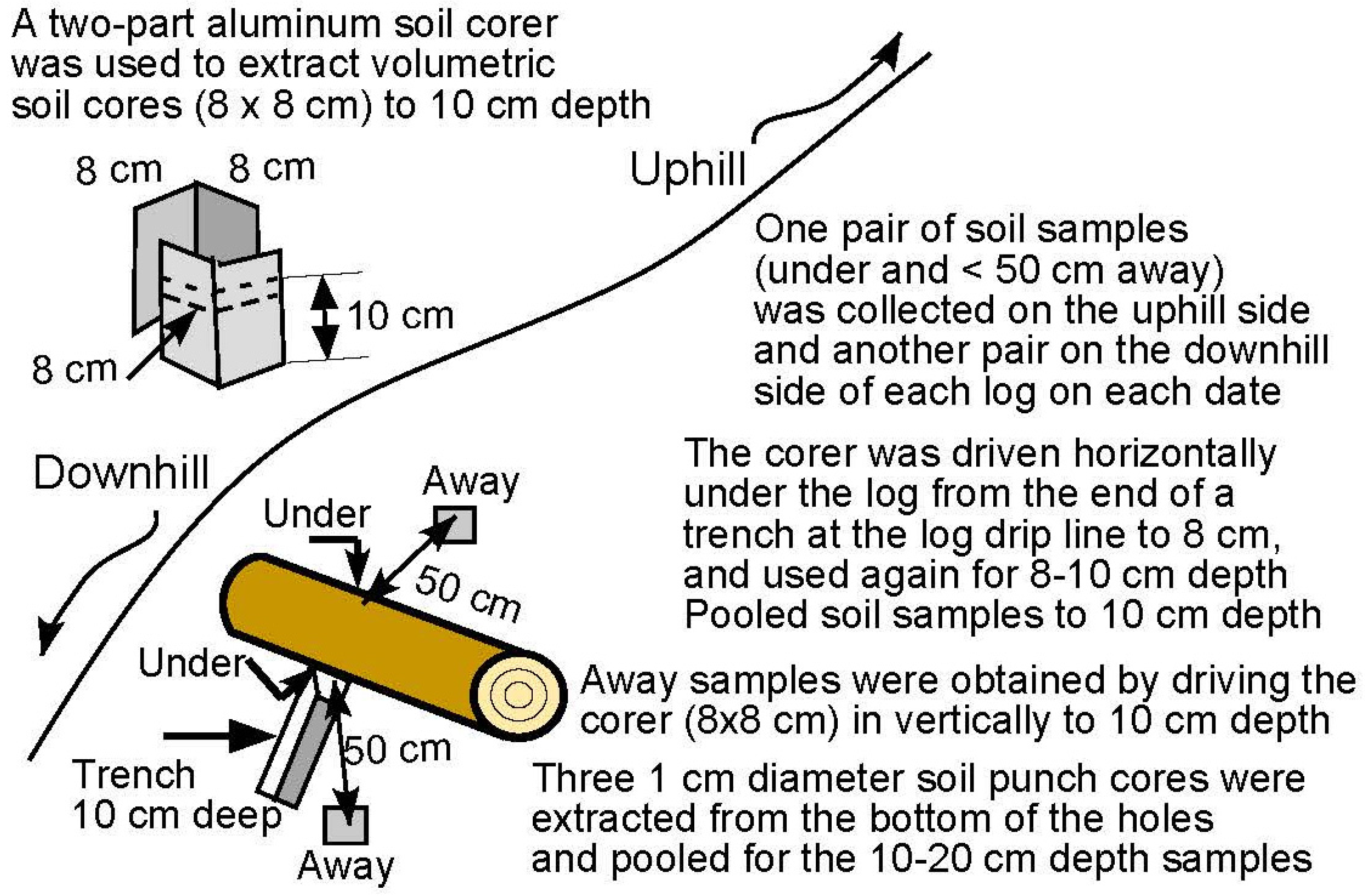
2.3. Soil Sampling and Analysis
2.3.1. Soil Sampling
2.3.2. Soil Nitrogen and Carbon Content
2.3.3. Soil Biota
2.4. Statistical Analyses
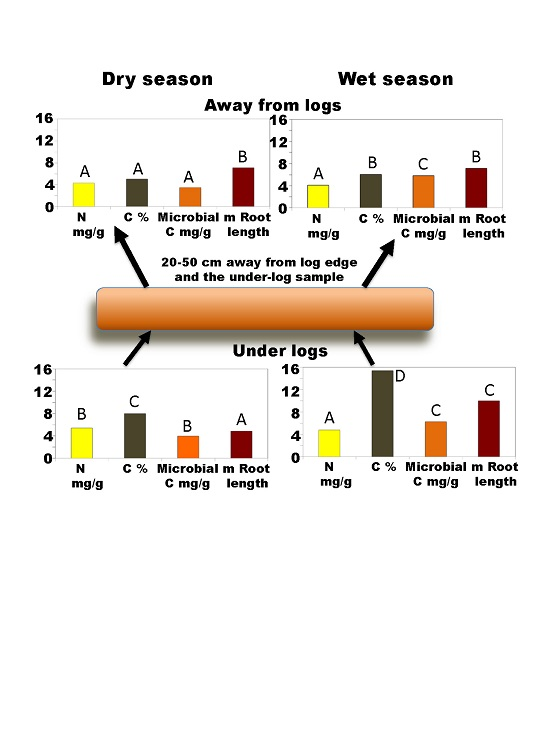
3. Results
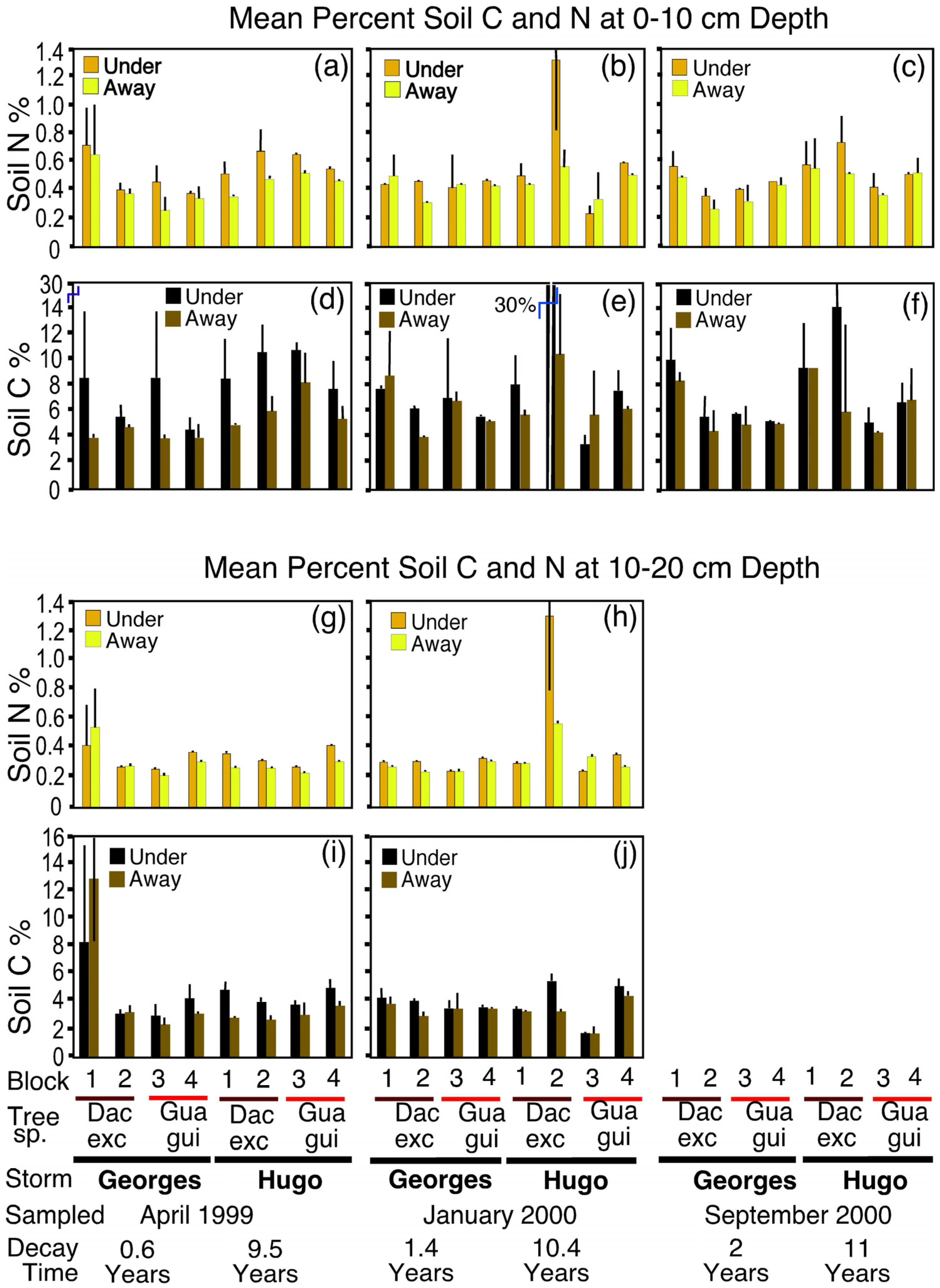
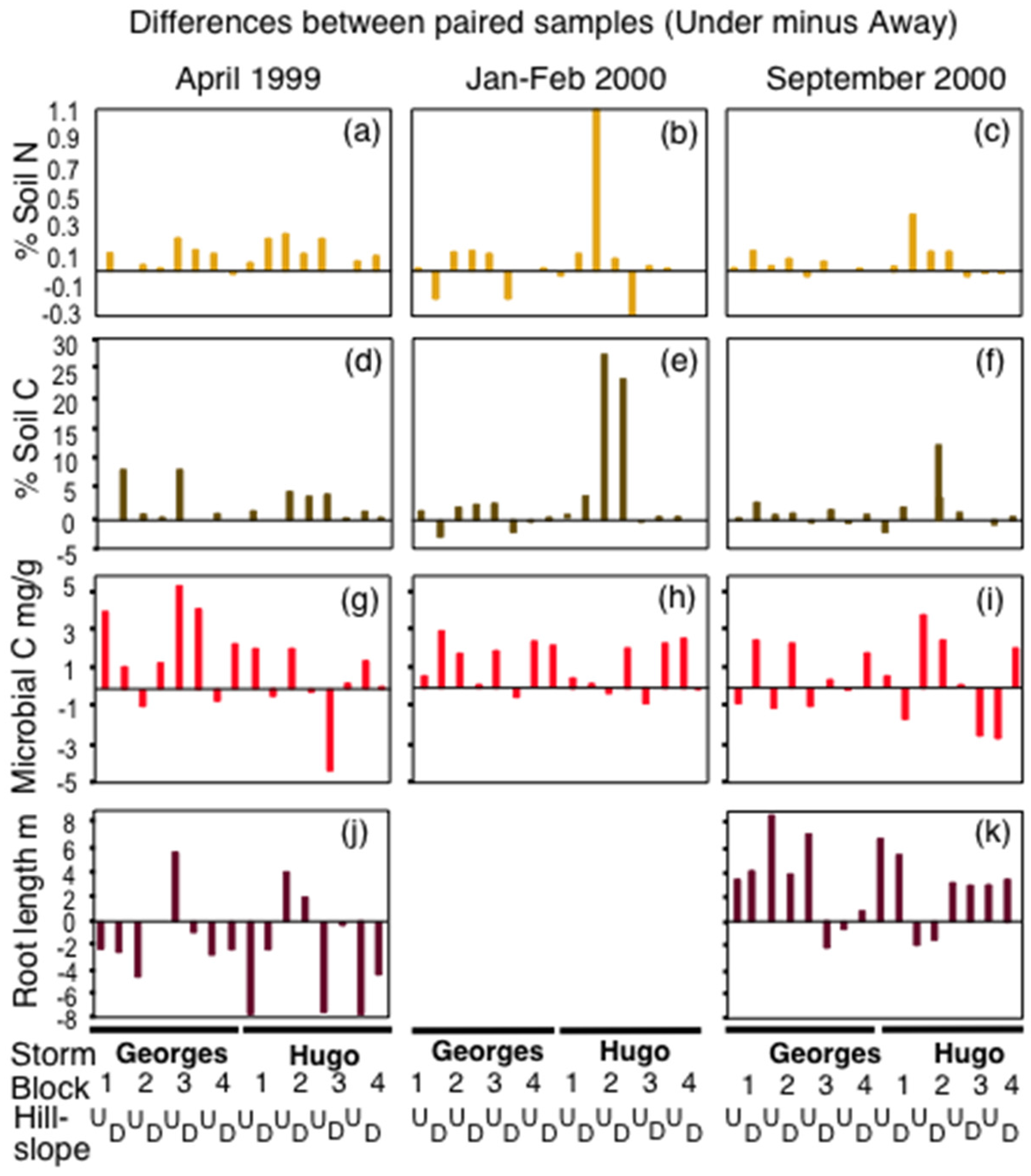
3.1. Soil Nitrogen and Carbon
3.2. Soil Biota
3.2.1. Soil Microbial Biomass C
3.2.2. Roots
3.3. Soil Moisture
3.4. Correlations among Variables
4. Discussion
4.1. Do Tree Roots Proliferate in Resource Hotspots?
4.2. Effects on Soil Nitrogen and Carbon
4.3. Effects on Biota
4.3.1. Root Responses
4.3.2. Soil Microbial Biomass Responses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bantle, A.; Borken, W.; Ellerbrock, R.H.; Schulze, E.D.; Weisser, W.W.; Matzner, E. Quantity and quality of dissolved organic carbon released from coarse woody debris of different tree species in the early phase of decomposition. For. Ecol. Manag. 2014, 329, 287–294. [Google Scholar] [CrossRef]
- Bantle, A.; Borken, W.; Matzner, E. Dissolved nitrogen release from coarse woody debris of different tree species in the early phase of decomposition. For. Ecol. Manag. 2014, 334, 277–283. [Google Scholar] [CrossRef]
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 1986, 15, 133–302. [Google Scholar]
- Edmonds, R.L.; Eglitis, A. The role of Douglas-fir beetle and wood borers in the decomposition of and nutrient release from Douglas-fir logs. Can. J. For. Res. 1989, 19, 853–859. [Google Scholar] [CrossRef]
- Schowalter, T.D. Heterogeneity and nutrient dynamics of oak (Quercus) logs during the first 2 years of decomposition. Can. J. For. Res. 1992, 22, 161–166. [Google Scholar] [CrossRef]
- Stevens, V. The Ecological Role of Coarse Woody Debris: An Overview of the Ecological Importance CWD in BC Forests; Work Paper 30; British Columbia Ministry of Forests: Victoria, BC, Canada, 1997.
- Ricker, M.C.; Graeme Lockaby, B.; Blosser, G.D.; Conner, W.H. Rapid wood decay and nutrient mineraliztion in an old-growth bottomland hardwood forest. Biogeochemistry 2016, 127, 323–338. [Google Scholar] [CrossRef]
- Johnson, D.W.; Woodward, C.; Meadows, M.W. A three-dimensional view of nutrient hotspots in a Sierra Nevada forest soil. Soil Sci. Soc. Am. J. 2014, 78, S225–S236. [Google Scholar] [CrossRef]
- Lodge, D.J.; McDowell, W.H.; Macy, J.; Ward, S.K.; Leisso, R.; Claudio Campos, K.; Kuhnert, K. Distribution and role of mat-forming saprobic basidiomycetes in a tropical forest. In Ecology of Saprobic Basidiomycetes; Boddy, L., Frankland, J.C., Eds.; Academic Press, Elsevier Ltd.: Amsterdam, The Netherlands, 2008; pp. 195–208. [Google Scholar]
- Stewart, C.G. A test of nutrient limitation in two tropical montane forests using root ingrowth cores. Biotropica 2000, 32, 369–373. [Google Scholar] [CrossRef]
- Cuevas, E.; Medina, E. Nutrient dynamics within Amazonian forests. II. Fine root growth, nutrient availability, and leaf litter decomposition. Oecologia 1988, 76, 222–235. [Google Scholar] [CrossRef]
- Haines, B. Impact of leaf-cutting ants on vegetation development at Barro Colorado Island. In Tropical Ecological Systems: Trends in Terrestrial and Aquatic Research; Golley, F.G., Medina, E., Eds.; Springer: New York, NY, USA, 1975; pp. 99–101. [Google Scholar]
- Raich, J.W.; Riley, R.H.; Vitousek, P.M. Use of root-ingrowth cores to assess nutrient limitations in forest ecosystems. Can. J. For. Res. 1994, 24, 2135–2138. [Google Scholar] [CrossRef]
- St. John, T.V.; Coleman, D.C.; Reid, C.P. Growth and spatial distribution of nutrient-absorbing organs: Selective exploitation of soil heterogeneity. Plant Soil 1983, 71, 487–493. [Google Scholar] [CrossRef]
- Sayer, E.J.; Banin, L.F. Tree nutrient cycling in tropical forest—Lessons from fertilization experiments. In Tropical Tree Physiology: Adaptation and Responses in a Changing Environment; Goldstein, G., Santiago, L.S., Eds.; Tree Physiology Volume 6; Springer: New York, NY, USA, 2016; pp. 275–297. [Google Scholar]
- Goldin, S.R.; Hutchinson, M.F. Coarse woody debris modifies surface soils of degraded temperate eucalypt woodlands. Plant Soil 2013, 370, 461–469. [Google Scholar] [CrossRef]
- Lindsay, A.E.; Cunningham, S.A. Native grass establishment in grassy woodlands with nutrient-enriched soil and exotic grass invasion. Restor. Ecol. 2011, 19, 131–140. [Google Scholar] [CrossRef]
- Hafner, S.D.; Groffman, P.M. Soil nitrogen cycling under litter and coarse woody debris in a mixed forest in New York State. Soil Biol. Biochem. 2005, 37, 2159–2162. [Google Scholar] [CrossRef]
- Kwak, J.H.; Chang, S.X.; Naeth, M.A.; Schaaf, W. Coarse woody debris extract decreases nitrogen availability in two reclaimed oil sands soils in Canada. Ecol. Eng. 2015, 84, 13–21. [Google Scholar] [CrossRef]
- Spears, J.D.H.; Holub, S.M.; Harmon, M.E.; Lajtha, K. The influence of decomposing logs on soil biology and nutrient cycling in an old-growth mixed coniferous forest in Oregon, USA. Can. J. For. Res. 2003, 33, 2193–2201. [Google Scholar] [CrossRef]
- Zimmerman, J.K.; Pulliam, W.M.; Lodge, D.J.; Quiñones-Orfila, V.; Fetcher, N.; Guzman-Grajáles, S.; Parrotta, J.A.; Asbury, C.E.; Walker, L.R.; Waide, R.B. Nitrogen immobilization by decomposing woody debris and the recovery of tropical wet forest from hurricane damage. Oikos 1995, 72, 314–322. [Google Scholar] [CrossRef]
- Zalamea, M.; González, G.; Lodge, D.J. Physical, chemical and biological properties of soil under decaying wood in a tropical wet forest in Puerto Rico. Forests 2016, 7, 168. [Google Scholar] [CrossRef]
- Zalamea, M.; González, G.; Ping, C.L.; Michaelson, G. Soil organic matter dynamics under decaying Wood in a subtropical wet Forest: Effect of tree species and decay stage. Plant Soil 2007, 296, 173–185. [Google Scholar] [CrossRef]
- Clark, D.B.; Clark, D.A.; Brown, S.; Oberbauer, S.F.; Veldkamp, E. Stocks and flows of coarse woody debris across a tropical rain forest nutrient and topography gradient. For. Ecol. Manag. 2002, 164, 237–248. [Google Scholar] [CrossRef]
- Delaney, M.; Brown, S.; Lugo, A.E.; Torres-Lezama, A.; Quintero, N.B. The quantity and turnover of dead wood in permanent forest plots in six life zones of Venezuela. Biotropica 1998, 30, 2–11. [Google Scholar] [CrossRef]
- Sanford, R.L., Jr.; Parton, W.J.; Ojima, D.S.; Lodge, D.J. Hurricane effects on soil organic matter dynamics and forest production in the Luquillo Experimental Forest, Puerto Rico: Results of simulation modeling. Biotropica 1991, 23, 364–372. [Google Scholar] [CrossRef]
- Emanuel, K.A. Increasing destructiveness of tropical cyclones over the past 30 years. Nature 2005, 436, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Knutson, T.R.; McBride, J.L.; Chan, J.; Emanuel, K.; Holland, G.; Landsea, C.; Held, I.; Kossin, J.P.; Srivastava, A.K.; Sugi, M. Tropical cyclones and climate change. Nat. Geosci. 2010, 3, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Mei, W.; Xie, S.P. Intensification of landfalling typhoons over the northwest Pacific since the late 1970s. Nat. Geosci. 2016, 9, 753–757. [Google Scholar] [CrossRef]
- Bentz, B. Climate Change and bark beetles of the western United States and Canada: Direct and indirect effects. Bioscience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate change and forest disturbance. Bioscience 2001, 51, 723–734. [Google Scholar] [CrossRef]
- Brown, S.; Lugo, A.E.; Silander, S.; Liegel, L. Research History and Opportunities in the Luquillo Experimental Forest; General Technical Report, SO-44; United States Forest Service: Washington, DC, USA, 1983. [Google Scholar]
- Soil Survey Staff. Order 1 Soil Survey of the Luquillo Long-Term Ecological Research Grid, Puerto Rico; United States Department of Agriculture, National Resources Conservation Service: Lincoln, NE, USA, 1995.
- Lodge, D.J.; Cantrell, S.A.; González, G. Effects of canopy opening and debris deposition on fungal connectivity, phosphorus movement between litter cohorts and mass loss. For. Ecol. Manag. 2014, 332, 11–21. [Google Scholar] [CrossRef]
- Shanley, J.B.; Lodge, D.J.; Krabbenhoft, D.P.; Olson, M.L.; McDowell, W.H. New and old mercury fluxes from mercury amendments to Puerto Rico soil columns. In Presented at the American Geophysical Union Fall 2008 Meetings, San Francisco, CA, USA, 15–19 December 2008; Available online: http://adsabs.harvard.edu/abs/2008AGUFM.B13C0457S (accessed on 17 August 2009).
- Jenkinson, D.S.; Powlson, D.S. The effects of biocidal treatments on metabolism in soil—V: A method for measuring soil biomass. Soil Biol. Biochem. 1976, 8, 209–213. [Google Scholar] [CrossRef]
- Lin, Q.; Brookes, P.C. An evaluation of the substrate-induced respiration method. Soil Biol. Biochem. 1999, 31, 1969–1983. [Google Scholar] [CrossRef]
- Torres, J.A. Wood decomposition of Cyrilla racemiflora in a tropical montane forest. Biotropica 1994, 26, 124–140. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremmer, J.M. Automated instruments for determination of total carbon, nitrogen, and sulfur in soils by combustion techniques. In Soil Analysis, Modern Instruments Techniques; Marcel Dekker, Inc.: New York, NY, USA, 1991; pp. 261–286. [Google Scholar]
- Vitousek, P.M.; Matson, P.A. Mechanisms of nitrogen retention in forest ecosystems: A field experiment. Science 1984, 225, 51–52. [Google Scholar] [CrossRef] [PubMed]
- Newmann, E.I. A method of estimating the total length of root in a sample. J. App. Ecol. 1966, 3, 139–145. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT® 9.3 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2011. [Google Scholar]
- Harvey, A.E.; Jurgensen, M.F.; Larssen, M.J. Effects of Soil Organic Matter on Regeneration in Northern Rocky Mountain Forests; GTR-PNW-163; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1983; pp. 239–242.
- Price, S.P.; Bradford, M.A.; Ashton, M.S. Characterizing organic carbon stocks and flows in forest soils. In Managing Forest Carbon in a Changing Environment; Ashton, M.S., Tyrrell, M.L., Spalding, D., Gentry, B., Eds.; Springer Science and Business Media: New York, NY, USA, 2012; pp. 7–30. [Google Scholar]
- Zinn, Y.L.; Lal, R.; Bigham, J.M.; Resck, D.V.S. Edaphic controls on soil organic carbon retention in the Brazilian Cerrado: Texture and mineralogy. Soil Sci. Soc. Am. J. 2007, 71, 1204–1214. [Google Scholar] [CrossRef]
- Kaiser, K.; Guggenberger, G. The role of DOM sorption to mineral surfaces in the preservation of organic matter in soils. Org. Geochem. 2000, 31, 711–725. [Google Scholar] [CrossRef]
- Kayahara, G.J.; Klinka, K.; Lavkulich, L.M. Effects of decaying wood on eluviation, podzolization, acidification and nutrition in soils with different moisture regimes. Environ. Monit. Assess. 1991, 39, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Kalbitz, K.; Schwesig, D.; Rethemeyer, J.; Matzner, E. Stabilization of dissolved organic matter by sorption to the mineral soil. Soil Bio. Biochem. 2005, 37, 1319–1331. [Google Scholar] [CrossRef]
- Gutiérrez del Arroyo, O.; Silver, W.L. How deep does disturbance go? The legacy of hurricanes on tropical forest soil biogeochemistry. In Proceedings of the American Geophysical Union Meeting, San Francisco, CA, USA, 12–16 December 2016; Available online: https://agu.confex.com/agu/fm16/meetingapp.cgi/Paper/168890 (accessed on 21 October 2016).
- Turner, B.L.; Yavitt, J.B.; Harms, K.E.; Garcia, M.N.; Wright, S.J. Seasonal changes in soil organic matter after a decade of nutrient addition in a lowland tropical forest. Biogeochemistry 2015, 123, 221–235. [Google Scholar] [CrossRef]
- Cobb, T.P.; Hannam, K.D.; Kishchuk, B.E.; Langor, D.W.; Quideau, S.A.; Spence, J.R. Wood-feeding beetles and soil nutrient cycling in burned forests: Implications of post-fire salvage logging. Agric. For. Entomol. 2010, 12, 9–17. [Google Scholar] [CrossRef]
- Boddy, L.; Watkinson, S.C. Wood decomposition, higher fungi, and their role in nutrient redistribution. Can. J. Bot. 1995, 73, S1377–S1383. [Google Scholar] [CrossRef]
- Boddy, L. Saprotrophic cord-forming fungi: Meeting the challenge of heterogeneous environments. Mycologia 1999, 91, 13–32. [Google Scholar] [CrossRef]
- Ulyshen, M.D. Insect-mediated nitrogen dynamics in decomposing wood. Ecol. Entomol. 2015, 40 (Suppl. 1), 97–112. [Google Scholar] [CrossRef]
- Ausmus, B.S. Regulation of wood decomposition rates by arthropod and annelid populations. Ecol. Bull. 1977, 25, 180–192. [Google Scholar]
- Lang, G.E.; Knight, D.H. Decay rates for boles of tropical trees in Panama. Biotropica 1979, 11, 316–317. [Google Scholar] [CrossRef]
- Torres, J.A.; González, G. Wood Decomposition of Cyrilla racemiflora (Cyrillaceae) in Puerto Rican Dry and Wet Forests: A 13-year Case Study. Biotropica 2005, 37, 452–456. [Google Scholar] [CrossRef]
- Shiels, A.; González, G.; Lodge, D.J.; Willig, M.R.; Zimmerman, J.K. Cascading effects of canopy opening and debris deposition from a large-scale hurricane experiment in a tropical rainforest. Bioscience 2015, 65, 871–881. [Google Scholar] [CrossRef]
- Bouget, C.; Duelli, P. The effects of windthrow on forest insect communities: A literature review. Biol. Conserv. 2004, 118, 281–299. [Google Scholar] [CrossRef]
- Swift, M.J.; Boddy, L. Animal-microbial interactions in wood decomposition. In Invertebrate-Microbial Interactions; Anderson, J.M., Rayner, A.D.M., Walton, D.W.H., Eds.; Cambridge University Press: Cambridge, UK, 1984; pp. 89–131. [Google Scholar]
- Winsor, G.W.; Pollard, A.G. Carbon-nitrogen relationships in soil. IV. Mineralization of carbon and nitrogen. J. Sci. Food Agric. 1956, 7, 618–624. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a ‘Redfield’ ratio for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Zalamea, M.; González, G. Leaf fall phenology in a subtropical wet forest in Puerto Rico: From species to community patterns. Biotropica 2008, 40, 295–304. [Google Scholar] [CrossRef]
- González, G.; Lodge, D.J.; Richardson, B.A.; Richardson, M.J. A canopy trimming experiment in Puerto Rico: The response of litter decomposition and nutrient release to canopy opening and debris deposition in a subtropical wet forest. For. Ecol. Manag. 2014, 332, 32–46. [Google Scholar] [CrossRef]
- Silver, W.L.; Hall, S.J.; González, G. Differential effects of canopy trimming and litter deposition on litterfall and nutrient dynamics in a wet subtropical forest. For. Ecol. Manag. 2014, 332, 47–55. [Google Scholar] [CrossRef]
- Lodge, D.J.; Scatena, F.N.; Asbury, C.E.; Sánchez, M.J. Fine litterfall and related nutrient inputs resulting from Hurricane Hugo in subtropical wet and lower montane tropical forest of Puerto Rico. Biotropica 1991, 23, 336–342. [Google Scholar] [CrossRef]
- Zou, X.; Zucca, C.P.; Waide, R.B.; McDowell, W.H. Long-term influence of deforestation on tree species composition and litter dynamics of a tropical rain forest in Puerto Rico. For. Ecol. Manag. 1995, 78, 147–157. [Google Scholar] [CrossRef]
- Lodge, D.J. Nutrient cycling by fungi in wet tropical forests. In Aspects of Tropical Mycology; BMS Symposium Series 19; Isaac, S., Frankland, J.C., Watling, R., Whalley, A.J.S., Eds.; Cambridge University Press: Cambridge, UK, 1993; pp. 37–57. [Google Scholar]
- Bouskill, N.J.; Lim, H.C.; Borglin, S.; Salve, R.; Wood, T.E.; Silver, W.L. Pre-exposure to drought increases the resistance of tropical forest soil bacterial communities to extended drought. ISME J. 2013, 7, 384–394. [Google Scholar] [CrossRef] [PubMed]
- Bouskill, N.J.; Wood, T.E.; Baran, R.; Ye, Z.; Bowen, B.P.; Lim, H.C.; Zhou, J.; Van Norstrand, J.D.; Nico, P.; Northern, T.R.; et al. Belowground response to drought in a tropical forest soil. I. Changes in microbial functional potential and metabolism. Front. Microbiol. 2016, 7, 525. [Google Scholar] [CrossRef] [PubMed]
- Paul, E.A.; Clark, E.E. Soil Microbiology and Biochemistry; Academic Press: San Diego, CA, USA, 1996. [Google Scholar]
- Khan, D.; Sahito, Z.A.; Dawar, S.; Zaki, M.J. Frass of saproxylic-cerambycid larvae from dead twigs of Acacia stenophylla A. Cunn. EX. Benth. and its effects on germination and seedling growth of Lactuca sativa L. var. grand rapids. Int. J. Biol. Biotechnol. 2016, 13, 461–470. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Block | Latitude, Longitude | Soil Type |
|---|---|---|---|
| Q. Sonadora watershed | 1 | 18°19′24′′ N, 65°49′03′′ W | Cristal clay-clay loam |
| Q. Prieta watershed in LFDP cells 02.09 and 07.10 | 2 | 18°19′30.7′′ N, 65°48′56.6′′ W 18°19′31′′ N, 65°49′02′′ W | Cristal clay-clay loam |
| Q. Sonadora watershed | 3 | 18°19′24′′ N, 65°49′04′′ W | Cristal clay-clay loam |
| Q. Prieta watershed in LFDP cells 10.15 and 10.16 | 4 | 18°19′37′′ N, 65°49′00′′ W 18°19′27′′ N, 65°49′00′′ W | Coloso clay |
| Location | Label | Hurricane | Tree Species | Diam. (cm) | Decay Class | Decay Rate | Slope (%) |
|---|---|---|---|---|---|---|---|
| Q. Sonadora | H.1 | Hugo 1989 | G. guidonia | 35–55 | 2 | Slow | 28 |
| Q. Sonadora | G.1 | Georges 1998 | G. guidonia | 38–80 | 1 | Slow | 35 |
| Q. Prieta, LFDP cell 02.09 | H.2 | Hugo 1989 | D. excelsa | 35–52 | 3 | Fast | 35 |
| Q. Prieta, LFDP cell 07.10 | G.2 | Georges 1998 | D. excelsa | 30–35 | 2 | Fast | 40 |
| Q. Sonadora | H.3 | Hugo 1989 | D. excelsa | 30–43 | 2 | Moderate | 20 |
| Q. Sonadora | G.3 | Georges 1998 | D. excelsa | 30–55 | 1 | Slow | 40 |
| LFDP cell 10.16 | H.4 | Hugo 1989 | G. guidonia | 39–53 | 3 | Fast | 25 |
| LFDP cell 10.15 | G.4 | Georges 1998 | G. guidonia | 45–70 | 2 | Fast | 25 |
| Variable (s) | Levels | Probability |
|---|---|---|
| 0–10 cm soil horizon | ||
| Carbon (total %) Lognormal, Identity Link function, Repeated Measures GLIMMIX, HQIC fit statistics, all possible interactions included in model (shown if significant) | ||
| Position (Under vs. Away) | 2 | 0.0324 * |
| Hurricane within Block (Hugo vs. Georges) | 2 | 0.3264 |
| Hillslope direction (Upslope vs. Downslope) | 2 | 0.8188 |
| Time (0.6, 1.4 and 2 years post-Georges) | 3 | 0.0374 * |
| Block (log number, Hurricane nested within Block) true replicates | 4 | |
| Nitrogen (total %) Lognormal, Identity Link function, Repeated Measures GLIMMIX, HQIC fit statistics, all possible interactions included in model (shown if significant) | ||
| Position (Under vs. Away) | 2 | 0.0020 ** |
| Hurricane within Block (Hugo vs. Georges) | 2 | 0.2177 |
| Hillslope direction (Upslope vs. Downslope) | 2 | 0.0420 * |
| Time (0.6, 1.4 and 2 years post-Georges) | 3 | 0.6816 |
| Block (log number, Hurricane nested within Block) true replicates | 4 | |
| Microbial biomass C (mg·kg−1 soil) Gaussian, Identity Link function, Repeated Measures GLIMMIX, HQIC fit statistics, all possible interactions included (shown if significant) | ||
| Position (Under vs. Away) | 2 | 0.0840 sug. |
| Hurricane within Block (Hugo vs. Georges) | 2 | 0.9038 |
| Hillslope direction (Upslope vs. Downslope) | 2 | 0.8660 |
| Time (0.6, 1.4 and 2 years post-Georges) | 3 | <0.001 *** |
| Block (log number, Hurricane nested within Block) true replicates | 4 | |
| Root length (total) Lognormal, Identity Link function, Repeated Measures GLIMMIX, HQIC fit statistics, all possible interactions included (shown if significant) | ||
| Position (Under vs. Away) | 2 | 0.9049 |
| Hurricane within Block (Hugo vs. Georges) | 2 | 0.0440 * |
| Hillslope direction (Upslope vs. Downslope) | 2 | 0.0250 * |
| Time (0.6, 1.4 and 2 years post-Georges) | 2 | 0.0182 * |
| Block (log number, Hurricane nested within Block) true replicates, deleted one incomplete block | 3 | |
| Position by Time interaction | 1 DF | 0.0368 * |
| 10–20 cm soil horizon | ||
| Carbon (total %) Lognormal, Identity Link function, Repeated Measures GLIMMIX, HQIC fit statistics, all possible interactions included (shown if significant) | ||
| Position (Under vs. Away) | 2 | 0.0273 * |
| Hurricane within Block (Hugo vs. Georges) | 2 | 0.5899 |
| Hillslope direction (Upslope vs. Downslope) | 2 | 0.7656 |
| Time (0.6, and 1.4 years post-Georges) | 2 | 0.1673 |
| Block (log number, Hurricane nested within Block) true replicates | 4 | |
| Hurricane by position interaction | 0.0870 sug. | |
| Nitrogen (total %) Lognormal, Identity Link function, Repeated Measures GLIMMIX, HQIC fit statistics, all possible interactions included (shown if significant) | ||
| Position (Under vs. Away) | 2 | 0.0188 * |
| Hurricane within Block (Hugo vs. Georges) | 2 | 0.7820 |
| Hillslope direction (Upslope vs. Downslope) | 2 | 0.8711 |
| Time (0.6, and 1.4 years post-Georges) | 2 | 0.0579 sug. |
| Block (log number, Hurricane nested within Block) true replicates | 4 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lodge, D.J.; Winter, D.; González, G.; Clum, N. Effects of Hurricane-Felled Tree Trunks on Soil Carbon, Nitrogen, Microbial Biomass, and Root Length in a Wet Tropical Forest. Forests 2016, 7, 264. https://doi.org/10.3390/f7110264
Lodge DJ, Winter D, González G, Clum N. Effects of Hurricane-Felled Tree Trunks on Soil Carbon, Nitrogen, Microbial Biomass, and Root Length in a Wet Tropical Forest. Forests. 2016; 7(11):264. https://doi.org/10.3390/f7110264
Chicago/Turabian StyleLodge, D. Jean, Dirk Winter, Grizelle González, and Naomi Clum. 2016. "Effects of Hurricane-Felled Tree Trunks on Soil Carbon, Nitrogen, Microbial Biomass, and Root Length in a Wet Tropical Forest" Forests 7, no. 11: 264. https://doi.org/10.3390/f7110264
APA StyleLodge, D. J., Winter, D., González, G., & Clum, N. (2016). Effects of Hurricane-Felled Tree Trunks on Soil Carbon, Nitrogen, Microbial Biomass, and Root Length in a Wet Tropical Forest. Forests, 7(11), 264. https://doi.org/10.3390/f7110264