
Increasing Water Use Efficiency Comes at a Cost for Norway Spruce


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
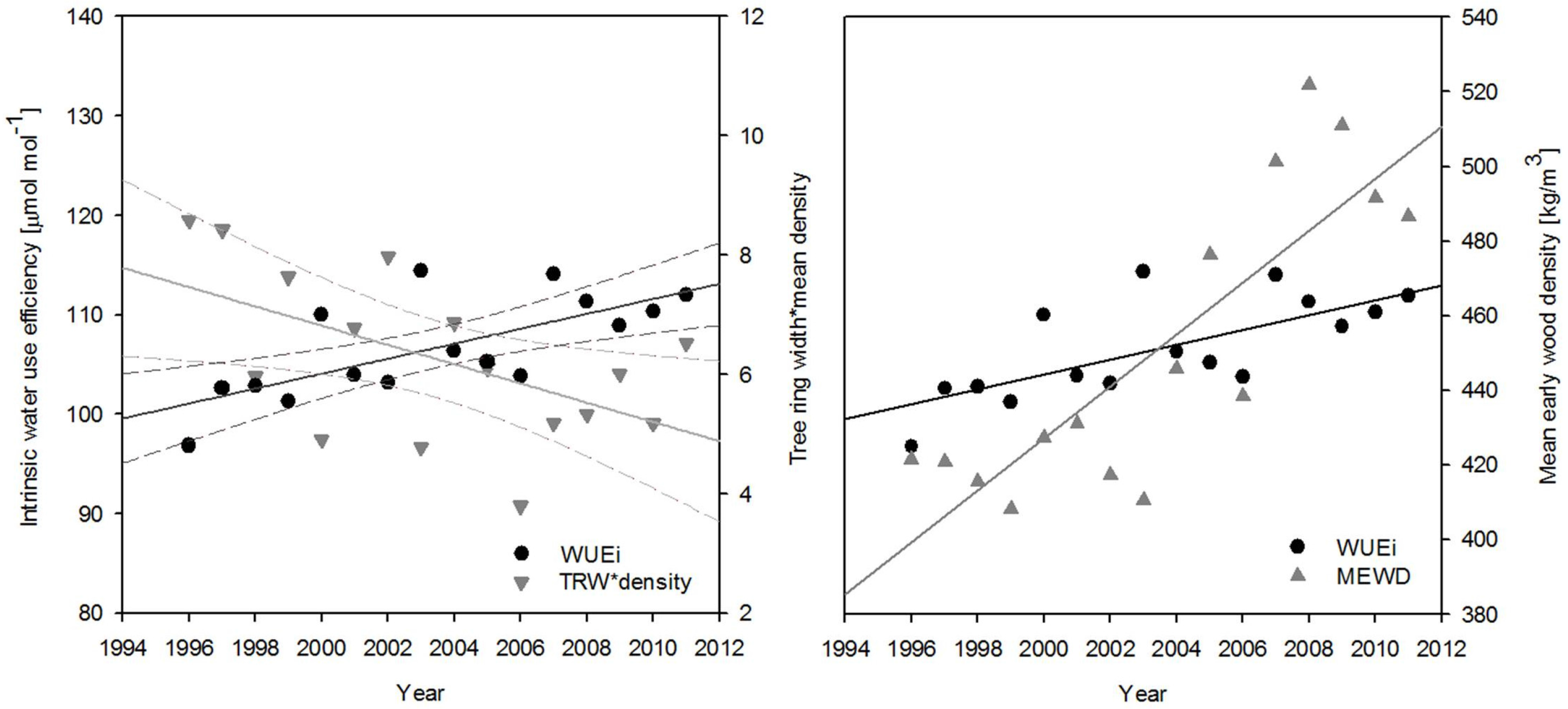
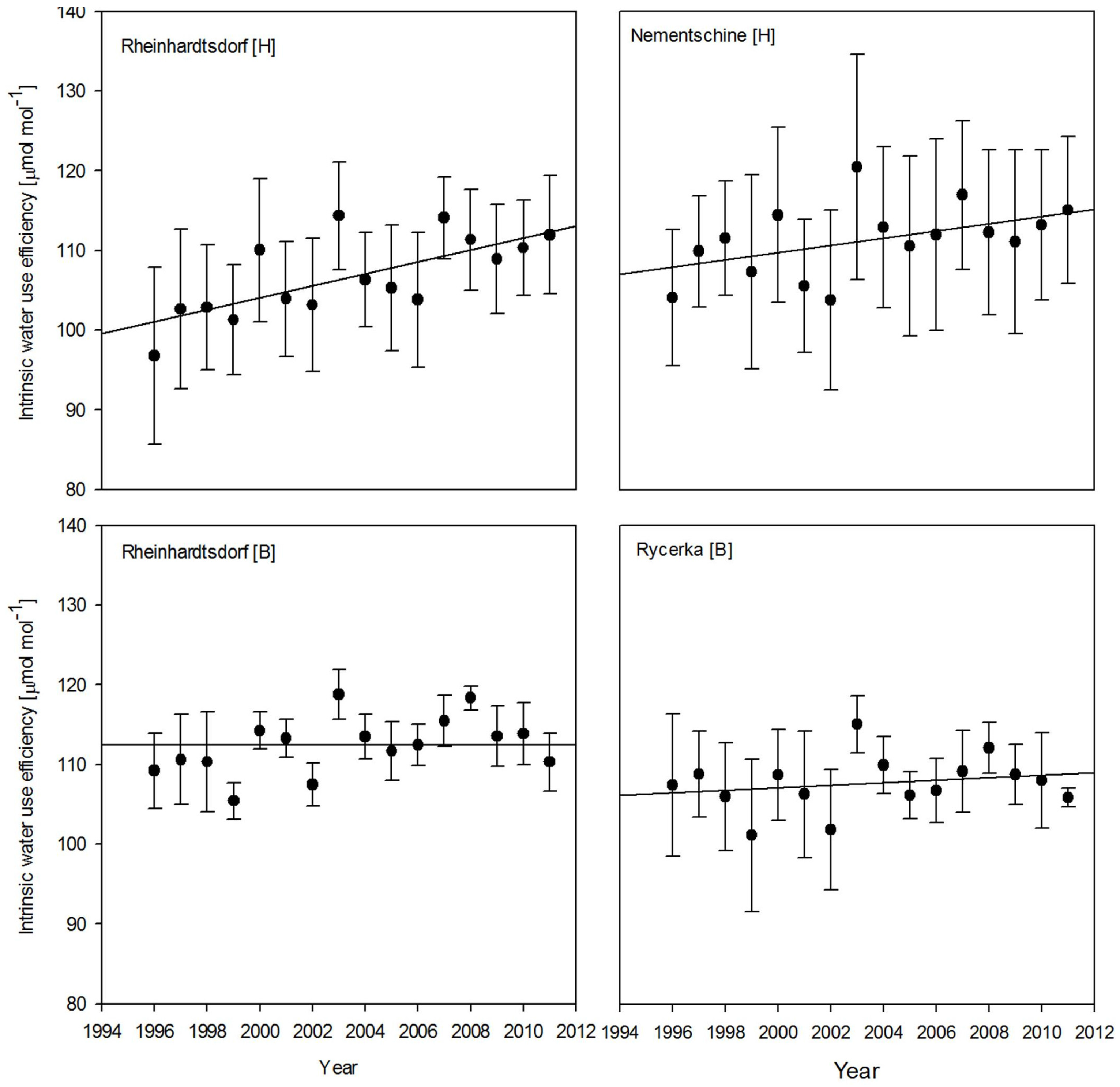
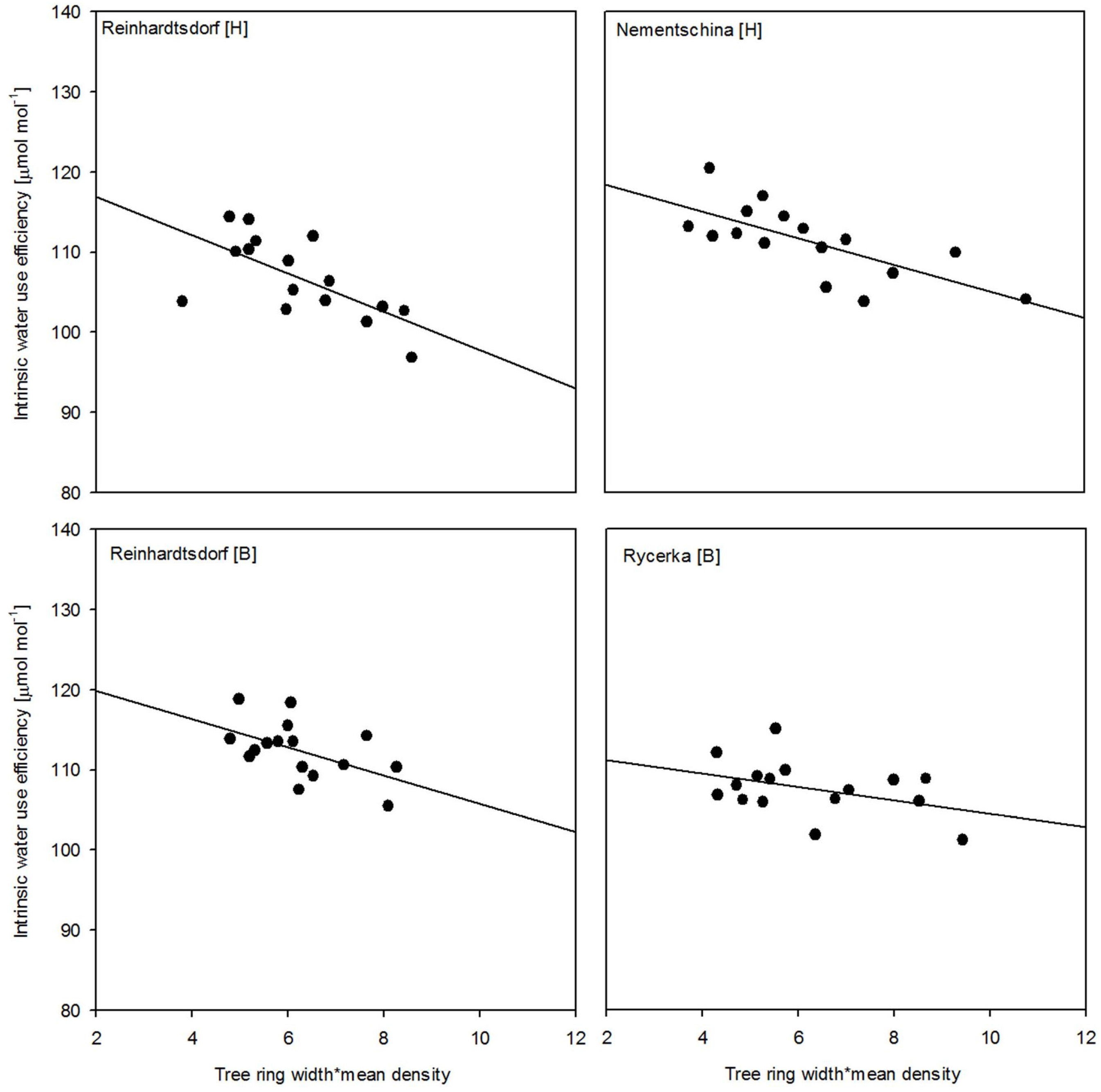
3. Results
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fardusi, M.J.; Ferrio, J.P.; Comas, C.; Voltas, J.; de Dios, V.R.; Serrano, L. Intra-specific association between carbon isotope composition and productivity in woody plants: A meta-analysis. Plant Sci. 2016, 251, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Norby, R.J.; Delucia, E.H.; Gielen, B.; Calfapietra, C.; Giardina, C.P.; King, J.S.; Ledford, J.; McCarthy, H.R.; Moore, D.J.; Ceulemans, R. Forest response to elevated CO2 is conserved across a broad range of productivity. Proc. Natl. Acad. Sci. USA 2005, 102, 18052–18056. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.A.; Pataki, D.E.; Körner, C.; Clark, H.; Del Grosso, S.J.; Grünzweig, J.M.; Knapp, A.K.; Mosier, A.R.; Newton, P.C.D.; Niklaus, P.A.; et al. Water relations in grassland and desert ecosystems exposed to elevated atmospheric CO2. Oecologia 2004, 140, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Wullschleger, S.D.; Post, W.; King, A. On the potential for a CO2 fertilization effect in forests: Estimates of the biotic growth factor based on 58 controlled-exposure studies. In Biotic Feedbacks in the Global Climatic System; Woodwell, G.M., Mackenzie, F.T., Eds.; Oxford University Press: New York, NY, USA, 1995; pp. 85–107. [Google Scholar]
- Körner, C.; Asshoff, R.; Bignucolo, O.; Hättenschwiler, S.; Keel, S.G.; Peláez-Riedl, S.; Pepin, S.; Siegwolf, R.T.W.; Zotz, G. Carbon Flux and Growth in Mature Deciduous Forest Trees Exposed to Elevated CO2. Science 2005, 309, 1360–1362. [Google Scholar] [CrossRef] [PubMed]
- HÄTtenschwiler, S.; Miglietta, F.; Raschi, A.; KÖRner, C. Thirty years of in situ tree growth under elevated CO2: A model for future forest responses? Glob. Chang. Biol. 1997, 3, 463–471. [Google Scholar] [CrossRef]
- Pastor, J.; Post, W. Response of northern forests to CO2-induced climate change. Nature 1988, 334, 55–58. [Google Scholar] [CrossRef]
- Morison, J.I.L. Sensitivity of stomata and water use efficiency to high CO2. Plant Cell Environ. 1985, 8, 467–474. [Google Scholar] [CrossRef]
- Keenan, T.F.; Hollinger, D.Y.; Bohrer, G.; Dragoni, D.; Munger, J.W.; Schmid, H.P.; Richardson, A.D. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 2013, 499, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.-H.; Lim, S.-S.; Lee, K.-S.; Viet, H.D.; Matsushima, M.; Lee, K.-H.; Jung, K.; Kim, H.-Y.; Lee, S.-M.; Chang, S.X. Temperature and air pollution affected tree ring δ13C and water-use efficiency of pine and oak trees under rising CO2 in a humid temperate forest. Chem. Geol. 2016, 420, 127–138. [Google Scholar] [CrossRef]
- Bert, D.; Leavitt, S.W.; Dupouey, J.-L. Variations Of Wood Δ13c And Water-Use Efficiency Of Abies Alba During The Last Century. Ecology 1997, 78, 1588–1596. [Google Scholar]
- Rydval, M.; Wilson, R. The impact of industrial SO2 pollution on north Bohemia conifers. Water Air Soil Pollut. 2012, 223, 5727–5744. [Google Scholar] [CrossRef]
- Meesenburg, H.; Ahrends, B.; Fleck, S.; Wagner, M.; Fortmann, H.; Scheler, B.; Klinck, U.; Dammann, I.; Eichhorn, J.; Mindrup, M.; et al. Long-term changes of ecosystem services at Solling, Germany: Recovery from acidification, but increasing nitrogen saturation? Ecol. Indic. 2016, 65, 103–112. [Google Scholar] [CrossRef]
- Schmitz, A.; Sanders, T.G. Deposition. 2016. Available online: http://blumwald.thuenen.de/level-ii/auswertungen/deposition/ (accessed on 30 August 2016).
- Peñuelas, J.; Canadell, J.G.; Ogaya, R. Increased water-use efficiency during the 20th century did not translate into enhanced tree growth. Glob. Ecol. Biogeogr. 2011, 20, 597–608. [Google Scholar] [CrossRef]
- Monclus, R.; Dreyer, E.; Villar, M.; Delmotte, F.M.; Delay, D.; Petit, J.-M.; Barbaroux, C.; Le Thiec, D.; Bréchet, C.; Brignolas, F. Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides × Populus nigra. New Phytol. 2006, 169, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Hubick, K.T.; Condon, A.G.; Richards, R.A. Carbon Isotope Fractionation and Plant Water-Use Efficiency. In Stable Isotopes in Ecological Research; Rundel, P.W., Ehleringer, J.R., Nagy, K.A., Eds.; Springer: New York, NY, USA, 1989; pp. 21–40. [Google Scholar]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Eamus, D. The interaction of rising CO2 and temperatures with water use efficiency. Plant Cell Environ. 1991, 14, 843–852. [Google Scholar] [CrossRef]
- Silva, L.C.; Horwath, W.R. Explaining global increases in water use efficiency: Why have we overestimated responses to rising atmospheric CO2 in natural forest ecosystems? PLoS ONE 2013, 8, e53089. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.C.; Anand, M. Probing for the influence of atmospheric CO2 and climate change on forest ecosystems across biomes. Glob. Ecol. Biogeogr. 2013, 22, 83–92. [Google Scholar] [CrossRef]
- Schweingruber, F.H. Der Jahrring; Standort, Methodik, Zeit und Klima in der Dendrochronologie; Haupt: Bern, Switzerland, 1983; Volume 234. [Google Scholar]
- Loader, N.; Robertson, I.; Barker, A.; Switsur, V.; Waterhouse, J. An improved technique for the batch processing of small wholewood samples to α-cellulose. Chem. Geol. 1997, 136, 313–317. [Google Scholar] [CrossRef]
- Wilson, A.; Grinsted, M. 12C/13C in cellulose and lignin as palaeothermometers. Nature 1977, 265, 133–135. [Google Scholar] [CrossRef]
- Schweingruber, F.; Fritts, H.; Bräker, O.; Drew, L.; Schär, E. The X-ray technique as applied to dendroclimatology. Tree-Ring Bull. 1978, 38, 61. [Google Scholar]
- Linares, J.C.; Camarero, J.J. From pattern to process: Linking intrinsic water-use efficiency to drought-induced forest decline. Glob. Chang. Biol. 2012, 18, 1000–1015. [Google Scholar] [CrossRef]
- Sanvill, P.; Sandels, A. The influence of early respacing on the wood density of Sitka spruce. Forestry 1983, 56, 109–120. [Google Scholar]
- Bouriaud, O.; Leban, J.-M.; Bert, D.; Deleuze, C. Intra-annual variations in climate influence growth and wood density of Norway spruce. Tree Physiol. 2005, 25, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Polley, H.W.; Johnson, H.B.; Marino, B.; Mayeux, H. Increase in C3 plant water-use efficiency and biomass over Glacial to present CO2 concentrations. Nature 1993, 361, 61. [Google Scholar]
- Mousseau, M.; Saugier, B. The Direct Effect of Increased CO2 on Gas Exchange and Growth of Forest Tree Species. J. Exp. Bot. 1992, 43, 1121–1130. [Google Scholar] [CrossRef]
- Yu, G.; Song, X.; Wang, Q.; Liu, Y.; Guan, D.; Yan, J.; Sun, X.; Zhang, L.; Wen, X. Water-use efficiency of forest ecosystems in eastern China and its relations to climatic variables. New Phytol. 2008, 177, 927–937. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, S.B.; Nosal, M.; Wullschleger, S.D.; Sun, G. Interactive effects of ozone and climate on tree growth and water use in a southern Appalachian forest in the USA. New Phytol. 2007, 174, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.; Bytnerowicz, A.; Thorstenson, Y.R. Effects of air pollutants on the composition of stable carbon isotopes, δ13C, of leaves and wood, and on leaf injury. Plant Physiol. 1988, 88, 218–223. [Google Scholar] [CrossRef] [PubMed]




© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanders, T.G.M.; Heinrich, I.; Günther, B.; Beck, W. Increasing Water Use Efficiency Comes at a Cost for Norway Spruce. Forests 2016, 7, 296. https://doi.org/10.3390/f7120296
Sanders TGM, Heinrich I, Günther B, Beck W. Increasing Water Use Efficiency Comes at a Cost for Norway Spruce. Forests. 2016; 7(12):296. https://doi.org/10.3390/f7120296
Chicago/Turabian StyleSanders, Tanja G M, Ingo Heinrich, Björn Günther, and Wolfgang Beck. 2016. "Increasing Water Use Efficiency Comes at a Cost for Norway Spruce" Forests 7, no. 12: 296. https://doi.org/10.3390/f7120296
APA StyleSanders, T. G. M., Heinrich, I., Günther, B., & Beck, W. (2016). Increasing Water Use Efficiency Comes at a Cost for Norway Spruce. Forests, 7(12), 296. https://doi.org/10.3390/f7120296