
Adaptive Variation and Introgression of a CONSTANS-Like Gene in North American Red Oaks


Abstract
:1. Introduction
2. Materials and Methods
3. Results
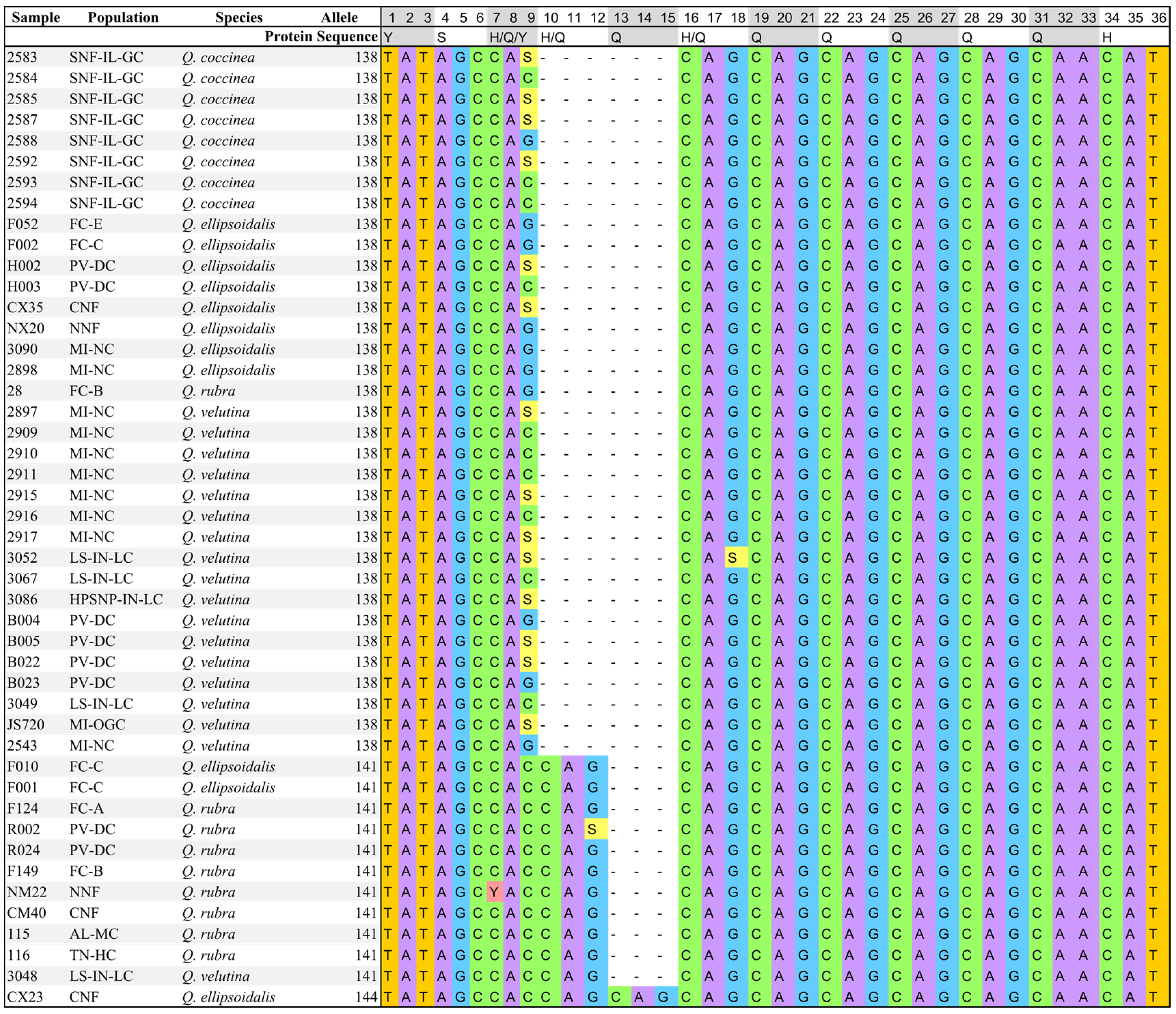
3.1. Nucleotide Sequence Analysis
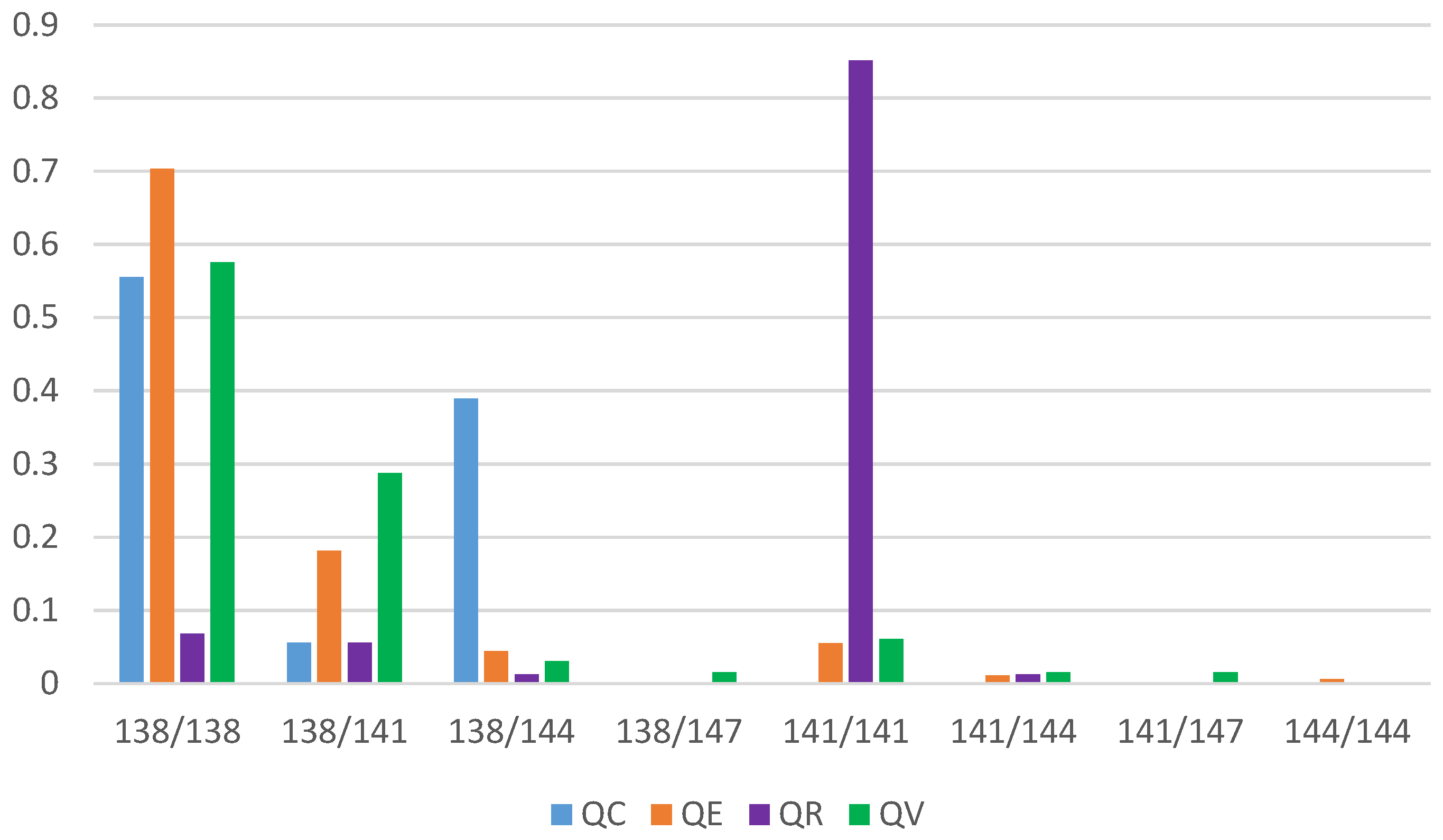
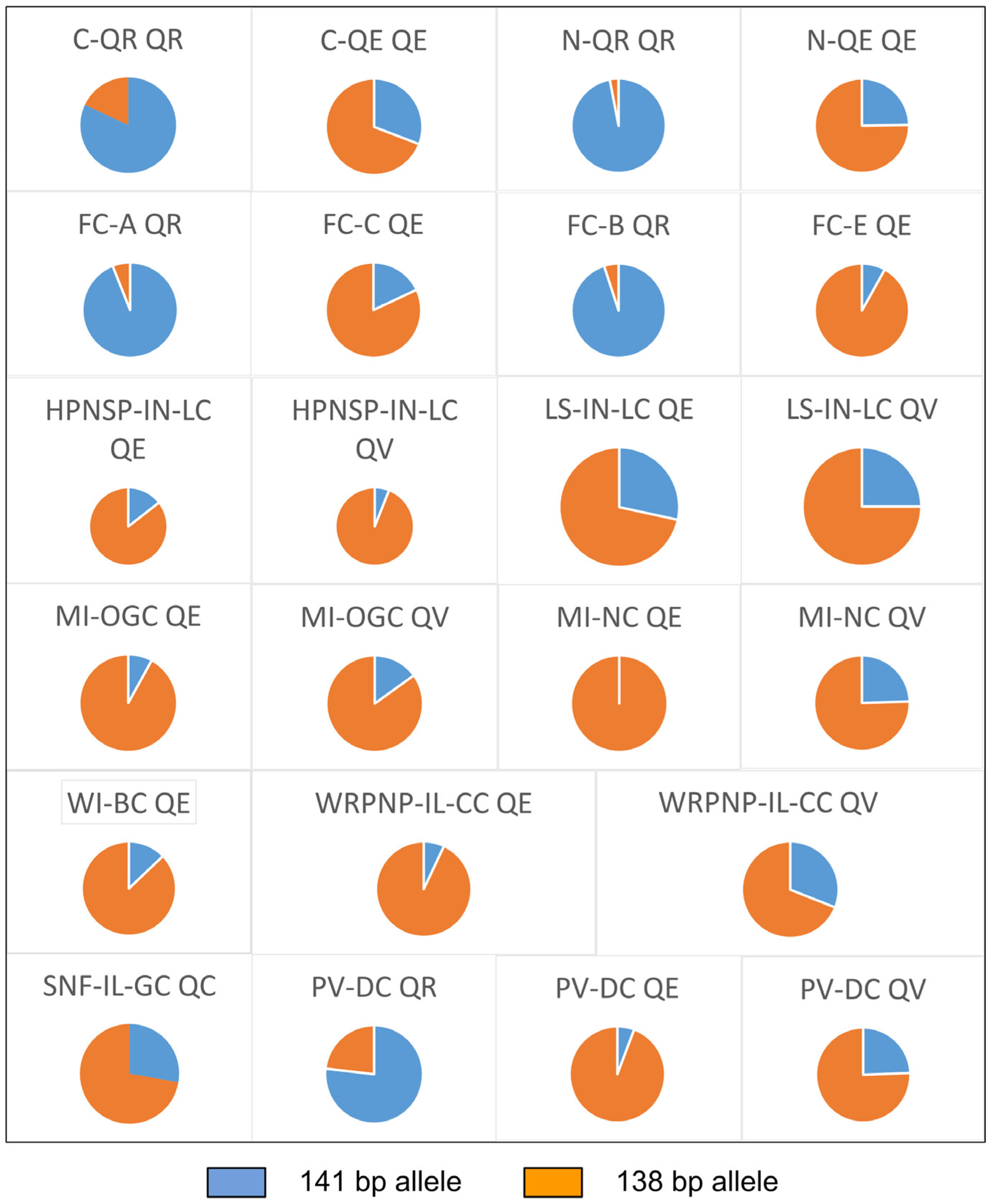
3.2. Genetic Diversity and Differentiation Analysis
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Arnold, M.L. Evolution through Genetic Exchange; Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Arnold, M.L. Divergence with Genetic Exchange; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Scotti-Saintagne, C.; Mariette, S.; Porth, I.; Goicoechea, P.G.; Barreneche, T.; Bodénès, C.; Burg, K.; Kremer, A. Genome scanning for interspecific differentiation between two closely related oak species (Quercus robur L. and Q. petraea (Matt.) Liebl.). Genetics 2004, 168, 1615–1626. [Google Scholar] [CrossRef] [PubMed]
- Gailing, O.; Curtu, A.L. Interspecific gene flow and maintenance of species integrity in oaks. Ann. For. Res. 2014, 57, 5–18. [Google Scholar] [CrossRef]
- Moran, E.V.; Willis, J.; Clark, J.S. Genetic evidence for hybridization in red oaks (Quercus sect. Lobatae, Fagaceae). Am. J. Bot. 2012, 99, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Lepais, O.; Gerber, S. Reproductive patterns shape introgression dynamics and species sucession within the European white oak species complex. Evolution 2011, 65, 156–170. [Google Scholar] [CrossRef] [PubMed]
- Lind-Riehl, J.F.; Sullivan, A.R.; Gailing, O. Evidence for selection on a CONSTANS-like gene between two red oak species. Ann. Bot. Lond. 2014, 113, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, A.R.; Owusu, S.A.; Weber, J.A.; Hipp, A.L.; Gailing, O. Hybridization and divergence in multi-species oak (Quercus) communities. Bot. J. Linn. Soc. 2016, 181, 99–114. [Google Scholar] [CrossRef]
- Hipp, A.L.; Weber, J.A. Taxonomy of Hill’s oak (Quercus ellipsoidalis: Fagaceae): Evidence from AFLP data. Syst. Bot. 2008, 33, 148–158. [Google Scholar] [CrossRef]
- Owusu, S.A.; Sullivan, A.R.; Weber, J.A.; Hipp, A.L.; Gailing, O. Taxonomic Relationships and Gene Flow in Four North American Quercus Species (Quercus section Lobatae). Syst. Bot. 2015, 40, 510–521. [Google Scholar] [CrossRef]
- Hipp, A.L.; Weber, J.A.; Srivastava, A. Who am I this time? The affinities and misbehaviors of Hill’s oak (Quercus ellipsoidalis). Int. Oak J. 2010, 21, 27–36. [Google Scholar]
- Hipp, A.L. Hill’s oak: The taxonomy and dynamics of a Western Great Lake endemic. Arnoldia 2010, 67, 2–14. [Google Scholar]
- Hipp, A.L. What do we mean by oak species and the oak phylogeny? Oak species coherence and parallel ecological diversification in the red and white oaks. In Presented at the IUFRO Genomics and forest tree genetics, Arcachon, France, 30 May–3 June 2016.
- Zhang, R.; Hipp, A.L.; Gailing, O. Sharing of chloroplast haplotypes among red oak species suggests interspecific gene flow between neighboring populations. Botany 2015, 93, 691–700. [Google Scholar] [CrossRef]
- Sullivan, A.R. Speciation with Gene-Flow in eastern North American Red Oaks (Quercus section Lobatae). Master’s Thesis, School of Forest Resources and Environmental Science, Houghton, MI, USA, 2013. [Google Scholar]
- Abrams, M.D. Adaptations and responses to drought in Quercus species of North America. Tree Physiol. 1990, 7, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.S.; Anderson, K.H.; Bartlein, P.J. Atlas of Relations between Climatic Parameters and Distributions of Important Trees and Shrubs in North America-Hardwoods; U.S.G. Survey: Denver, CO, USA, 1999.
- Durand, J.; Bodenes, C.; Chancerel, E.; Frigerio, J.-M.; Vendramin, G.; Sebastiani, F.; Buonamici, A.; Gailing, O.; Koelewijn, H.-P.; Villani, F.; et al. A fast and cost-effective approach to develop and map EST-SSR markers: Oak as a case study. BMC Genom. 2010, 11, 570. [Google Scholar] [CrossRef] [PubMed]
- Schmalenbach, I.; Zhang, L.; Reymond, M.; Jimenez-Gomez, J.M. The relationship between flowering time and growth responses to drought in the Arabidopsis Landsberg erecta x Antwerp-1 population. Front. Plant Sci. 2014, 5, 609. [Google Scholar] [CrossRef] [PubMed]
- Alberto, F.J.; Derory, J.; Boury, C.; Frigerio, J.M.; Zimmerman, N.E.; Kremer, A. Imprints of natural selection along environmental gradients in phenology-related genes of Quercus petraea. Genetics 2013, 195, 495–512. [Google Scholar] [CrossRef] [PubMed]
- Cavender-Bares, J.; Pahlich, A. Molecular, morphological, and ecological niche differentiation of sympatric sister oak species, Quercus virginiana and Q. geminata (Fagaceae). Am. J. Bot. 2009, 96, 1690–1702. [Google Scholar] [CrossRef] [PubMed]
- Ducousso, A.; Guyon, J.; Kremer, A. Latitudinal and altitudinal variation of bud burst in western populations of sessile oak (Quercus petraea (Matt) Liebl). Ann. Sci. For. 1996, 53, 775–782. [Google Scholar] [CrossRef]
- Gailing, O. Difference in growth, survival and phenology in Quercus rubra and Q. ellipsoidalis seedlings. Dendrobiology 2013, 70, 71–79. [Google Scholar] [CrossRef]
- Collins, E.; Sullivan, A.R.; Gailing, O. Limited effective gene flow between two interfertile red oak species. Trees 2015, 29, 1135–1148. [Google Scholar] [CrossRef]
- Lind, J.F.; Gailing, O. Genetic structure of Quercus rubra L. and Quercus ellipsoidalis E.J. Hill populations at gene-based EST-SSR and nuclear SSR markers. Tree Genet. Genomes 2013, 9, 707–722. [Google Scholar] [CrossRef]
- Ma, X.-F.; Hall, D.; Onge, K.R.S.; Jansson, S.; Ingvarsson, P.K. Genetic differentiation, clinal variation and phenotypic associations with growth cessation across the Populus tremula photoperiodic pathway. Genetics 2010, 186, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, K.G.; Banks, M.A. A latitudinal cline in the Chinook salmon (Oncorhynchus tshawytscha) Clock gene: Evidence for selection on PolyQ length variants. Proc. R. Soc. B Biol. Sci. 2008, 275, 2813–2821. [Google Scholar] [CrossRef] [PubMed]
- Weih, M. Genetic and environmental variation in spring and autumn phenology of biomass willows (Salix spp.): Effects on shoot growth and nitrogen economy. Tree Physiol. 2009, 29, 1479–1490. [Google Scholar] [CrossRef] [PubMed]
- Faux, N.G.; Bottomley, S.P.; Lesk, A.M.; Irving, J.A.; Morrison, J.R.; Garcia de la Banda, M.; Whisstock, J.C. Functional insights from the distribution and role of homopeptide repeat-containing proteins. Genome Res. 2005, 15, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Lobanov, M.Y.; Klus, P.; Sokolovsky, I.V.; Tartaglia, G.G.; Galzitskaya, O.V. Non-random distribution of homo-repeats: Links with biological functions and human disease. Sci. Rep. 2016, 6, 26941. [Google Scholar] [CrossRef] [PubMed]
- Haerty, W.; Golding, G.B. Low-complexity sequences and single amino acid repeats: Not just “junk” peptide sequences. Genome 2010, 53, 753–762. [Google Scholar] [PubMed]
- Hall, T.A. Bioedit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.O.D.; Smouse, P.E. GeneAlEx 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GenePop (Version-1.2)—Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar]
- Antao, T.; Lopes, A.; Lopes, R.; Beja-Pereira, A.; Luikart, G. LOSITAN: A workbench to detect molecular adaptation based on a Fst-outlier method. BMC Bioinform. 2008, 9, 323. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Barton, A.M.; Gleeson, S.K. Ecophysiology of seedlings of oaks and red maple across a topographic gradient in eastern Kentucky. For. Sci. 1996, 42, 335–342. [Google Scholar]
- Gailing, O.; Lind, J.F.; Lilleskov, E. Leaf morphological and genetic differentiation between Quercus rubra L. and Q. ellipsoidalis E.J. Hill populations in contrasting environments. Plant Syst. Evol. 2012, 298, 1533–1545. [Google Scholar] [CrossRef]
- Van Valen, L. Ecological species, multispecies, and oaks. Taxon 1976, 25, 233–239. [Google Scholar]
- Minder, A.M.; Widmer, A. A population genomic analysis of species boundaries: Neutral processes, adaptive divergence and introgression between two hybridizing plant species. Mol. Ecol. 2008, 17, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.D.; Mekel-Bobrov, N.; Vallender, E.J.; Hudson, R.R.; Lahn, B.T. Evidence that the adaptive allele of the brain size gene microcephalin introgressed into Homo sapiens from an archaic Homo lineage. Proc. Natl. Acad. Sci. USA 2006, 103, 18178–18183. [Google Scholar] [CrossRef] [PubMed]
- Whitney, K.; Randell, R.A.; Rieseberg, L.H. Adaptive introgression of herbivore resistance traits in the weedy sunflower Helianthus annuus. Am. Nat. 2006, 167, 794–807. [Google Scholar] [CrossRef] [PubMed]
- Motzkin, G.; Ciccarello, S.C.; Foster, D.R. Frost pockets on a level sand plain: Does variation in microclimate help maintain persistent vegetation patterns? J. Torrey Bot. Soc. 2002, 129, 154–163. [Google Scholar] [CrossRef]
- Abrams, M.D. Comparative water relations of three successional hardwood species in central Wisconsin. Tree Physiol. 1988, 4, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Abrams, M. Distribution, historical development and ecophysiological attributes of oak species in the eastern United States. Ann. Sci. For. 1996, 53, 487–512. [Google Scholar] [CrossRef]
- Jensen, R.J. Quercus Linnaeus sect. Lobatae Loudon, the red or black oaks. In Flora of North America North of Mexico; Flora of North America Editoral Committee; Oxford University Press on Demand: Oxford, UK, 1997; Volume 3, pp. 447–468. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Q. ellipsoidalis | Q. rubra | Q. velutina | |
|---|---|---|---|
| Q. ellipsoidalis | - | 0.043 *** | 0.030 *** |
| Q. rubra | 0.672 *** | - | 0.032 *** |
| Q. velutina | 0.009 * | 0.625 *** | - |
| Population*** | Species | Population Type | Sample Size (N) | Allele 141 * | Allele 138 * |
|---|---|---|---|---|---|
| C-QE | QE | parapatric | 31 | 0.29 | 0.65 |
| C-QR | QR | parapatric | 31 | 0.82 | 0.18 |
| N-QE | QE | parapatric | 31 | 0.24 | 0.73 |
| N-QR | QR | parapatric | 32 | 0.98 | 0.02 |
| FC-A | QR | parapatric | 36 | 0.94 | 0.06 |
| FC-C | QE | parapatric | 40 | 0.18 | 0.82 |
| FC-B | QR | parapatric | 36 | 0.95 | 0.05 |
| FC-E | QE | parapatric | 37 | 0.08 | 0.91 |
| HPNSP-IN-LC | QE | parapatric | 15 | 0.13 | 0.77 |
| QV | 8 | 0.06 | 0.94 | ||
| LS-IN-LC | QE | parapatric | 4 | 0.25 | 0.63 |
| QV | 4 | 0.25 | 0.75 | ||
| MI-NC | QE | parapatric | 8 | 0 | 0.93 |
| QV | 17 | 0.24 | 0.74 | ||
| MI-OGC | QE | parapatric | 6 | 0.08 | 0.92 |
| QV | 7 | 0.14 | 0.79 | ||
| WI-BC | QE | parapatric | 8 | 0.13 | 0.88 |
| WRPNP-IL-CC ** | QE | parapatric | 7 | 0.07 | 0.93 |
| QV | 8 | 0.31 | 0.69 | ||
| QR | 1 | 0 | 1.00 | ||
| SNF-IL-GC | QC | parapatric | 20 | 0.03 | 0.78 |
| PV-DC | QE | sympatric | 11 | 0.05 | 0.82 |
| QV | 17 | 0.21 | 0.65 | ||
| QR | 16 | 0.63 | 0.19 |
| Q. rubra with Introgression from Q. ellipsoidalis or Q. velutina | Q. ellipsoidalis/Q. velutina with Introgression from Q. rubra | F1 Hybrids of Q. rubra with Q. ellipsoidalis or Q. velutina | Total Introgressive Forms and F1 Hybrids between Q. rubra and Q. ellipsoidalis or Q. velutina | |
|---|---|---|---|---|
| Parapatric populations of Q. ellipsoidalis and Q. rubra** | ||||
| C-QE | - | 0.20 | 0.03 | 0.23 |
| C-QR | 0.23 | - | - | 0.23 |
| N-QE | - | 0.18 | 0.03 | 0.21 |
| N-QR | 0.20 | - | - | 0.20 |
| FC-A | 0.13 | - | - | 0.13 |
| FC-C | - | 0.10 | 0 | 0.10 |
| FC-B | 0.00 | - | - | 0.00 |
| FC-E | - | 0.08 | 0 | 0.08 |
| Sympatric populations of Q. ellipsoidalis and Q. velutina in parapatry with Q. rubra** | ||||
| HPNSP-IN-LC | - | 0.04 | 0.00 | 0.04 |
| LS-IN-LC | - | 0.00 | 0.00 | 0 |
| MI-NC | - | 0.03 | 0.00 | 0.03 |
| MI-OGC | - | 0.18 | 0.03 | 0.21 |
| WI-BC | - | 0.04 | 0.04 | 0.08 |
| WRPNP-IL-CC | - | 0.00 | 0.00 | 0.00 |
| Sympatric populations of Q. ellipsoidalis, Q. velutina and Q. rubra** | ||||
| PV-DC | 0.24 | 0.11 | 0.10 | 0.45 |
| Q. coccinea population** | ||||
| SNF-IL-GC | - | 0.00 | 0.00 | 0.00 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lind-Riehl, J.F.; Gailing, O. Adaptive Variation and Introgression of a CONSTANS-Like Gene in North American Red Oaks. Forests 2017, 8, 3. https://doi.org/10.3390/f8010003
Lind-Riehl JF, Gailing O. Adaptive Variation and Introgression of a CONSTANS-Like Gene in North American Red Oaks. Forests. 2017; 8(1):3. https://doi.org/10.3390/f8010003
Chicago/Turabian StyleLind-Riehl, Jennifer F., and Oliver Gailing. 2017. "Adaptive Variation and Introgression of a CONSTANS-Like Gene in North American Red Oaks" Forests 8, no. 1: 3. https://doi.org/10.3390/f8010003
APA StyleLind-Riehl, J. F., & Gailing, O. (2017). Adaptive Variation and Introgression of a CONSTANS-Like Gene in North American Red Oaks. Forests, 8(1), 3. https://doi.org/10.3390/f8010003