
Variation in Mountain Vegetation Composition between the East and the West Sides of Southern Taiwan

Abstract
:1. Introduction
2. Materials and Methods
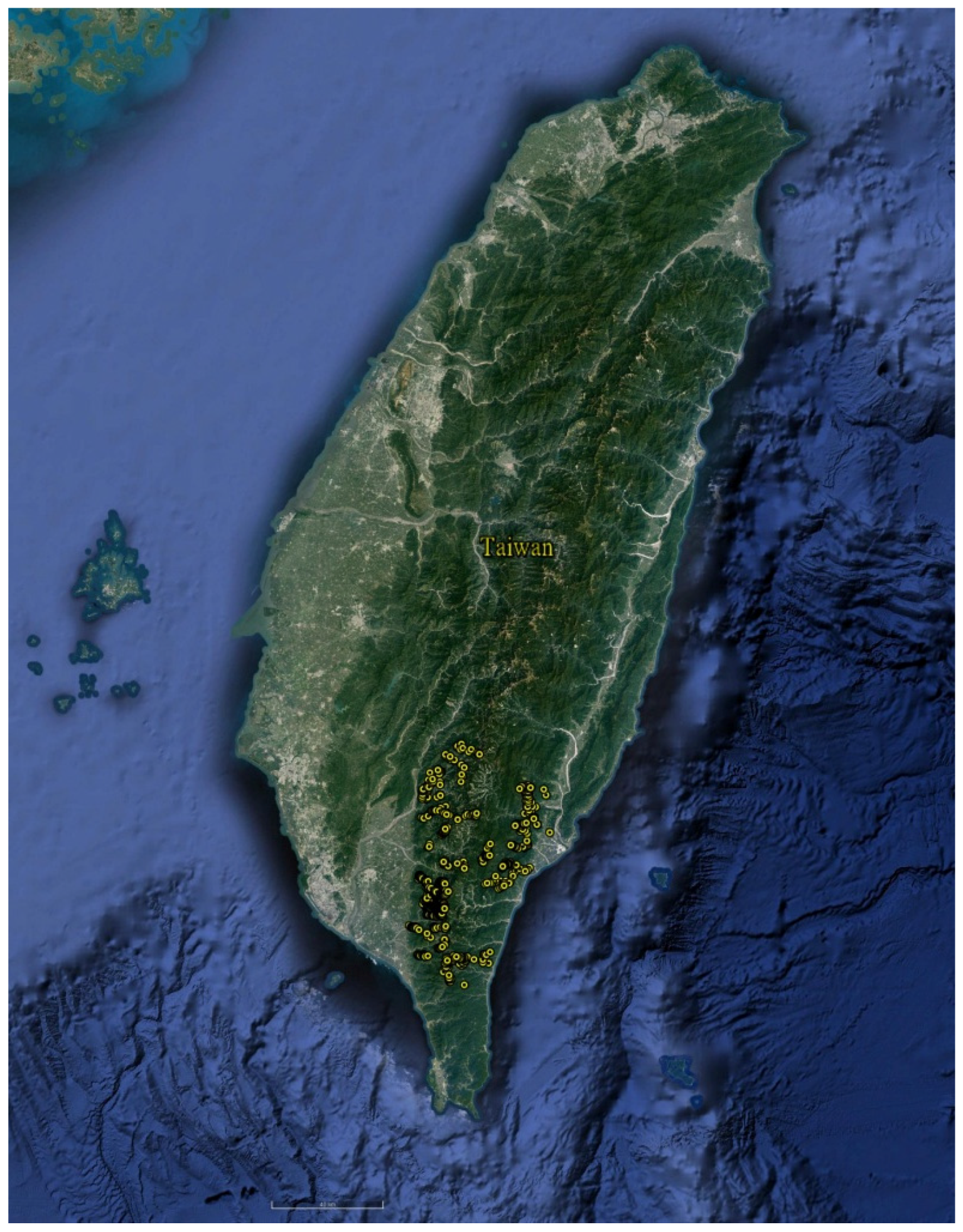
2.1. Study Area
2.2. Vegetation Data and Environmental Variables
2.3. Classification of Vegetation Types
2.4. Ordination, Synoptic Table and Software
3. Results
3.1. Vascular Plants and Rare Plant Species
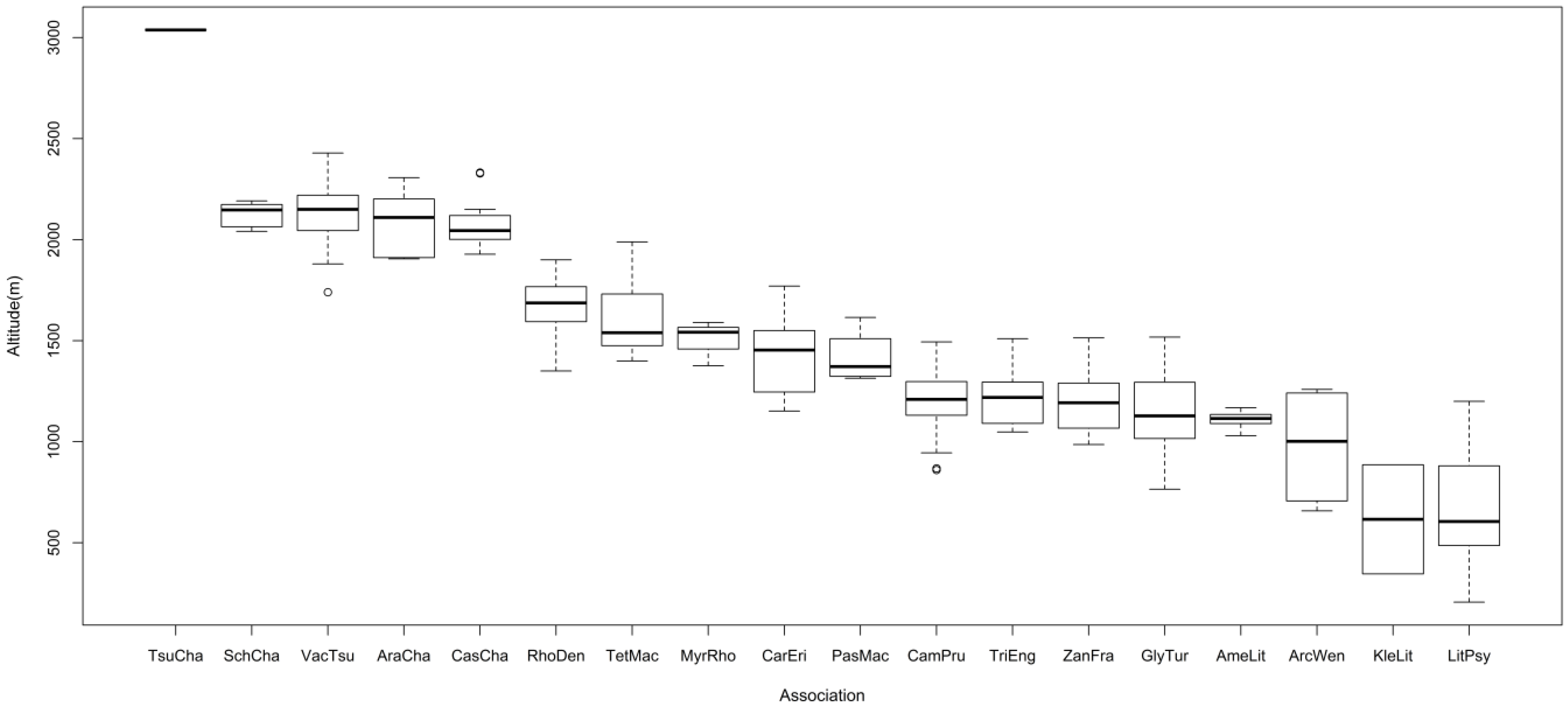
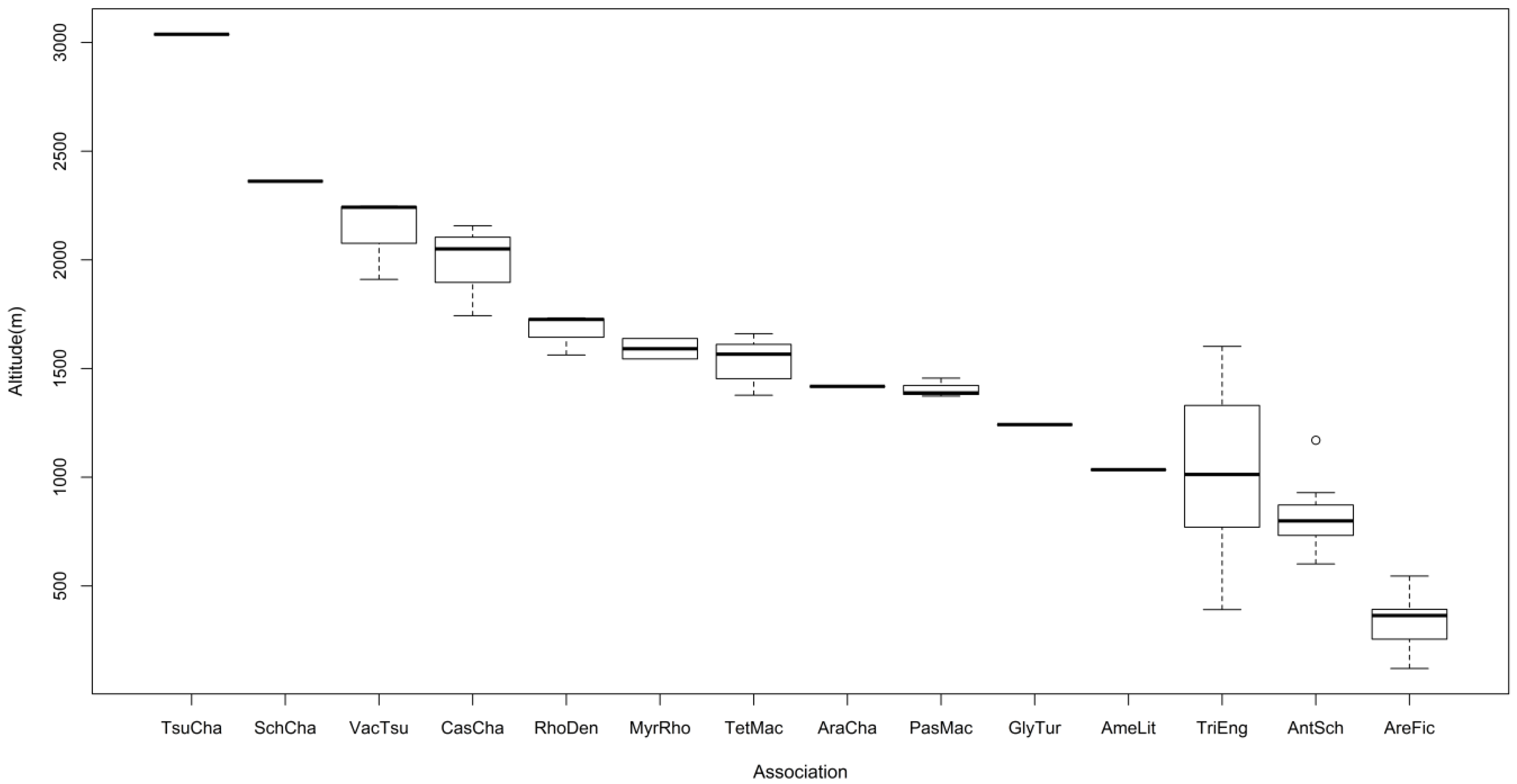
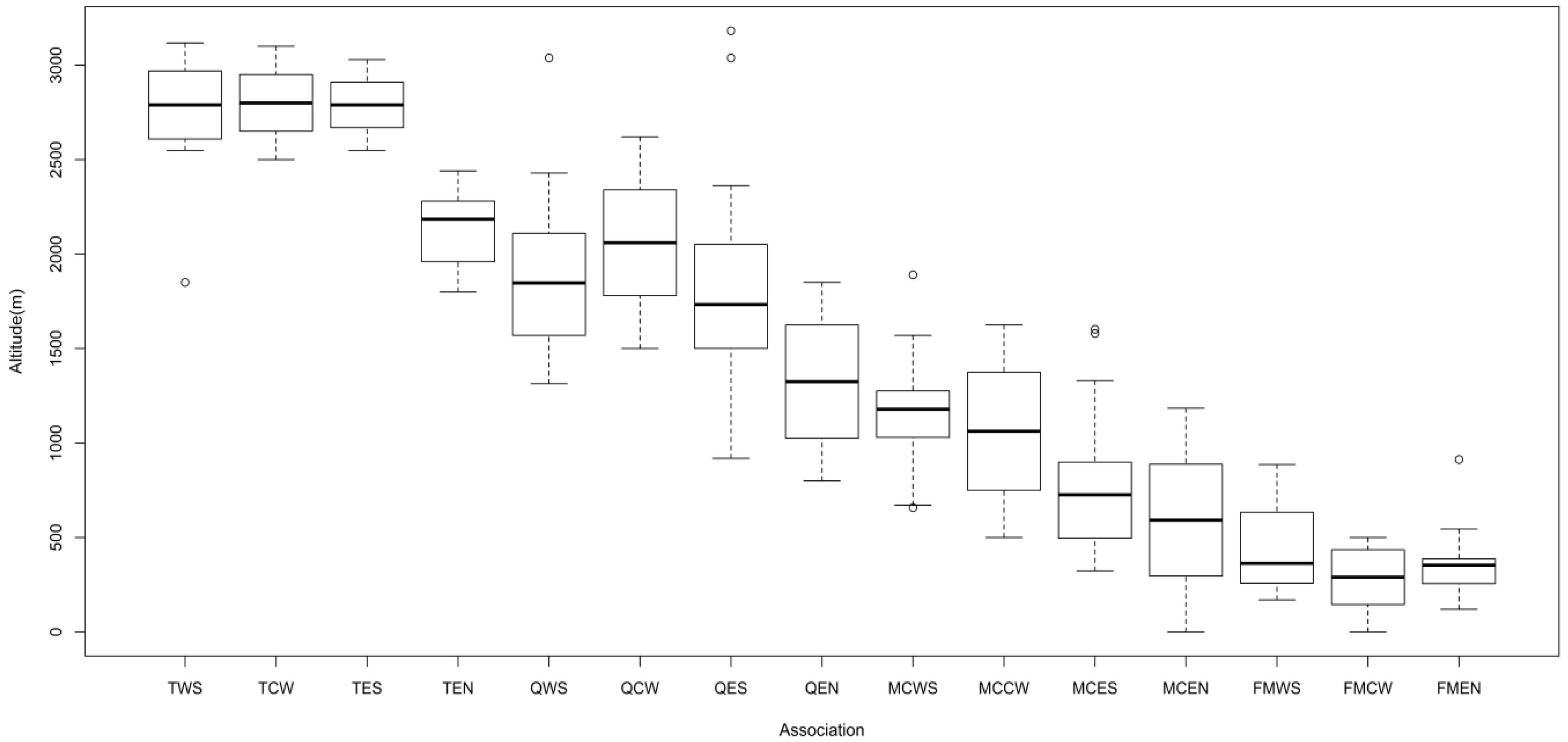
3.2. Vegetation Types
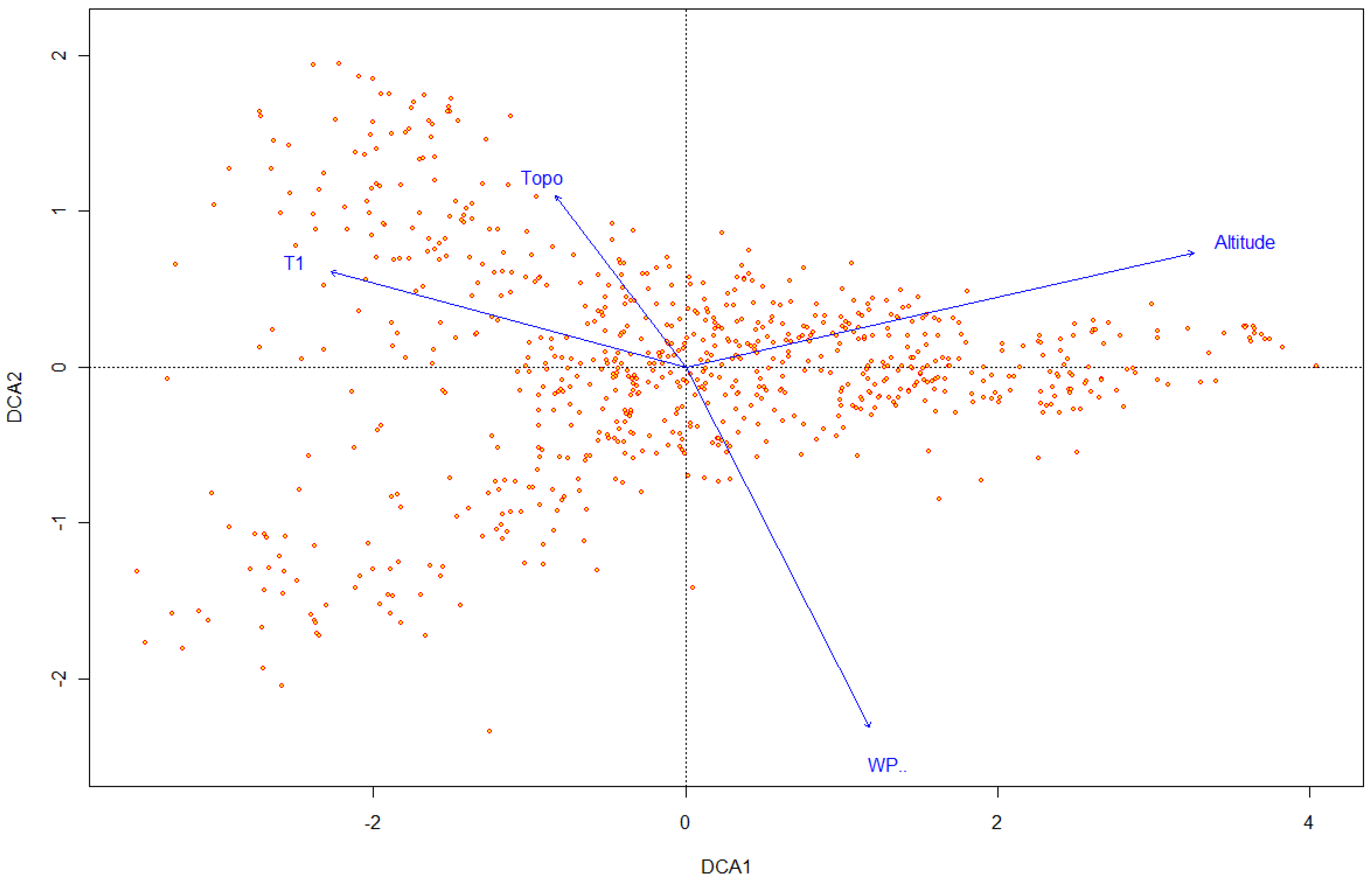
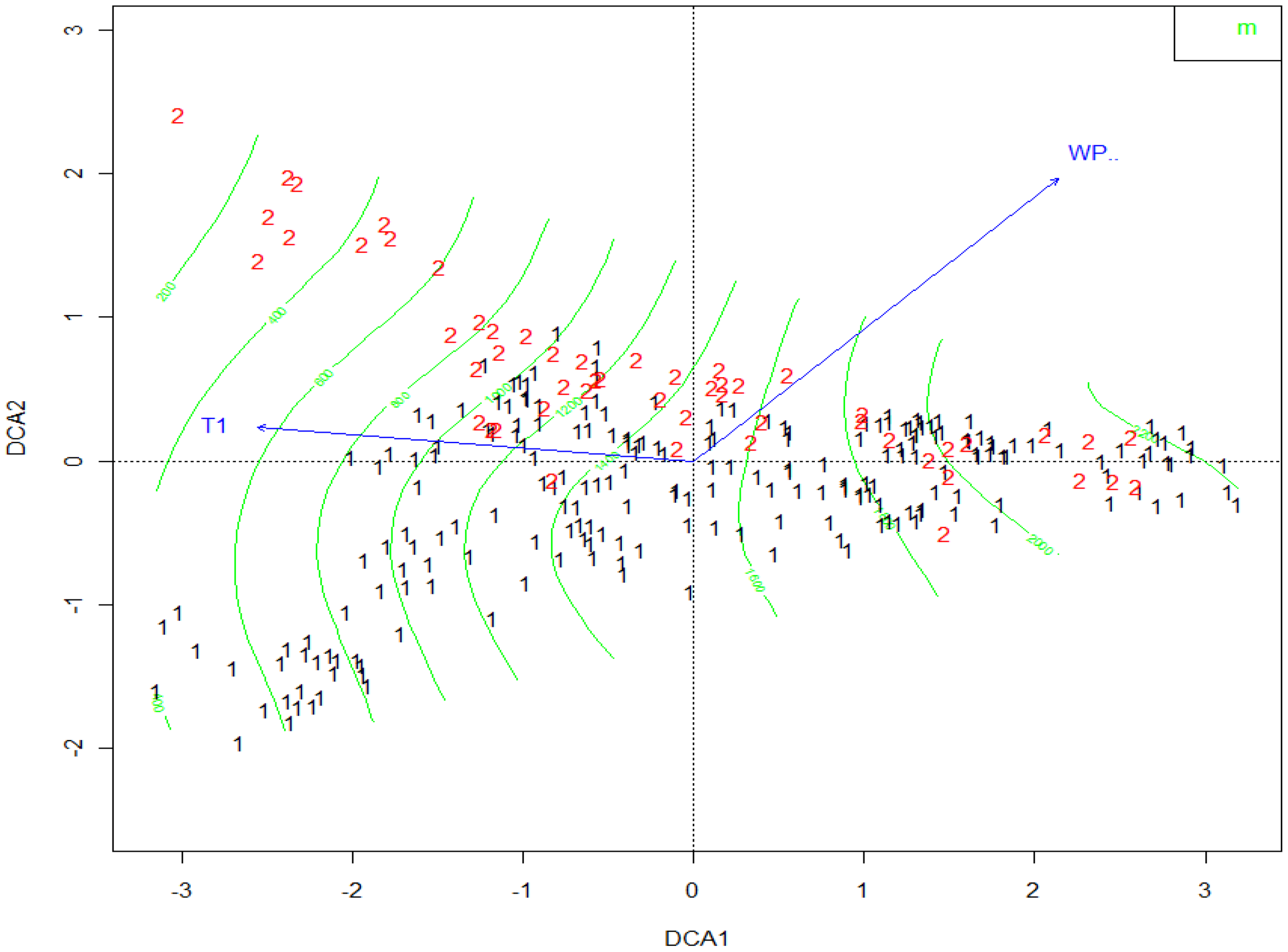
3.3. Relationship between Vegetation and Environmental Factors
4. Discussion
Relationship between Distribution of Vegetation and Environmental Factors
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, C.F.; Chytrý, M.; Zeleny, D.; Chen, M.Y.; Chen, T.Y.; Chiou, C.R.; Hsia, Y.J.; Liu, H.Y.; Yang, S.Z.; Yeh, C.L.; et al. Classification of Taiwan forest vegetation. Appl. Veg. Sci. 2013, 16, 698–719. [Google Scholar] [CrossRef]
- Su, H.J. Studies on the climate and vegetation types of the natural forests in Taiwan (III): A scheme of geographical climatic regions. Q. J. Chin. For. 1985, 18, 33–44. [Google Scholar]
- Chen, C.F. Studies on Vegetation Ecology between Chinshuiying and Chachayalaishan Areas of Southern Taiwan. Master’s Thesis, NPUST, Pingtung, Taiwan, 2006. [Google Scholar]
- Chian, Y.S.; Liao, C.F.; Yeh, C.L. Vegetation Analysis of Shihkochiehshan Area. Hwa Kang J Agric. 2010, 25, 15–40. [Google Scholar]
- Chian, Y.S.; Liu, H.Y.; Yeh, C.L.; Liao, C.F. Vegetation classification and mapping of Ailao river basin. Q. J. For. Res. 2010, 32, 1–14. [Google Scholar]
- Lo, K.P. Classification and Ordination of Plant Community on the Kutzulun River Basin of Eastern Taiwan. Master’s Thesis, NPUST, Pingtung, Taiwan, 2006. [Google Scholar]
- Wang, C.C.; Ou, C.H.; Lu, K.C.; Yeh, C.L.; Chiu, C.A.; Fan, K.C.; Chian, Y.S. The studies on flora and vegetation of Shuangkueihu wildlife important habitat. Q. J. Chin. For. 2014, 47, 27–36. [Google Scholar]
- Yeh, C.L.; Liao, J.H. Study on vegetation diversity of Peitawushan. J. Exp. For. NTU 2007, 21, 117–132. [Google Scholar]
- Yeh, C.L.; Liao, J.H. Vegetation ecology of Peitawushan in Pingtung County. Q. J. Chin. For. 2008, 41, 165–179. [Google Scholar]
- Yeh, C.L.; Liao, C.C. Vegetation mapping of the Peitawushan. Q. J. Chin. For. 2009, 42, 319–333. [Google Scholar]
- Yeh, Q.W.; Hsieh, S.I.; Chen, T.Y.; Yeh, C.L.; Lee, C.T. Study of vegetation diversity at Lilung mountains. Q. J. Chin. For. 2012, 45, 451–470. [Google Scholar]
- Li, C.F.; Zeleny, D.; Chytrý, M.; Chen, M.Y.; Chen, T.Y.; Chiou, C.R.; Hsia, Y.J.; Liu, H.Y.; Yang, S.Z.; Yeh, C.L.; et al. Chamaecyparis montane cloud forest in Taiwan: Ecology and vegetation classification. Ecol. Res. 2015, 30, 771–791. [Google Scholar] [CrossRef]
- Lin, C.T.; Li, C.F.; David, Z.; Chytrý, M.; Yukito, N.; Chen, M.Y.; Chen, T.Y.; Hsia, Y.J.; Hsieh, C.F.; Liu, H.Y.; et al. Classification of the high-mountain coniferous forests in Taiwan. Folia Geobot. 2012, 47, 373–401. [Google Scholar] [CrossRef]
- Hukusima, T.; Lu, S.Y.; Matsui, T.; Nishio, T.; Liu, C.L.; Pignatti, S. Phytosociology of Fagus hayatae forests in Taiwan. Rend. Lincei 2005, 16, 171–189. [Google Scholar] [CrossRef]
- Chytrý, M.; Schaminée, J.H.J.; Schwabe, A. Vegetation survey: A new focus for Applied Vegetation Science. Appl. Veg. Sci. 2011, 14, 435–439. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Hill, M.O. TWINSPAN—A FORTRAN Program for Arranging Multivariate Data in an Ordered Two-Way Table by Classification of the Individuals and Attributes; Cornell University: Ithaca, NY, USA, 1979. [Google Scholar]
- Bruelheide, H. Using formal logic to classify vegetation. Folia Geobot. 1997, 32, 41–46. [Google Scholar] [CrossRef]
- Bruelheide, H. A new measure of fidelity and its application to defining species groups. J. Veg. Sci. 2000, 11, 167–178. [Google Scholar] [CrossRef]
- Bruelheide, H.; Chytrý, M. Towards unification of national vegetation classifications: A comparison of two methods for analysis of large data sets. J. Veg. Sci. 2000, 11, 295–306. [Google Scholar] [CrossRef]
- Koci, M.; Chytrý, M.; Tichý, L. Formalized reproduction of an expert-based phytosociological classification: A case study of subalpine tall-forb vegetation. J. Veg. Sci. 2003, 14, 601–610. [Google Scholar] [CrossRef]
- Janišová, M.; Dúbravková, D. Formalized classification of rocky Pannonian grasslands and dealpine Sesleria-dominated grasslands in Slovakia using a hierarchical expert system. Phytocoenologia 2010, 40, 267–291. [Google Scholar] [CrossRef]
- Landucci, F.; Tichý, L.; Šumberová, K.; Chytrý, M. Formalized classification of species-poor vegetation: A proposal of a consistent protocol for aquatic vegetation. J. Veg. Sci. 2015, 26, 791–803. [Google Scholar] [CrossRef]
- Yang, W.F. Hydrological year book of Taiwan Republic of China; Water Resources Agency Ministry of Economic Affairs: Taipei, Taiwan, 2014. [Google Scholar]
- Forestry Bureau, Council of Agriculture, Executive Yuan. Available online: http://www.forest.gov.tw/0001493 (accessed on 5 July 2016).
- Su, H.-J. Studies on the climate and vegetation types of the natural forests in Taiwan (II): Altitudinal vegetation zones in relation to temperature gradient. Q. J. Chin. For. 1984, 17, 57–73. [Google Scholar]
- Chiou, C.R.; Hsieh, C.F.; Wang, J.C.; Chen, M.Y.; Liu, H.Y.; Yeh, C.L.; Yang, S.Z.; Chen, T.Y.; Hsia, Y.J.; Song, G.Z.M. The first national vegetation inventory in Taiwan. Taiwan J. For. Sci. 2009, 24, 295–302. [Google Scholar]
- Huang, T.C.; Hsieh, C.F. Flora of Taiwan, 2nd ed.; National Taiwan University: Taipei, Taiwan, 1994; Volume I–VI. [Google Scholar]
- WorldClim—Global Climate Data. Available online: http://www.worldclim.org (accessed on 5 October 2014).
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; Wiley & Sons: New York, NY, USA, 1974. [Google Scholar]
- Su, H.J.; Liou, C.Y. The Floristic-sociological Approach of Vegetation Classification. J. Exp. For. NTU 2004, 18, 129–151. [Google Scholar]
- Landucci, F.; Gigante, D.; Venanzoni, R.; Chytrý, M. Wetland vegetation of the class Phragmito-Magno-Caricetea in central Italy. Phytocoenologia 2013, 43, 67–100. [Google Scholar] [CrossRef]
- Kruskal, J.B. Nonmetric multidimensional scaling: A numerical method. Psychometrika 1964, 29, 115–129. [Google Scholar] [CrossRef]
- Tichý, L. JUICE, Software for vegetation classification. J. Veg. Sci. 2002, 13, 451–453. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2013. Available online: http://www.R-project.org (accessed on 30 April 2013).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.0-6. 2013. Available online: http://CRAN.R-project.org/package=vegan (accessed on 30 April 2013).
- Su, H.J. The diversification and synthesis of vegetation classification. J. Exp. For. NTU 2002, 16, 269–282. [Google Scholar]
- Su, H.J. A geographical data organization system for the botanical inventory of Taiwan. Institute of Botany, Academia Sinica. Monogr. Ser. 1992, 12, 23–36. [Google Scholar]
- Chiu, C.A.; Lin, P.H.; Lu, K.C. GIS-based tests for quality control of meteorological data and spatial interpolation of climate data. MRD 2009, 29, 339–349. [Google Scholar] [CrossRef]
- Chian, Y.S.; Liao, C.F.; Yeh, C.L.; Wang, C.C. The Study of Vegetation in Shihbaluohanshan Natural Reserve Area. Q. J. Chin. For. 2012, 45, 299–308. [Google Scholar]
- Wang, J.C.; Chiou, W.L.; Chang, H.M. A Preliminary Red List of Taiwanese Vascular Plants; Endemic Species Research Institute and Taiwan Society of Plant Systematics: Taipei, Taiwan, 2012. [Google Scholar]
- Chen, S.Y. Vegetation Ecology Along Varus River to Nantawushan in Pingtung County. Master’s Thesis, NPUST, Pingtung, Taiwan, 2014. [Google Scholar]
- Liu, C.P.; Yeh, C.L.; Chen, T.Y. Natural vegetation classification system of the Hengchun peninsula. J. Exp. For. NTU 2006, 20, 133–152. [Google Scholar]
- Chiu, C.A.; Wang, C.C.; Lu, K.C.; Lin, P.H.; Tzeng, H.Y. Examination of semi-arid regions and potential savanna vegetation in Taiwan. Taiwan J. For. Sci. 2008, 23, 23–36. [Google Scholar]
- Lin, C.J. Conferring the Composition and Distribution of Vegetation Diversity in Taiwan. Master’s Thesis, NTU, Taipei, Taiwan, 2009. [Google Scholar]
- Chiu, C.A.; Lin, P.H.; Tsai, C.Y. Spatio-temporal variation and monsoon effect on the temperature lapse rate of a subtropical island. TAO Sci. 2014, 20, 203–217. [Google Scholar] [CrossRef]
- Wang, J.C.; Huang, S.; Lue, K.Y.; Hsu, Y.F.; Chen, S.H. The Investigation on the Biota of Tawushan Nature Preserve—Jinlun Watershed; Taitung Forest District Office, Bureau of Forestry, Council of Agriculture Executive Yuan: Taipei, Taiwan, 2003. [Google Scholar]
- Sarah, J.I.; Regan, E.; Dov, F.S.; James, R. Niche expansion and temperature sensitivity of tropical African montane forests. Glob. Ecol. Biogeogr. 2016, 25, 693–703. [Google Scholar] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Axis 1 | Axis 2 | Axis 3 | |
|---|---|---|---|
| eigenvalue | 0.743 | 0.431 | 0.343 |
| length | 8.159 | 5.074 | 5.071 |
| Monte Carlo significance testing | |||
| p value | 0.002 ** | 0.062 | 0.086 |
| Variable | Axis 1 | Axis 2 | Axis 3 |
|---|---|---|---|
| Altitude | −0.915 ** | −0.109 | −0.112 |
| Canopy cover | −0.057 | −0.046 | −0.099 |
| Slope | 0.15 | −0.323 | −0.320 |
| Rock cover | 0.069 | −0.261 | 0.26 |
| Rockiness | 0.375 | −0.211 | −0.497 |
| WP | −0.415 ** | −0.814 ** | −0.006 |
| T1 | 0.402 ** | −0.023 | 0.172 |
| Topography | 0.343 | 0.368 ** | −0.338 |
| Scientific Name | Ecoclimatic Regions | |
|---|---|---|
| Southwestern | Southeastern | |
| Prunus phaeosticta | 38.2 | |
| Litsea akoensis | 34.8 | |
| Turpinia formosana | 32.6 | |
| Camellia sinensis f. formosana | 27.5 | |
| Ilex asprella | 25.2 | |
| Maesa perlaria v. formosana | 22.7 | |
| Pittosporum illicioides | 21.3 | |
| Polygonum chinense | 21.2 | |
| Wendlandia uvariifolia | 20.8 | |
| Schima superba v. kankaoensis | 43.2 | |
| Psychotria serpens | 41.9 | |
| Calamus quiquesetinervius | 41.4 | |
| Syzygium euphlebium | 37.1 | |
| Turpinia ternata | 36.8 | |
| Ventilago elegans | 36.4 | |
| Antidesma hiiranense | 35.5 | |
| Wendlandia formosana | 34.2 | |
| Ficus fistulosa | 32.9 | |
| Selaginella delicatula | 31.8 | |
| Ormosia hengchuniana | 31.5 | |
| Alpinia pricei | 30.8 | |
| Callicarpa remotiserrulata | 30.1 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chian, Y.-S.; Yeh, C.-L.; Wang, C.-C. Variation in Mountain Vegetation Composition between the East and the West Sides of Southern Taiwan. Forests 2016, 7, 179. https://doi.org/10.3390/f7080179
Chian Y-S, Yeh C-L, Wang C-C. Variation in Mountain Vegetation Composition between the East and the West Sides of Southern Taiwan. Forests. 2016; 7(8):179. https://doi.org/10.3390/f7080179
Chicago/Turabian StyleChian, Yi-Shin, Ching-Long Yeh, and Chih-Chiang Wang. 2016. "Variation in Mountain Vegetation Composition between the East and the West Sides of Southern Taiwan" Forests 7, no. 8: 179. https://doi.org/10.3390/f7080179
APA StyleChian, Y. -S., Yeh, C. -L., & Wang, C. -C. (2016). Variation in Mountain Vegetation Composition between the East and the West Sides of Southern Taiwan. Forests, 7(8), 179. https://doi.org/10.3390/f7080179