
Short-Term vs. Long-Term Effects of Understory Removal on Nitrogen and Mobile Carbohydrates in Overstory Trees


Abstract
:1. Introduction
2. Material and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Tissue Sampling
2.4. Total Nitrogen and NSC (Soluble Sugars and Starch) Analysis
2.5. Data Analysis
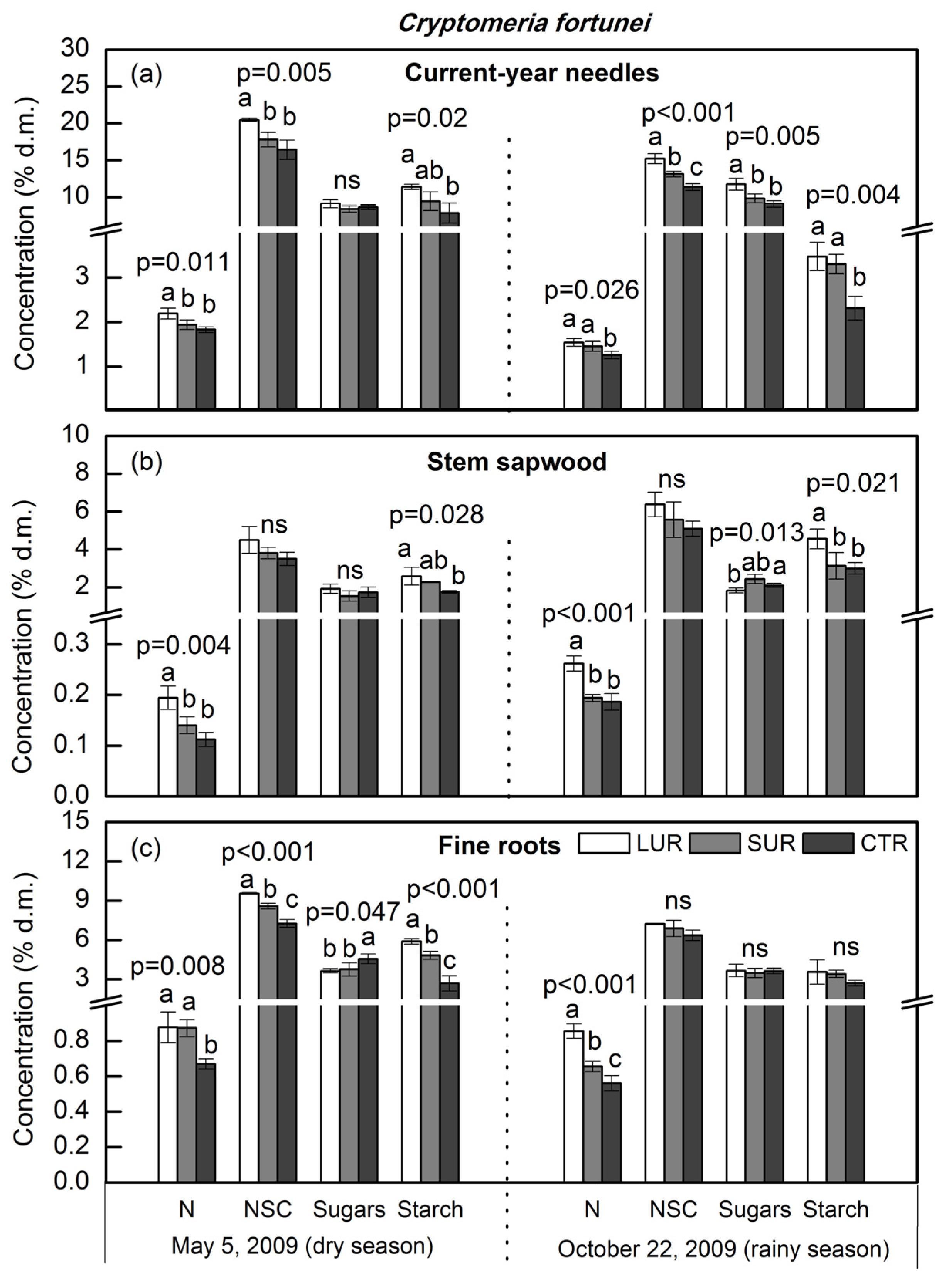
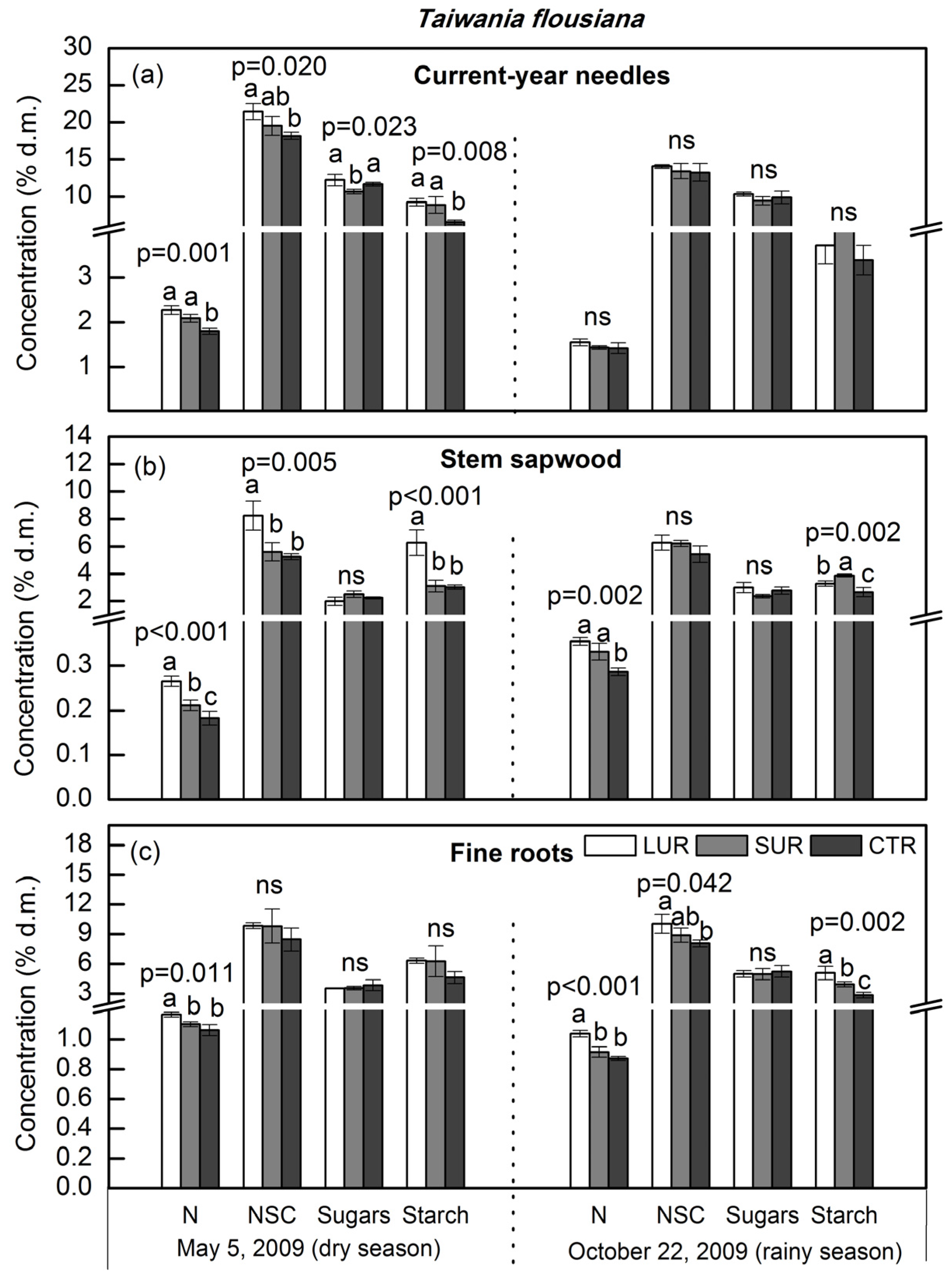
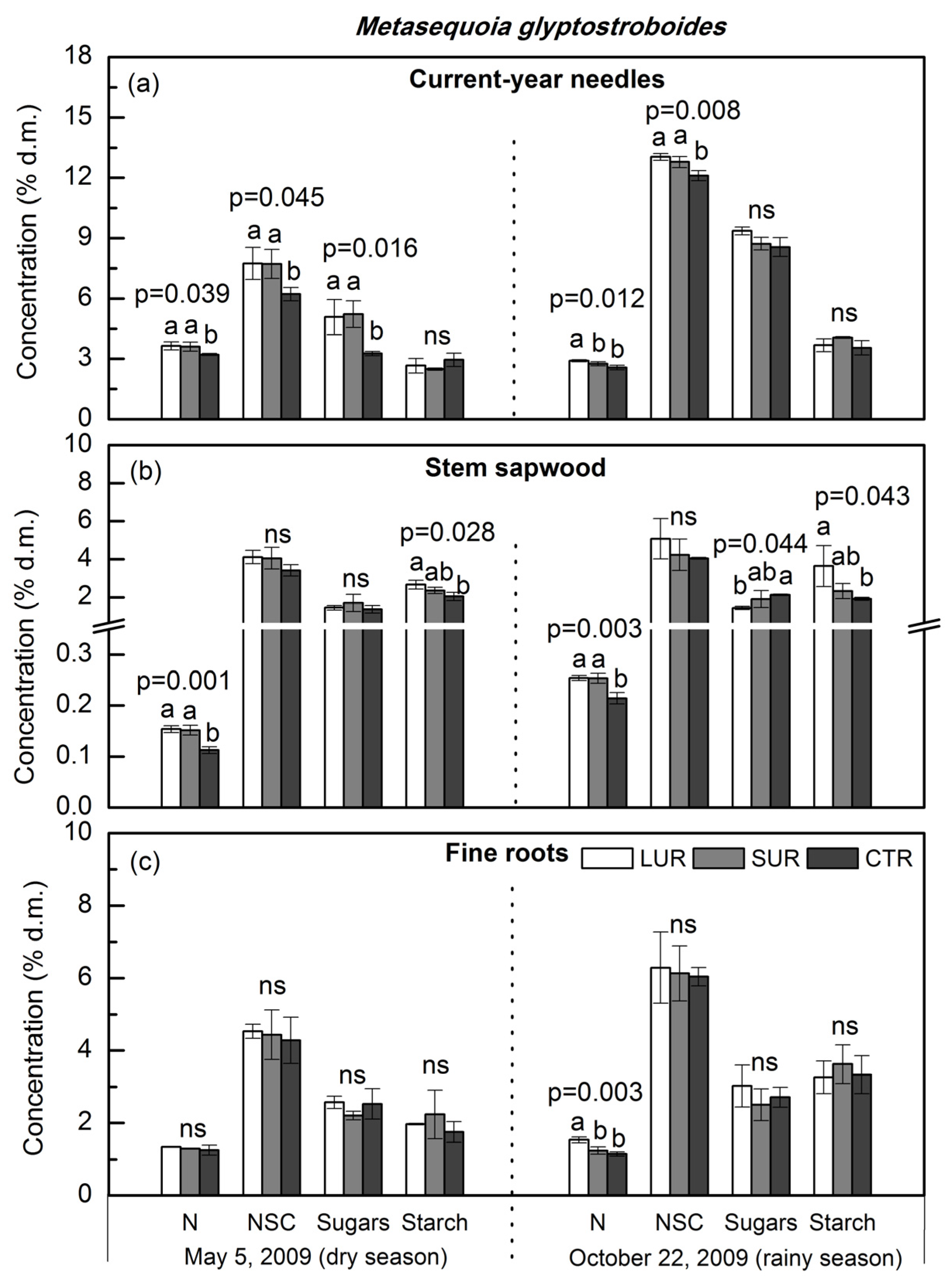
3. Results
4. Discussion
4.1. Tissue Nitrogen and Mobile Carbohydrates in Overstory Trees
4.2. Dry Season vs. Wet Season
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brooker, R.W. Plant-plant interactions and environmental change. New Phytol. 2006, 171, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Craine, J.M.; Dybzinski, R. Mechanisms of plant competition for nutrients, water and light. Funct. Ecol. 2013, 27, 833–840. [Google Scholar] [CrossRef]
- Silvertown, J. Plant coexistence and the niche. Trend. Ecol. Evol. 2004, 19, 605–611. [Google Scholar] [CrossRef]
- Hart, S.A.; Chen, H.Y.H. Understory vegetation dynamics of North American boreal forests. Crit. Rev. Plant Sci. 2006, 25, 381–397. [Google Scholar] [CrossRef]
- Sagar, R.; Raghubanshi, A.S.; Singh, J.S. Comparison of community composition and species diversity of understorey and overstorey tree species in a dry tropical forest of northern India. J. Environ. Manag. 2008, 88, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Oren, R.; Waring, R.H.; Stafford, S.G.; Barrett, J.W. Twenty-four Years of Ponderosa Pine Growth in Relation to Canopy Leaf Area and Understory Competition. For. Sci. 1987, 33, 538–547. [Google Scholar]
- Takahashi, K. Regeneration and coexistence of two subalpine conifer species in relation to dwarf bamboo in the understorey. J. Veg. Sci. 1997, 8, 529–536. [Google Scholar] [CrossRef]
- O’Brien, M.J.; O’Hara, K.L.; Erbilgin, N.; Wood, D.L. Overstory and shrub effects on natural regeneration processes in native Pinus radiata stands. For. Ecol. Manag. 2007, 240, 178–185. [Google Scholar] [CrossRef]
- Barnes, P.W.; Archer, S. Tree-shrub interactions in a subtropical savanna parkland: Competition or facilitation? J. Veg. Sci. 1999, 10, 525–536. [Google Scholar] [CrossRef]
- Parker, W.C.; Dey, D.C. Influence of overstory density on ecophysiology of red oak (Quercus rubra) and sugar maple (Acer saccharum) seedlings in central Ontario shelterwoods. Tree Physiol. 2008, 28, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Shainsky, L.J.; Rose, C.L. Effects of competition on the foliar chemistry of young Douglas-fir in monoculture and mixed stands with young red alder. Can. J. For. Res. 1995, 25, 1969–1977. [Google Scholar] [CrossRef]
- Matsushima, M.; Chang, S.X. Effects of understory removal, N fertilization, and litter layer removal on soil N cycling in a 13-year-old white spruce plantation infested with Canada bluejoint grass. Plant Soil 2007, 292, 243–258. [Google Scholar] [CrossRef]
- Nilsson, M.C.; Wardle, D.A. Understory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest. Front Ecol. Environ. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- Gilliam, F.S. The ecological significance of the herbaceous layer in temperate forest ecosystems. BioScience 2007, 57, 845–858. [Google Scholar] [CrossRef]
- Elliott, K.J.; Vose, J.M.; Knoepp, J.D.; Clinton, B.D.; Kloeppel, B.D. Functional Role of the Herbaceous Layer in Eastern Deciduous Forest Ecosystems. Ecosystems 2015, 18, 221–236. [Google Scholar] [CrossRef]
- Miller, B.J.; Clinton, P.W.; Buchan, G.D.; Robson, A.B. Transpiration rates and canopy conductance of Pinus radiata growing with different pasture understories in agroforestry systems. Tree Physiol. 1998, 18, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Kume, A.; Satomura, T.; Tsuboi, N.; Chiwa, M.; Hanba, Y.T.; Nakane, K.; Horikoshi, T.; Sakugawa, H. Effects of understory vegetation on the ecophysiological characteristics of an overstory pine, Pinus densiflora. For. Ecol. Manag. 2003, 176, 195–203. [Google Scholar] [CrossRef]
- Li, M.H.; Hoch, G.; Körner, C. Source/sink removal affects mobile carbohydrates in Pinus cembra at the Swiss treeline. Trees-Struct. Funct. 2002, 16, 331–337. [Google Scholar] [CrossRef]
- Hoch, G.; Körner, C. The carbon charging of pines at the climatic treeline: Aglobal comparison. Oecologia 2003, 135, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. Carbon limitation in trees. J. Ecol. 2003, 91, 4–17. [Google Scholar] [CrossRef]
- Li, M.H.; Xiao, W.F.; Shi, P.L.; Wang, S.G.; Zhong, Y.D.; Liu, X.L.; Wang, X.D.; Cai, X.H.; Shi, Z.M. Nitrogen and carbon source-sink relationships in trees at the Himalayan treelines compared with lower elevations. Plant Cell Environ. 2008, 31, 1377–1387. [Google Scholar] [CrossRef] [PubMed]
- Li, M.H.; Xiao, W.F.; Wang, S.G.; Cheng, G.W.; Cherubini, P.; Cai, X.H.; Liu, X.L.; Wang, X.D.; Zhu, W.Z. Mobile carbohydrates in Himalayan treeline trees I. Evidence for carbon gain limitation but not for growth limitation. Tree Physiol. 2008, 28, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Li, M.H.; Hoch, G.; Körner, C. Spatial variability of mobile carbohydrates within Pinus cembra trees at the alpine treeline. Phyton-Ann. Rei Bot. A. 2001, 41, 203–213. [Google Scholar]
- Hoch, G.; Popp, M.; Körner, C. Altitudinal increase of mobile carbon pools in Pinus cembra suggests sink limitation of growth at the Swiss treeline. Oikos 2002, 98, 361–374. [Google Scholar] [CrossRef]
- Li, M.H.; Du, Z.; Pan, H.; Yan, C.; Xiao, W.; Lei, J. Effects of neighboring woody plants on target trees with emphasis on effects of understorey shrubs on overstorey physiology in forest communities: a mini-review. Community Ecol. 2012, 13, 117–128. [Google Scholar] [CrossRef]
- Graham, E.A.; Mulkey, S.S.; kitajima, K.; Phillips, N.G.; Wright, S.J. Cloud cover limits net CO2 uptake and growth of a rainforest tree during tropical rainy seasons. Proc. Natl. Acad. Sci. USA 2003, 100, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Nambiar, E.K.S.; Sands, R. Competition for Water and Nutrients in Forests. Can. J. For. Res. 1993, 23, 1955–1968. [Google Scholar] [CrossRef]
- Matsushima, M.; Chang, S.X. Vector analysis of understory competition, N fertilization, and litter layer removal effects on white spruce growth and nutrition in a 13-year-old plantation. For. Ecol. Manag. 2006, 236, 332–341. [Google Scholar] [CrossRef]
- Chang, S.X.; Preston, C.M. Understorey competition affects tree growth and fate of fertilizer-applied 15N in a coastal British Columbia plantation forest: 6-year results. Can. J. For. Res. 2000, 30, 1379–1388. [Google Scholar] [CrossRef]
- Staples, T.E.; van Rees, K.C.J.; van Kessel, C. Nitrogen competition using 15N between early successional plants and planted white spruce seedlings. Can. J. For. Res. 1999, 29, 1282–1289. [Google Scholar] [CrossRef]
- Hangs, R.D.; Knight, J.D.; van Rees, K.C.J. Nitrogen accumulation by conifer seedlings and competitor species from 15N-labeled controlled-release fertilizer. Soil Sci. Soc. Am. J. 2003, 67, 300–308. [Google Scholar] [CrossRef]
- Woods, P.V.; Nambiar, E.K.S.; Smethurst, P.J. Effect of Annual Weeds on Water and Nitrogen Availability to Pinus-Radiata Trees in a Young Plantation. For. Ecol. Manag. 1992, 48, 145–163. [Google Scholar] [CrossRef]
- Ginn, S.E.; Seiler, J.R.; Cazell, B.H.; Kreh, R.E. Physiological and Growth-Responses of 8-Year-Old Loblolly-Pine Stands to Thinning. For. Sci. 1991, 37, 1030–1040. [Google Scholar]
- Groninger, J.W.; Seiler, J.R.; Peterson, J.A.; Kreh, R.E. Growth and photosynthetic responses of four Virginia Piedmont tree species to shade. Tree Physiol. 1996, 16, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Binkley, D.; Stape, J.L.; Ryan, M.G. Thinking about efficiency of resource use in forests. For. Ecol. Manag. 2004, 193, 5–16. [Google Scholar] [CrossRef]
- Martindale, W.; Leegood, R.C. Acclimation of photosynthesis to low temperature in Spinacia oleracea L. 2. Effects of nitrogen supply. J. Exp. Bot. 1997, 48, 1873–1880. [Google Scholar] [CrossRef]
- Davey, P.A.; Parsons, A.J.; Atkinson, L.; Wadge, K.; Long, S.P. Does photosynthetic acclimation to elevated CO2 increase photosynthetic nitrogen-use efficiency? A study of three native UK grassland species in open-top chambers. Funct. Ecol. 1999, 13, 21–28. [Google Scholar] [CrossRef]
- Kitajima, K.; Hogan, K.P. Increases of chlorophyll a/b ratios during acclimation of tropical woody seedlings to nitrogen limitation and high light. Plant Cell Environ. 2003, 26, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Espejo, A.; Nicolas, E.; Fernandez, J.E. Seasonal evolution of diffusional limitations and photosynthetic capacity in olive under drought. Plant Cell Environ. 2007, 30, 922–933. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.D.; Newton, M.; Zedaker, S.M. Influence of Ceanothus velutinus and Associated Forbs on the Water-Stress and Stemwood Production of Douglas-Fir. For. Sci. 1998, 34, 333–343. [Google Scholar]
- Carter, G.A.; Miller, J.H.; Davis, D.E.; Patterson, R.M. Effect of Vegetative Competition on the Moisture and Nutrient Status of Loblolly-Pine. Can. J. For. Res. 1984, 14, 1–9. [Google Scholar] [CrossRef]
- Elliott, K.J.; White, A.S. Competitive Effects of Various Grasses and Forbs on Ponderosa Pine-Seedlings. For. Sci. 1987, 33, 356–366. [Google Scholar]
- Sands, R.; Nambiar, E.K.S. Water Relations of Pinus radiata in Competition with Weeds. Can. J. For. Res. 1984, 14, 233–237. [Google Scholar] [CrossRef]
- Watt, M.S.; Whitehead, D.; Mason, E.G.; Richardson, B.; Kimberley, M.O. The influence of weed competition for light and water on growth and dry matter partitioning of young Pinus radiata, at a dryland site. For. Ecol. Manag. 2003, 183, 363–376. [Google Scholar] [CrossRef]
- Nambiar, E.K.S.; Zed, P.G. Influence of Weeds on the Water Potential, Nutrient Content and Growth of Young Radiata Pine. Aust. For. Res. 1980, 10, 279–288. [Google Scholar]
- Balandier, P.; Collet, C.; Miller, J.H.; Reynolds, P.E.; Zedaker, S.M. Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighbouring vegetation. Forestry 2006, 79, 3–27. [Google Scholar] [CrossRef]
- Casper, B.B.; Jackson, R.B. Plant competition underground. Annu. Rev. Ecol. Syst. 1997, 28, 545–570. [Google Scholar] [CrossRef]
- Montgomery, R.A.; Reich, P.B.; Palik, B.J. Untangling positive and negative biotic interactions: views from above and below ground in a forest ecosystem. Ecology 2010, 91, 3641–3655. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.K.; Sumida, A.; Ono, K.; Shibata, H.; Uemura, S.; Takahashi, K.; Hara, T. The effects of understorey dwarf bamboo (Sasa kurilensis) removal on soil fertility in a Betula ermanii forest of northern Japan. Ecol. Res. 2006, 21, 315–320. [Google Scholar] [CrossRef]
- Tripathi, S.K.; Sumida, A.; Shibata, H.; Uemura, S.; Ono, K.; Hara, T. Growth and substrate quality of fine root and soil nitrogen availability in a young Betula ermanii forest of northern Japan: Effects of the removal of understory dwarf bamboo (Sasa kurilensis). For. Ecol. Manag. 2005, 212, 278–290. [Google Scholar] [CrossRef]
- Takahashi, K.; Uemura, S.; Suzuki, J.I.; Hara, T. Effects of understory dwarf bamboo on soil water and the growth of overstory trees in a dense secondary Betula ermanii forest, northern Japan. Ecol. Res. 2003, 18, 767–774. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Plots (n) | Slope exposure | Soil humidity | Age (year) | Height | Diameter | Treatments | |||
|---|---|---|---|---|---|---|---|---|---|
| H (m) | m/year | dhb (cm) | cm/year | ||||||
| Cryptomeria fortunei plantation in the Gan-Gou Valley | |||||||||
| LUR | 3 | W | Mesic | 16 | 10.9 ± 2.6 | 0.68 | 14.4 ± 3.0 | 0.90 | Long-term understory removal |
| SUR | 3 | W | Mesic | 16 | 8.9 ± 2.5 | 0.56 | 10.6 ± 3.5 | 0.66 | Short-term understory removal |
| CUR | 3 | W | Mesic | 16 | 9.8 ± 2.1 | 0.61 | 11.8 ± 3.3 | 0.74 | Understory intact |
| Taiwania flousiana plantation in the Gan-Gou Valley | |||||||||
| LUR | 3 | W | Mesic | 16 | 8.2 ± 1.6 | 0.51 | 14.1 ± 3.3 | 0.88 | Long-term understory removal |
| SUR | 3 | W | Mesic | 16 | 8.0 ± 3.5 | 0.50 | 10.5 ± 3.9 | 0.66 | Short-term understory removal |
| CUR | 3 | W | Mesic | 16 | 8.0 ± 2.7 | 0.50 | 10.8 ± 3.2 | 0.68 | Understory intact |
| Metasequoia glyptostroboides plantation in the Dou-Peng Valley | |||||||||
| LUR | 3 | NW | Mesic | 29 | 19.8 ± 2.7 | 0.68 | 27.9 ± 5.2 | 0.96 | Long-term understory removal |
| SUR | 3 | NW | Mesic | 29 | 22.6 ± 1.3 | 0.78 | 32.3 ± 4.8 | 1.11 | Short-term understory removal |
| CUR | 3 | NW | Mesic | 29 | 21.3 ± 2.8 | 0.73 | 30.6 ± 5.9 | 1.06 | Understory intact |
| N | NSC | Sugars | Starch | NSC-N ratio | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p | F | p | F | p | F | p | F | p | |||
| Cryptomeria fortunei | |||||||||||||
| C-needles | Season (S) | 1 | 155.07 | 0.000 | 194.34 | 0.000 | 35.82 | 0.000 | 304.66 | 0.000 | 144.93 | 0.000 | |
| Type (T) | 2 | 16.66 | 0.000 | 40.89 | 0.000 | 15.14 | 0.001 | 13.30 | 0.001 | 17.66 | 0.000 | ||
| S*T | 2 | 1.06 | ns | 0.21 | ns | 6.36 | 0.013 | 3.65 | ns | 1.17 | ns | ||
| P-needles | Season (S) | 1 | 101.14 | 0.000 | 44.07 | 0.000 | 1.22 | ns | 274.53 | 0.000 | 138.89 | 0.000 | |
| Type (T) | 2 | 17.26 | 0.000 | 36.49 | 0.000 | 19.15 | 0.000 | 28.77 | 0.000 | 19.33 | 0.000 | ||
| S*T | 2 | 0.23 | ns | 1.73 | ns | 1.24 | ns | 1.56 | ns | 2.43 | ns | ||
| Sapwood | Season (S) | 1 | 74.10 | 0.000 | 37.79 | 0.000 | 13.57 | 0.003 | 45.78 | 0.000 | 73.81 | 0.000 | |
| Type (T) | 2 | 40.65 | 0.000 | 5.67 | 0.019 | 0.35 | ns | 12.59 | 0.001 | 32.90 | 0.000 | ||
| S*T | 2 | 0.68 | ns | 0.08 | ns | 7.46 | 0.008 | 2.70 | ns | 4.61 | 0.033 | ||
| Fine roots | Season (S) | 1 | 24.03 | 0.000 | 34.92 | 0.000 | 5.31 | 0.040 | 28.20 | 0.000 | 37.01 | 0.000 | |
| Type (T) | 2 | 38.11 | 0.000 | 11.01 | 0.002 | 3.21 | ns | 26.55 | 0.000 | 58.81 | 0.000 | ||
| S*T | 2 | 5.82 | 0.017 | 2.18 | ns | 2.52 | ns | 8.56 | 0.005 | 13.56 | 0.001 | ||
| Taiwania flousiana | |||||||||||||
| C-needles | Season (S) | 1 | 213.07 | 0.000 | 188.48 | 0.000 | 38.82 | 0.000 | 257.10 | 0.000 | 217.90 | 0.000 | |
| Type (T) | 2 | 18.59 | 0.000 | 6.95 | 0.010 | 7.36 | 0.008 | 12.74 | 0.001 | 28.43 | 0.000 | ||
| S*T | 2 | 7.50 | 0.008 | 2.58 | ns | 0.63 | ns | 6.38 | 0.013 | 0.52 | ns | ||
| P-needles | Season (S) | 1 | 1.64 | ns | 520.93 | 0.000 | 80.50 | 0.000 | 188.84 | 0.000 | 0.57 | ns | |
| Type (T) | 2 | 10.82 | 0.002 | 17.09 | 0.000 | 2.79 | ns | 9.29 | 0.004 | 12.83 | 0.001 | ||
| S*T | 2 | 0.37 | ns | 7.94 | 0.006 | 1.08 | ns | 8.56 | 0.005 | 0.55 | ns | ||
| Sapwood | Season (S) | 1 | 290.04 | 0.000 | 1.78 | ns | 15.31 | 0.002 | 15.73 | 0.002 | 266.28 | 0.000 | |
| Type (T) | 2 | 50.65 | 0.000 | 15.01 | 0.001 | 0.16 | ns | 27.64 | 0.000 | 42.39 | 0.000 | ||
| S*T | 2 | 2.13 | ns | 7.49 | 0.008 | 7.85 | 0.007 | 26.30 | 0.000 | 7.26 | 0.009 | ||
| Roots | Season (S) | 1 | 211.91 | 0.000 | 0.65 | ns | 48.07 | 0.000 | 24.67 | 0.000 | 651.56 | 0.000 | |
| Type (T) | 2 | 42.62 | 0.000 | 4.37 | 0.038 | 0.87 | ns | 10.52 | 0.002 | 124.74 | 0.000 | ||
| S*T | 2 | 3.64 | 0.058 | 0.46 | ns | 0.01 | ns | 0.73 | ns | 15.70 | 0.000 | ||
| Metasequoia glyptostroboides | |||||||||||||
| C-needles | Season (S) | 1 | 131.27 | 0.000 | 552.87 | 0.000 | 325.95 | 0.000 | 64.29 | 0.000 | 145.54 | 0.000 | |
| Type (T) | 2 | 12.56 | 0.001 | 11.33 | 0.002 | 11.13 | 0.002 | 0.25 | ns | 7.82 | 0.007 | ||
| S*T | 2 | 0.95 | ns | 1.11 | ns | 4.61 | 0.033 | 4.58 | 0.033 | 0.74 | ns | ||
| Sapwood | Season (S) | 1 | 647.68 | 0.000 | 3.99 | ns | 5.90 | 0.032 | 1.40 | ns | 915.76 | 0.000 | |
| Type (T) | 2 | 44.42 | 0.000 | 2.86 | ns | 2.93 | ns | 9.01 | 0.004 | 112.53 | 0.000 | ||
| S*T | 2 | 0.01 | ns | 0.59 | ns | 3.08 | ns | 2.32 | ns | 34.42 | 0.000 | ||
| Roots | Season (S) | 1 | 0.10 | ns | 27.53 | 0.000 | 4.28 | ns | 32.27 | 0.000 | 1.12 | ns | |
| Type (T) | 2 | 13.49 | 0.001 | 0.40 | ns | 1.47 | ns | 0.10 | ns | 9.54 | 0.003 | ||
| S*T | 2 | 5.71 | 0.018 | 0.24 | ns | 0.28 | ns | 0.93 | ns | 5.21 | 0.024 | ||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Z.; Cai, X.; Bao, W.; Chen, H.; Pan, H.; Wang, X.; Zhao, Q.; Zhu, W.; Liu, X.; Jiang, Y.; et al. Short-Term vs. Long-Term Effects of Understory Removal on Nitrogen and Mobile Carbohydrates in Overstory Trees. Forests 2016, 7, 67. https://doi.org/10.3390/f7030067
Du Z, Cai X, Bao W, Chen H, Pan H, Wang X, Zhao Q, Zhu W, Liu X, Jiang Y, et al. Short-Term vs. Long-Term Effects of Understory Removal on Nitrogen and Mobile Carbohydrates in Overstory Trees. Forests. 2016; 7(3):67. https://doi.org/10.3390/f7030067
Chicago/Turabian StyleDu, Zhong, Xiaohu Cai, Weikai Bao, Huai Chen, Hongli Pan, Xue Wang, Qingxia Zhao, Wanze Zhu, Xingliang Liu, Yong Jiang, and et al. 2016. "Short-Term vs. Long-Term Effects of Understory Removal on Nitrogen and Mobile Carbohydrates in Overstory Trees" Forests 7, no. 3: 67. https://doi.org/10.3390/f7030067
APA StyleDu, Z., Cai, X., Bao, W., Chen, H., Pan, H., Wang, X., Zhao, Q., Zhu, W., Liu, X., Jiang, Y., & Li, M. -H. (2016). Short-Term vs. Long-Term Effects of Understory Removal on Nitrogen and Mobile Carbohydrates in Overstory Trees. Forests, 7(3), 67. https://doi.org/10.3390/f7030067