Climate Impacts on Tree Growth in the Sierra Nevada


Abstract
:1. Introduction
2. Material and Methods
2.1. Data
2.2. Simulated Dataset
2.2.1. Testing the Effect of Different Observation Time Intervals
2.2.2. Testing the Ability to Detect Subtle Climate Effects
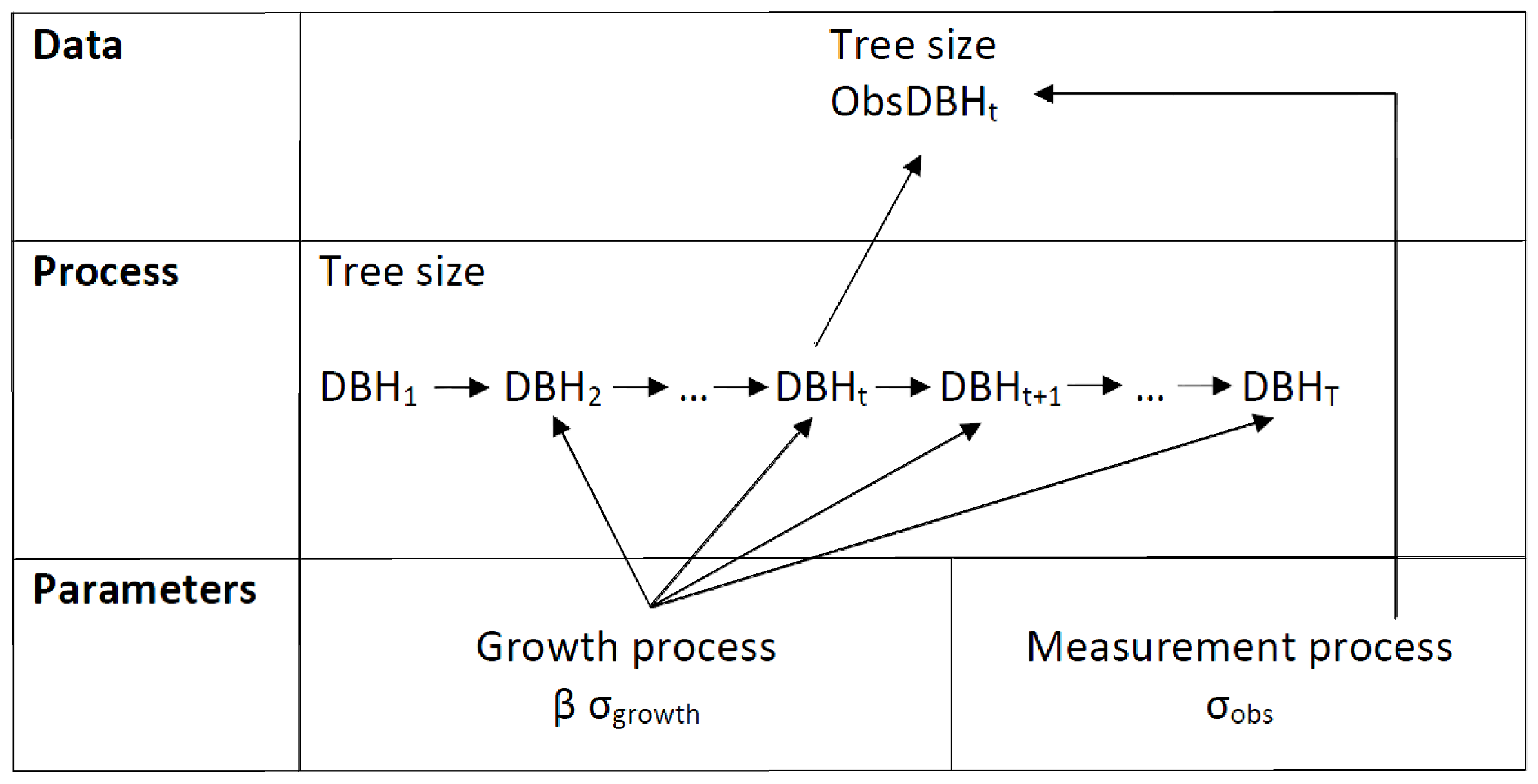
2.3. Model
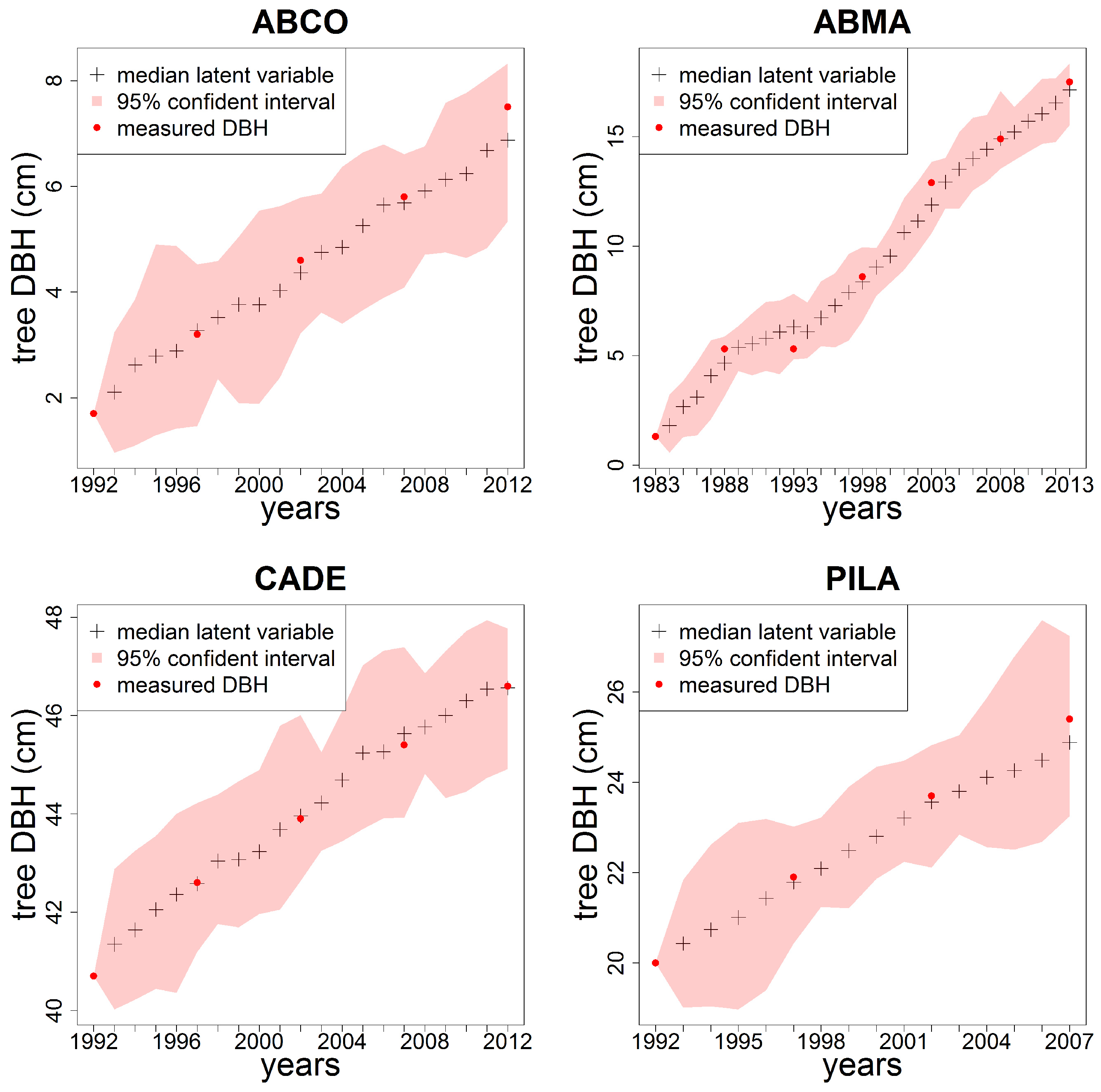
3. Results
4. Discussion
4.1. The Growth Model
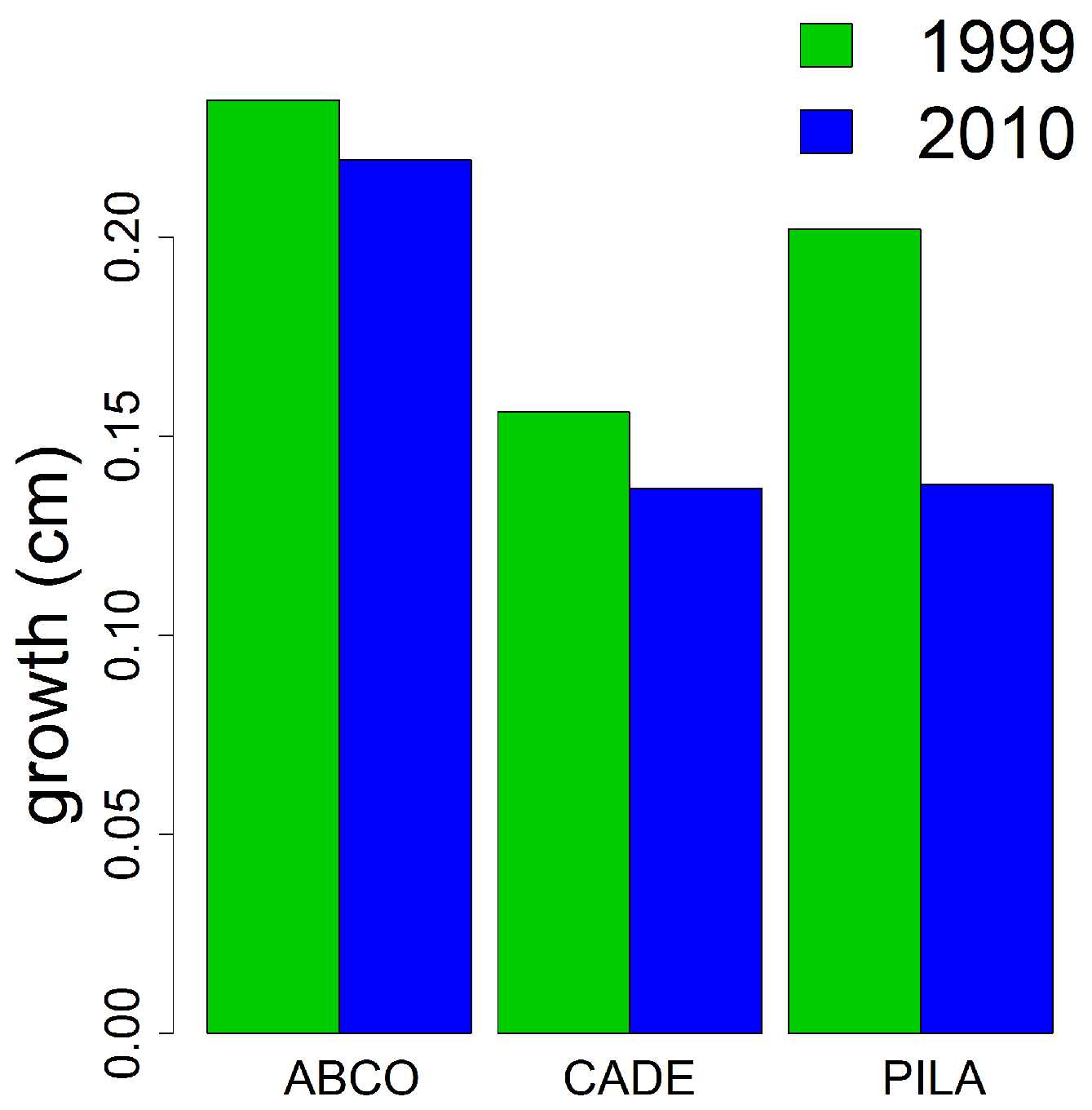
4.2. Growth Response to Climate
4.3. Consequences in the Sierra Nevada
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
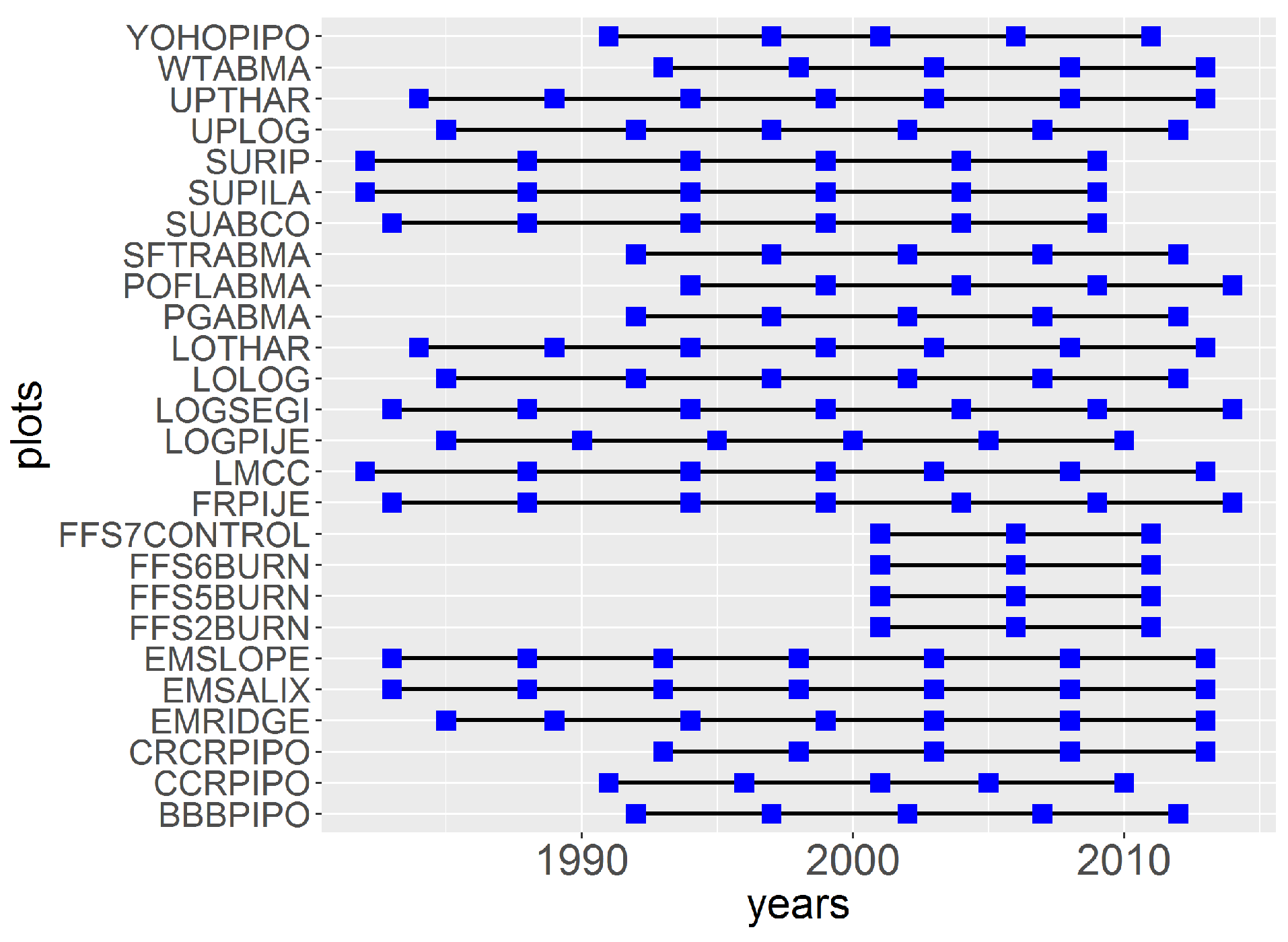
Appendix A. Years of DBH Measurement for the Different Plots

Appendix B. DIC Used for the Model Selection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No Species-Specific Term | Species-Specific Term for Size Effect | Species-Specific Term for Competition Effect |
|---|---|---|
| 267598 | 273867 | 273907 |
| AvTemp | JanMin | JulMax | precip | snow | CWD | AET | ASpck |
| 273615 | 273014 | 273535 | 273149 | 272191 | 272984 | 272827 | 274591 |
| AvTemp | JanMin | JulMax | precip | snow | CWD | AET | ASpck |
| 271408 | 275333 | 273214 | 274070 | 273626 | 272997 | 274212 | 273046 |
| AvTemp | JanMin | JulMax | precip | snow | CWD | AET | ASpck |
| 272991 | 272411 | 273620 | 273252 | 273008 | 271669 | 273464 | 274833 |
| AvTemp | JanMin | JulMax | precip | snow | CWD | AET | ASpck |
| 274132 | 273904 | 274560 | 273662 | 272365 | 272829 | 272870 | 272255 |
Appendix C. Results Table
| PIMO | PICO | ABMA | PIJE | ABCO | |
| AvTemp | 0.0158 | −0.0555 | 0.0131 | 0.0455 | 0.00347 |
| [−0.0115; 0.0497] | [−0.0942; −0.0189] | [0.00606; 0.0183] | [0.0187; 0.0792] | [0.000897; 0.00681] | |
| JanMin | 0.0124 | −0.0812 | 0.0023 | 0.0151 | 0.00339 |
| [−0.0125; 0.0433] | [−0.125; −0.0271] | [−0.00324; 0.00884] | [0.00701; 0.0287] | [0.00113; 0.00535] | |
| JulMax | −0.00331 | −0.0317 | 0.0096 | −0.00594 | 0.000864 |
| [−0.0169; 0.0139] | [−0.0507; −0.00941] | [0.00552; 0.0129] | [−0.0204; 0.00741] | [−0.00133; 0.00375] | |
| precip | 0.000159 | −0.000119 | −2.6e−05 | 8.43e−05 | 5.72e−05 |
| [−8.62e−05; 0.000258] | [−0.000452; 0.000114] | [−8.94e−05; 5.75e−06] | [−6.69e−05; 0.000216] | [−0.000104; 8.86e−05] | |
| snow | 0.000136 | 7.17e−05 | −3.34e−05 | 1.45e−05 | −1.15e−05 |
| [−2.6e−05; 0.000255] | [−0.000165; 0.000402] | [−0.000119; −4.76e−06] | [−0.000219; 0.000158] | [−6.22e−05; 1.79e−05] | |
| CWD | 0.000648 | −0.000732 | 0.000162 | 0.000809 | −1.48e−05 |
| [0.000125; 0.00111] | [−0.0015; −5.2e−05] | [7.11e−05; 0.00027] | [0.000388; 0.00124] | [−7.05e−05; 0.000101] | |
| AET | −0.00104 | −0.00111 | 0.000291 | 0.000165 | 9.25e−05 |
| [−0.00166; −0.000317] | [−0.00265; 0.000617] | [0.000159; 0.000411] | [−0.000298; 0.000618] | [−0.000136; 0.000216] | |
| ASpck | 0.000139 | 0.000179 | −5.28e−05 | −2.86e−05 | 1.99e−05 |
| [−1.64e−05; 0.00027] | [−5.3e−06; 0.000416] | [−8.56e−05; −2.03e−05] | [−0.00015; 8.55e−05] | [−3.25e−05; 5.01e−05] | |
| QUKE | QUCH | PILA | CADE | PIPO | |
| AvTemp | 0.029 | −0.0675 | −0.00264 | −0.00391 | −0.0496 |
| [0.0206; 0.0373] | [−0.191; −9.63e−05] | [−0.00723; 0.00343] | [−0.00805; 0.000868] | [−0.0789; −0.0249] | |
| JanMin | 0.0171 | −0.0683 | −0.00102 | −0.00524 | −0.0211 |
| [0.00999; 0.0236] | [−0.122; −0.0127] | [−0.0047; 0.00364] | [−0.00779; −0.00296] | [−0.0332; −0.0104] | |
| JulMax | 0.0309 | −0.0245 | −0.00185 | 0.000691 | 0.0224 |
| [0.0234; 0.0383] | [−0.057; 0.0289] | [−0.00632; 0.00324] | [−0.00211; 0.00485] | [0.00389; 0.0401] | |
| precip | −0.000147 | 0.000489 | −4.25e−05 | 7.07e−05 | 8.25e−05 |
| [−0.000279; −1.86e−05] | [−0.000273; 0.00174] | [−0.000149; 2.55e−05] | [3.22e−05; 0.000121] | [−5.03e−05; 0.000211] | |
| snow | −0.00029 | 0.000445 | 1.31e−05 | 8.25e−05 | 0.000375 |
| [−0.000378; −0.00021] | [−0.000493; 0.00142] | [−3.23e−05; 6.62e−05] | [2.69e−05; 0.00012] | [0.000125; 0.000523] | |
| CWD | 0.000688 | 0.000482 | 6.68e−06 | −0.000106 | −0.000107 |
| [0.000486; 0.000908] | [−0.00109; 0.0017] | [−0.000147; 0.000125] | [−0.000194; −8.26e−06] | [−0.000648; 0.000246] | |
| AET | 0.000573 | −0.0035 | −6.87e−05 | −8.05e−05 | −0.000664 |
| [0.000299; 0.000813] | [−0.00587; −0.000261] | [−0.000396; 0.000224] | [−0.000262; 0.000122] | [−0.00134; 0.000367] | |
| ASpck | −0.000232 | 7e−04 | 6.42e−05 | 4e−05 | 0.000165 |
| [−0.000301; −0.000184] | [−0.000601; 0.00172] | [2.47e−06; 0.00013] | [−1.57e−05; 8.83e−05] | [−0.000484; 0.00073] | |
| PIMO | PICO | ABMA | PIJE | ABCO | |
| AvTemp | 0.167 | −0.0746 | −0.04 | 0.111 | −0.0243 |
| [0.0945; 0.251] | [−0.203; 0.04] | [−0.0694; 0.00358] | [0.0507; 0.183] | [−0.039; 0.0491] | |
| JanMin | 0.0253 | −0.119 | −0.0248 | 0.0552 | 0.0154 |
| [−0.0166; 0.0619] | [−0.276; −0.00465] | [−0.0335; −0.0151] | [0.0185; 0.104] | [0.00463; 0.0371] | |
| JulMax | −0.000347 | −0.000227 | 0.000272 | 0.0164 | −0.0114 |
| [−0.0301; 0.0181] | [−0.0384; 0.0584] | [−0.00631; 0.00923] | [−0.0155; 0.038] | [−0.0168; 0.00069] | |
| precip | 0.000116 | −0.000137 | 1.79e−05 | 5.27e−05 | 7.22e−05 |
| [−0.00011; 0.000239] | [−0.000388; 3.6e−05] | [−6.48e−05; 6.13e−05] | [−8.55e−05; 0.000243] | [−4.92e−05; 9.68e−05] | |
| snow | 0.000153 | -0.000204 | 3.76e−05 | 5.03e−05 | 0.000117 |
| [−4.56e−05; 0.000266] | [−0.000528; 3.72e−05] | [2.78e−06; 6.93e−05] | [−0.000171; 0.000225] | [−0.000129; 0.000164] | |
| CWD | 0.000756 | 7.63e−06 | −6.78e−05 | 0.00107 | −0.000314 |
| [0.000247; 0.00149] | [−0.000949; 0.0011] | [−0.000218; 0.000178] | [0.00048; 0.00173] | [−4e−04; 0.000153] | |
| AET | −0.00084 | −0.000601 | 4.53e−05 | −0.000955 | 0.000366 |
| [−0.00215; 0.000122] | [−0.00263; 0.00125] | [−0.000353; 0.00026] | [−0.00221; 0.00043] | [−6.78e−05; 0.000529] | |
| ASpck | 0.000163 | −0.000172 | −5.59e−06 | 1.48e−05 | 0.000136 |
| [2.88e−05; 0.000288] | [−0.000408; 2.26e−05] | [−0.000115; 3.44e−05] | [−0.000146; 0.000153] | [2.55e−05; 0.000172] | |
| QUKE | QUCH | PILA | CADE | PIPO | |
| AvTemp | −0.0172 | −0.0614 | −0.0136 | −0.0602 | −0.0821 |
| [−0.0856; 0.0382] | [−0.268; 0.181] | [−0.0505; 0.0287] | [−0.0928; −0.0313] | [−0.173; -0.00601] | |
| JanMin | −0.00503 | −0.0969 | 0.00421 | −0.00732 | 0.00965 |
| [−0.0321; 0.0147] | [−0.151; −0.0212] | [−0.0157; 0.024] | [−0.0204; 0.00962] | [−0.0377; 0.0672] | |
| JulMax | 0.00679 | 0.196 | 0.00109 | −0.0229 | −0.0179 |
| [−0.0167; 0.0246] | [0.0226; 0.304] | [−0.00792; 0.0118] | [−0.0326; −0.0131] | [−0.0505; 0.00794] | |
| precip | 0.000121 | 0.000138 | 5.75e−06 | 9e−05 | 0.000154 |
| [−1.12e−05; 0.000242] | [−0.000954; 0.000908] | [−7.87e−05; 7.28e−05] | [−1.41e−05; 0.000151] | [8.18e−06; 0.000299] | |
| snow | 0.000176 | 7.32e−05 | −6.23e−07 | 0.000194 | 0.00046 |
| [4.96e−06; 0.000359] | [−0.00161; 0.00155] | [−0.000124; 8.97e−05] | [0.000109; 0.000309] | [3.22e−06; 0.000888] | |
| CWD | −0.000641 | 0.00263 | −0.000163 | −0.000359 | 1.65e−05 |
| [−0.00114; −0.000298] | [0.000957; 0.00414] | [−0.000396; 5.67e−05] | [−0.000538; 1.43e−05] | [−0.00062; 0.000558] | |
| AET | 0.00094 | −0.00372 | 9.91e−05 | 0.000385 | −0.000124 |
| [0.000511; 0.00166] | [−0.00538; −0.000731] | [−0.000321; 0.000502] | [−5.43e−06; 0.00055] | [−0.000849; 0.000662] | |
| ASpck | 0.000154 | −0.00164 | 9.45e−05 | 0.000127 | −0.000313 |
| [−2.76e−05; 0.000437] | [−0.00385; 0.000829] | [−1.81e−05; 0.000201] | [−7.33e−05; 0.000264] | [−0.00176; 0.000858] |
| PIMO | PICO | ABMA | PIJE | ABCO | |
| AvTemp | 0.0367 | −0.0549 | 0.0137 | 0.033 | 0.00385 |
| [0.0111; 0.0756] | [−0.094; −0.0215] | [0.00767; 0.018] | [0.0116; 0.0587] | [0.00147; 0.00707] | |
| JanMin | 0.0241 | −0.0801 | 0.0068 | 0.0148 | 0.00312 |
| [−0.00401; 0.0469] | [−0.138; −0.0293] | [0.000873; 0.0123] | [0.00582; 0.0271] | [0.00121; 0.00475] | |
| JulMax | −0.00629 | −0.0296 | 0.00927 | −0.00445 | 0.00123 |
| [−0.0269; 0.0132] | [−0.0457; −0.00719] | [0.00675; 0.0125] | [−0.0175; 0.00789] | [−0.000615; 0.004] | |
| precip | −1.94e−05 | −0.00012 | −2.48e−05 | −6.46e−05 | 6.05e−05 |
| [−0.000177; 0.000124] | [−0.000337; 0.00016] | [−5.97e−05; 1.16e−05] | [−0.000212; 7.31e−05] | [3.55e−05; 8.05e−05] | |
| snow | −3.93e−05 | 0.000157 | −6.1e−05 | −0.000105 | −3.62e−06 |
| [−0.000117; 9.55e−05] | [−0.000182; 0.000463] | [−0.000126; −2.38e−06] | [−0.000226; 1.44e−05] | [−2.3e−05; 1.88e−05] | |
| CWD | 0.00133 | −0.000969 | 0.00019 | 0.000567 | 1.69e−05 |
| [0.000822; 0.00166] | [−0.00159; −0.000382] | [4.66e−05; 0.000283] | [0.000192; 0.000847] | [−3.44e−05; 7.89e−05] | |
| AET | −0.00169 | 0.00043 | 0.000301 | −3.93e−05 | 7.88e−05 |
| [−0.00255; −0.000792] | [−0.00163; 0.00247] | [0.000143; 0.000423] | [−0.000504; 0.000517] | [−2.74e−05; 0.000192] | |
| ASpck | 1.46e−05 | 0.000172 | −5.87e−05 | −0.000101 | 1.34e−05 |
| [−9.16e−05; 0.000135] | [−6.22e−06; 0.000358] | [−9.51e−05; −3.48e−05] | [−0.000197; 1.47e−05] | [−7.91e−06; 3.79e−05] | |
| QUKE | QUCH | PILA | CADE | PIPO | |
| AvTemp | 0.0294 | −0.0405 | −0.0021 | −0.0034 | −0.0545 |
| [0.0226; 0.0388] | [−0.113; 0.0463] | [−0.00931; 0.00352] | [−0.00759; −1.88e−05] | [−0.0812; −0.0224] | |
| JanMin | 0.0189 | −0.016 | −0.000411 | −0.00475 | −0.0222 |
| [0.0135; 0.0243] | [−0.0973; 0.0596] | [−0.00395; 0.00288] | [−0.00788; −0.00166] | [−0.0348; −0.0114] | |
| JulMax | 0.0298 | 0.000459 | −0.00118 | 0.000997 | 0.0215 |
| [0.0199; 0.0371] | [−0.0492; 0.048] | [−0.00679; 0.00468] | [−0.0031; 0.00427] | [0.00271; 0.0382] | |
| precip | −0.000126 | 0.000623 | 2.21e−06 | 5.87e−05 | 7.55e−05 |
| [−0.00029; −1.06e−05] | [−0.000143; 0.00151] | [−5.16e−05; 5.01e−05] | [1.47e−05; 0.000106] | [−7.39e−05; 0.00023] | |
| snow | −0.00028 | 0.000491 | 2.17e−05 | 6.75e−05 | 0.000347 |
| [−0.000371; −0.000193] | [−0.000239; 0.00127] | [−2.7e−05; 7.21e−05] | [3.53e−05; 0.000112] | [0.000188; 0.000543] | |
| CWD | 0.000605 | −0.000128 | −3.07e−06 | −4.21e−05 | −0.00015 |
| [0.000435; 0.000803] | [−0.00109; 0.00127] | [−0.000104; 0.000109] | [−0.000141; 2.17e−05] | [−0.000563; 0.000316] | |
| AET | 0.000634 | −0.00233 | −4.69e−05 | −0.000158 | −0.000785 |
| [0.000394; 0.000901] | [−0.0045; 0.000135] | [−0.000238; 0.000207] | [−0.000372; 9.58e−06] | [−0.00158; −0.00018] | |
| ASpck | −0.000236 | 0.00063 | 5.6e−05 | 1.64e−05 | 8.69e−05 |
| [−0.000319; −0.000142] | [−0.00028; 0.00145] | [6.41e−06; 0.000106] | [−2.27e−05; 6.27e−05] | [−0.000615; 0.00109] | |
| PIMO | PICO | ABMA | PIJE | ABCO | |
| AvTemp | 0.266 | −0.0923 | −0.0532 | 0.0809 | −0.017 |
| [0.193; 0.341] | [−0.264; 0.0499] | [−0.0731; −0.0164] | [0.0217; 0.161] | [−0.0317; 0.0242] | |
| JanMin | 0.0405 | 0.00663 | −0.0325 | 0.0617 | 0.00547 |
| [−0.00321; 0.0784] | [−0.136; 0.192] | [−0.0448; −0.0173] | [0.0102; 0.1] | [−0.00148; 0.0126] | |
| JulMax | −0.024 | −0.0238 | 0.00043 | 0.0254 | −0.015 |
| [−0.0587; 0.0428] | [−0.0723; 0.0595] | [−0.00728; 0.0109] | [−0.000453; 0.054] | [−0.0203; −0.00276] | |
| precip | −1.79e−05 | −0.000119 | 5.72e−05 | −6.8e−05 | 8.15e−05 |
| [−0.000143; 8.15e−05] | [−0.000358; 0.000168] | [1.09e−05; 1e−04] | [−0.000221; 4.93e−05] | [5.7e−05; 0.000108] | |
| snow | −9.72e−06 | −0.000121 | 6.95e−05 | −0.000101 | 0.000114 |
| [−0.000104; 9.86e−05] | [−0.000415; 0.000161] | [1.56e−05; 0.000137] | [−0.000297; 8.04e−05] | [8.03e−05; 0.000154] | |
| CWD | 0.00174 | −0.000487 | −0.000141 | 0.000746 | −0.000194 |
| [0.00119; 0.00247] | [−0.00124; 0.000273] | [−0.00031; −1.09e−06] | [0.000246; 0.0012] | [−0.000286; −0.000102] | |
| AET | −0.00197 | 0.000892 | 0.000118 | −0.00132 | 0.000277 |
| [−0.00276; −0.00129] | [−0.000944; 0.00239] | [−7.62e−05; 0.000362] | [−0.00266; −2e−04] | [0.000136; 0.000389] | |
| ASpck | −1.2e−06 | −0.000136 | 2.24e−05 | −7.82e−05 | 0.000121 |
| [−0.000121; 0.000136] | [−0.000333; 0.000132] | [−4.14e−05; 7.31e−05] | [−0.00022; 7.38e−05] | [6.93e−05; 0.000252] | |
| QUKE | QUCH | PILA | CADE | PIPO | |
| AvTemp | 0.0193 | 0.197 | −0.000308 | −0.0402 | −0.0354 |
| [−0.0282; 0.0848] | [−0.0303; 0.489] | [−0.0382; 0.0322] | [−0.0664; −0.00766] | [−0.166; 0.0382] | |
| JanMin | 0.0524 | −0.184 | 0.00383 | 0.00739 | 0.0454 |
| [0.0233; 0.0852] | [−0.353; 0.247] | [−0.0125; 0.0245] | [−0.0109; 0.024] | [−0.0127; 0.0985] | |
| JulMax | −0.00655 | 0.168 | 0.007 | −0.0177 | −0.0157 |
| [−0.0218; 0.0111] | [0.059; 0.263] | [−0.00472; 0.0209] | [−0.0263; −0.0101] | [−0.0354; 0.00754] | |
| precip | 0.000117 | 0.000448 | 4.78e−05 | 4.26e−05 | 4.89e−05 |
| [−3.78e−05; 0.000247] | [−0.000289; 0.000972] | [−3.09e−05; 0.00012] | [−1.35e−05; 0.000104] | [−0.000192; 0.000266] | |
| snow | 8.85e−05 | 0.000334 | 7.46e−05 | 0.000148 | 0.000168 |
| [−7.96e−05; 0.000318] | [−0.000861; 0.00172] | [−4.73e−05; 0.000154] | [7.42e−05; 0.000248] | [−0.000232; 0.000579] | |
| CWD | −0.000581 | 0.00155 | −0.000164 | −0.000141 | 0.000234 |
| [−0.00095; −0.000149] | [−3.18e−05; 0.00385] | [−0.000331; 8.21e−05] | [−0.000278; 2.59e−05] | [−0.000516; 0.000881] | |
| AET | 0.000902 | −0.00294 | 0.000229 | 0.000153 | −0.000225 |
| [0.000499; 0.00136] | [−0.00631; 0.000564] | [−0.000253; 0.000529] | [−0.000148; 0.000422] | [−0.00113; 0.000736] | |
| ASpck | 0.000133 | −0.000427 | 0.000113 | 8.14e−05 | −0.000386 |
| [−7.98e−05; 0.000343] | [−0.00217; 0.00161] | [2.16e−05; 0.000234] | [−3.51e−06; 0.000164] | [−0.00147; 0.000704] |
Appendix D. Final Model
| DBH | Competition | Growth | Measurement | ||
| Error | Error | ||||
| 0.00118 | −0.00119 | 0.752 | 0.695 | ||
| [0.00111; 0.00128] | [−0.00125; −0.0011] | [0.743; 0.76] | [0.678; 0.725] | ||
| Species | Intercept | AvTemp | Snow | Precip | CWD |
| s | |||||
| PIMO | 0.0597 | 0.00687 | 2e−04 | 0.000148 | 0.00219 |
| [−0.87; 0.862] | [−0.0295; 0.039] | [−0.000449; 0.000802] | [−0.000483; 0.000799] | [0.00165; 0.0031] | |
| PICO | −0.46 | −0.0394 | −0.00108 | 0.000661 | −0.000566 |
| [−2.34; 1.04] | [−0.0733; −0.00423] | [−0.00246; 0.00036] | [−0.000335; 0.00204] | [−0.00176; 0.00114] | |
| ABMA | 0.425 | 0.0127 | 0.000285 | −0.00018 | −7.16e−05 |
| [0.327; 0.526] | [0.00666; 0.0179] | [0.000183; 0.000404] | [−0.000245; −0.000113] | [−0.000247; 0.000117] | |
| PIJE | −0.197 | 0.0112 | −0.000189 | 0.000262 | 0.000663 |
| [−0.75; 0.308] | [−0.0153; 0.0394] | [−0.000722; 0.000461] | [−0.000221; 0.000697] | [0.000128; 0.00119] | |
| ABCO | 0.343 | 0.00161 | 0.000145 | −4.92e−05 | −0.000116 |
| [0.249; 0.417] | [−0.0018; 0.00482] | [4.17e−05; 0.000238] | [−9.96e−05; 2.06e−05] | [−0.000223; −3.59e−05] | |
| QUKE | 0.453 | 0.0195 | 0.000564 | −0.000387 | −0.000171 |
| [0.159; 0.99] | [0.00587; 0.0307] | [0.000217; 0.0012] | [−0.000779; −0.000171] | [−0.000632; 0.00017] | |
| QUCH | 2.18 | −0.103 | 0.0027 | −0.000904 | 0.00417 |
| [0.517; 4.57] | [−0.221; −0.0119] | [−0.000111; 0.00477] | [−0.00201; 0.00057] | [0.00215; 0.00631] | |
| PILA | 0.636 | −0.00686 | 0.000438 | −0.000274 | −9.36e−05 |
| [0.446; 0.864] | [−0.0136; −0.00134] | [0.000163; 0.000687] | [−0.000439; −0.000131] | [−0.000324; 0.00013] | |
| CADE | 0.431 | −0.00732 | 0.000415 | −9.92e−05 | 1.75e−05 |
| [0.324; 0.564] | [−0.0121; −0.00293] | [0.000239; 0.000593] | [−0.000191; −2.17e−05] | [−0.000148; 0.000191] | |
| PIPO | 0.856 | −0.0498 | 0.000741 | −0.000105 | 0.000528 |
| [0.416; 1.32] | [−0.0873; −0.0106] | [−0.000162; 0.00203] | [−0.000457; 0.000188] | [−0.000355; 0.00164] |
Appendix E. Effect of Different Climate on Growth for Common Species

References
- Pederson, N.; Dyer, J.M.; Mcewan, R.W.; Hessl, A.E.; Mock, C.J.; Orwig, D.A.; Rieder, H.E.; Cook, B.I. The legacy of episodic climatic events in shaping temperate, broadleaf forests. Ecol. Monogr. 2014, 84, 599–620. [Google Scholar] [CrossRef]
- Fischlin, A.; Ayres, M.; Karnosky, D.; Kellomäki, S.; Louman, B.; Ong, C.; Plattner, C.; Santoso, H.; Thompson, I.; Booth, T.; et al. Future environmental impacts and vulnerabilities. Assessment 2009, 22, 53–100. [Google Scholar]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Klos, R.J.; Wang, G.G.; Bauerle, W.L.; Rieck, J.R.; Los, R.; Ang, G.; Auerle, W.; Ieck, J. Drought impact on forest growth and mortality in the southeast USA: An analysis using Forest Health and Monitoring data. Ecol. Appl. 2009, 19, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Kloeppel, B.D.; Clinton, B.D.; Vose, J.M.; Cooper, A.R. Drought Impacts on Tree Growth and Mortality of Southern Appalachian Forests. In Climate Variability and Ecosystem Response at Long-Term Ecological Research Sites; Number 686 m; Oxford University Press: New York, NY, USA, 2003; p. 459. [Google Scholar]
- Fei, S.; Desprez, J.M.; Potter, K.M.; Jo, I.; Knott, J.A.; Oswalt, C.M. Divergence of species responses to climate change. Sci. Adv. 2017. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Summary for Policymakers; Cambeidge University Press: Cambeidge, UK, 2007; pp. 1–36. [Google Scholar]
- Battles, J.J.; Robards, T.; Das, A.; Waring, K.; Gilless, J.K.; Biging, G.; Schurr, F. Climate change impacts on forest growth and tree mortality: A data-driven modeling study in the mixedconifer forest of the Sierra Nevada, California. Clim. Chang. 2008, 87, 193–213. [Google Scholar] [CrossRef]
- Koeppe, C.E.; Long, G.C.D. Weather and Climate; OSS Foundation: Los Angeles, CA, USA, 1958. [Google Scholar]
- Rapacciuolo, G.; Maher, S.P.; Schneider, A.C.; Hammond, T.T.; Jabis, M.D.; Walsh, R.E.; Iknayan, K.J.; Walden, G.K.; Oldfather, M.F.; Ackerly, D.D.; et al. Beyond a warming fingerprint: Individualistic biogeographic responses to heterogeneous climate change in California. Glob. Chang. Biol. 2014, 20, 2841–2855. [Google Scholar] [CrossRef] [PubMed]
- Cook, E.R.; Seager, R.; Cane, M.A.; Stahle, D.W. North American Drought: Reconstructions, Causes, and Consequences. Earth-Sci. Rev. 2007, 81, 93–134. [Google Scholar] [CrossRef]
- Diffenbaugh, N.S.; Swain, D.L.; Touma, D. Anthropogenic warming has increased drought risk in California. Proc. Natl. Acad. Sci. USA 2015, 112, 3931–3936. [Google Scholar] [CrossRef] [PubMed]
- Pierce, D.W.; Das, T.; Cayan, D.R.; Maurer, E.P.; Miller, N.L.; Bao, Y.; Kanamitsu, M.; Yoshimura, K.; Snyder, M.A.; Sloan, L.C.; et al. Probabilistic estimates of future changes in California temperature and precipitation using statistical and dynamical downscaling. Clim. Dyn. 2013, 40, 839–856. [Google Scholar] [CrossRef]
- Hayhoe, K.; Cayan, D.; Field, C.B.; Frumhoff, P.C.; Maurer, E.P.; Miller, N.L.; Moser, S.C.; Schneider, S.H.; Cahill, K.N.; Cleland, E.E.; et al. Emissions pathways, climate change, and impacts on California. Proc. Natl. Acad. Sci. USA 2004, 101, 12422–12427. [Google Scholar] [CrossRef] [PubMed]
- Pierce, D.W.; Cayan, D.R.; Das, T.; Maurer, E.P.; Miller, N.L.; Bao, Y.; Kanamitsu, M.; Yoshimura, K.; Snyder, M.A.; Sloan, L.C.; et al. The key role of heavy precipitation events in climate model disagreements of future annual precipitation changes in california. J. Clim. 2013, 26, 5879–5896. [Google Scholar] [CrossRef]
- Leung, L.R.; Qian, Y.; Bian, X.; Washington, W.M.; Han, J.; Roads, J.O. Mid-century ensemble regional climate change scenarios for the Western united States. Clim. Chang. 2004, 62, 75–113. [Google Scholar] [CrossRef]
- Vicuna, S.; Maurer, E.P.; Joyce, B.; Dracup, J.A.; Purkey, D. The Sensitivity of California Water Resources to Climate Change Scenarios1. JAWRA J. Am. Water Resour. Assoc. 2007, 43, 482–498. [Google Scholar] [CrossRef]
- Dracup, J.; Dracup, J.; Vicuna, S.; Vicuna, S. An Overview of Hydrology and Water Resources Studies on Climate Change: The California Experience. Change 2003, 82, 1–8. [Google Scholar]
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Das, A.J.; Stephenson, N.L. Improving estimates of tree mortality probability using potential growth rate. Can. J. For. Res. 2015, 45, 920–928. [Google Scholar] [CrossRef]
- Wyckoff, P.H.; Clark, J.S. The relationship between growth and mortality for seven co-occurring tree species in the southern Appalachian Mountains. J. Ecol. 2002, 90, 604–615. [Google Scholar] [CrossRef]
- Van Mantgem, P.J.; Stephenson, N.L.; Mutch, L.S.; Johnson, V.G.; Esperanza, A.M.; Parsons, D.J. Growth rate predicts mortality of Abies concolor in both burned and unburned stands. Can. J. For. Res. 2003, 33, 1029–1038. [Google Scholar] [CrossRef]
- Das, A.J.; Battles, J.J.; Stephenson, N.L.; Mantgem, P.J.V.; van Mantgem, P.J. The relationship between tree growth patterns and likelihood of mortality: A study of two tree species in the Sierra Nevada. Can. J. For. Res. 2007, 37, 580–597. [Google Scholar] [CrossRef]
- Clark, J.S.; Iverson, L.; Woodall, C.W.; Allen, C.D.; Bell, D.M.; Bragg, D.C.; D’Amato, A.W.; Davis, F.W.; Hersh, M.H.; Ibanez, I.; et al. The impacts of increasing drought on forest dynamics, structure, and biodiversity in the United States. Glob. Chang. Biol. 2016, 22, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.T.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Yeh, H.Y.; Wensel, L.C. The relationship between tree diameter growth and climate for coniferous species in northern California. Can. J. For. Res. 2000, 30, 1463–1471. [Google Scholar] [CrossRef]
- Bales, R.C.; Goulden, M.L.; Hunsaker, C.T.; Conklin, M.H.; Hartsough, P.C.; O’Geen, A.T.; Hopmans, J.W.; Safeeq, M. Drought impacts on subsurface water storage, evapotranspiration and vegetation mortality in the southern Sierra Nevada, 2009–2015. J. Chem. Inf. Model. 2013, 53, 1689–1699. [Google Scholar]
- Guarin, A.; Taylor, A.H. Drought triggered tree mortality in mixed conifer forests in Yosemite National Park, California, USA. For. Ecol. Manag. 2005, 218, 229–244. [Google Scholar] [CrossRef]
- Millar, C.I.; Westfall, R.D.; Delany, D.L. Response of high-elevation limber pine (Pinus flexilis) to multiyear droughts and 20th-century warming, Sierra Nevada, California, USA. Can. J. For. Res. 2007, 37, 2508–2520. [Google Scholar] [CrossRef]
- Itter, M.S.; Finley, A.O.; D’Amato, A.W.; Foster, J.R.; Bradford, J.B. Variable Effects of Climate on Forest Growth in Relation to Ecosystem State. Ecol. Appl. 2016, 27, 1082–1095. [Google Scholar] [CrossRef] [PubMed]
- Medvigy, D.; Wofsy, S.C.; Munger, J.W.; Moorcroft, P.R. Responses of terrestrial ecosystems and carbon budgets to current and future environmental variability. Proc. Natl. Acad. Sci. USA 2010, 107, 8275–8280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenihan, J.M.; Drapek, R.; Bachelet, D.; Neilson, R.P. Climate Change Effects on Vegetation Distribution, Carbon, and Fire in California. Ecol. Appl. 2003, 13, 1667–1681. [Google Scholar] [CrossRef]
- Lenihan, J.M.; Bachelet, D.; Neilson, R.P.; Drapek, R. Response of vegetation distribution, ecosystem productivity, and fire to climate change scenarios for California. Clim. Chang. 2008, 87, 215–230. [Google Scholar] [CrossRef]
- Dymond, S.F.; D’Amato, A.W.; Kolka, R.K.; Bolstad, P.V.; Sebestyen, S.D.; Bradford, J.B. Growth-climate relationships across topographic gradients in the northern Great Lakes. Ecohydrology 2015, 929, 918–929. [Google Scholar] [CrossRef]
- Williams, A.P.; Seager, R. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Kunstler, G.; Albert, C.H.; Courbaud, B.; Lavergne, S.; Thuiller, W.; Vieilledent, G.; Zimmermann, N.E.; Coomes, D.A. Effects of competition on tree radial-growth vary in importance but not in intensity along climatic gradients. J. Ecol. 2011, 99, 300–312. [Google Scholar] [CrossRef]
- Hurteau, M.; Zald, H.; North, M. Species-specific response to climate reconstruction in upper-elevation mixed-conifer forests of the western Sierra Nevada, California. Can. J. For. Res. 2007, 37, 1681–1691. [Google Scholar] [CrossRef]
- Potito, A.P.; Macdonald, G.M. The Effects of Aridity on Conifer Radial Growth, Recruitment, and Mortality Patterns in The Eastern Sierra Nevada, California. Arct. Antarct. Alpine Res. 2008, 40, 129–139. [Google Scholar] [CrossRef]
- Johnson, C.; Chhin, S.; Zhang, J. Effects of climate on competitive dynamics in mixed conifer forests of the Sierra Nevada. For. Ecol. Manag. 2017, 394, 1–12. [Google Scholar] [CrossRef]
- Das, A. The effect of size and competition on tree growth rate in old-growth coniferous forests. Can. J. For. Res. 2012, 42, 1983–1995. [Google Scholar] [CrossRef]
- Van Mantgem, P.; Das, A. An individual-based growth and competition model for coastal redwood forest restoration. Can. J. For. Res. 2014, 1057, 1–35. [Google Scholar] [CrossRef]
- Mérian, P.; Lebourgeois, F. Size-mediated climate-growth relationships in temperate forests: A multi-species analysis. For. Ecol. Manag. 2011, 261, 1382–1391. [Google Scholar] [CrossRef]
- Evans, M.E.; Falk, D.A.; Arizpe, A.; Swetnam, T.L.; Babst, F.; Holsinger, K.E. Fusing tree-ring and forest inventory data to infer influences on tree growth. Ecosphere 2017, 8. [Google Scholar] [CrossRef]
- Eitzel, M.; Battles, J.; York, R.; Knape, J.; de Valpine, P. Estimating tree growth from complex forest monitoring data. Ecol. Appl. 2013, 23, 1288–1296. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.S.; Wolosin, M.; Dietze, M.; Ibáñez, I.; LaDeau, S.; Welsh, M.; Kloeppel, B. Tree growth inference and prediction from diameter censuses and ring widths. Ecol. Appl. 2007, 17, 1942–1953. [Google Scholar] [CrossRef] [PubMed]
- Bowman, D.M.J.S.; Williamson, G.J.; Keenan, R.J.; Prior, L.D. A warmer world will reduce tree growth in evergreen broadleaf forests: Evidence from Australian temperate and subtropical eucalypt forests. Glob. Ecol. Biogeogr. 2014, 23, 925–934. [Google Scholar] [CrossRef]
- Flint, L.E.; Flint, A.L.; Thorne, J.H.; Boynton, R. Fine-scale hydrologic modeling for regional landscape applications: the California Basin Characterization Model development and performance. Ecol. Process. 2013, 2, 25. [Google Scholar] [CrossRef]
- Hegyi, F. A simulation model for managing jack-pine stands. In Growth Models for Tree and Stand Simulation; Fries, J., Ed.; Royal College of Music, Stockholm: Stockholm, Sweden, 1974; pp. 74–90. [Google Scholar]
- Lorimer, C.G. Tests of age-independent competition indices for individual trees in natural hardwood stands. For. Ecol. Manag. 1983, 6, 343–360. [Google Scholar] [CrossRef]
- Coates, K.D.; Canham, C.D.; LePage, P.T. Above-versus below-ground competitive effects and responses of a guild of temperate tree species. J. Ecol. 2009, 97, 118–130. [Google Scholar] [CrossRef]
- Canham, C.D.; LePage, P.T.; Coates, K.D. A neighborhood analysis of canopy tree competition: Effects of shading versus crowding. Can. J. For. Res. 2004, 34, 778–787. [Google Scholar] [CrossRef]
- Contreras, M.A.; Affleck, D.; Chung, W. Evaluating tree competition indices as predictors of basal area increment in western Montana forests. For. Ecol. Manag. 2011, 262, 1939–1949. [Google Scholar] [CrossRef]
- Slack, A.W.; Kane, J.M.; Knapp, E.E.; Sherriff, R.L. Contrasting impacts of climate and competition on large sugar pine growth and defense in a fire-excluded forest of the central Sierra Nevada. Forests 2017, 8, 244. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P.; et al. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Virlouvet, L.; Fromm, M. Physiological and transcriptional memory in guard cells during repetitive dehydration stress. New Phytol. 2015, 205, 596–607. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. In R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2015. [Google Scholar]
- Sturtz, S.; Ligges, U.; Gelman, A. R2WinBUGS: A Package for Running WinBUGS from R. J. Stat. Softw. 2005, 12, 1–16. [Google Scholar] [CrossRef]
- Bigelow, S.W.; Papaik, M.J.; Caum, C.; North, M.P. Faster growth in warmer winters for large trees in a Mediterranean-climate ecosystem. Clim. Chang. 2014, 123, 215–224. [Google Scholar] [CrossRef]
- Stephenson, N.L. Actual evapotranspiration and deficit: Biologically meaningful correlates of vegetation distribution across spatial scales. J. Biogeogr. 1998, 25, 855–870. [Google Scholar] [CrossRef]
- Stephenson, N.L. Climatic control of vegetation distribution: the role of the water balance. Nat. Am. 1990, 135, 649–670. [Google Scholar] [CrossRef]
- Niemelä, P.; Chapin, F.S.; Danell, K.; Bryant, J.P. Herbivory-mediated responses of selected boreal forests to climatic change. Clim. Chang. 2001, 48, 427–440. [Google Scholar] [CrossRef]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Trân, J.K.; Ylioja, T.; Billings, R.F.; Régnière, J.; Ayres, M.P. Impact of minimum winter temperatures on the population dynamics of Dendroctonus frontalis. Ecol. Appl. 2007, 17, 882–899. [Google Scholar] [CrossRef] [PubMed]
- Hurteau, M.D.; Robards, T.A.; Stevens, D.; Saah, D.; North, M.; Koch, G.W. Modeling climate and fuel reduction impacts on mixed-conifer forest carbon stocks in the Sierra Nevada, California. For. Ecol. Manag. 2014, 315, 30–42. [Google Scholar] [CrossRef]
- D’Amato, A.W.; Bradford, J.B.; Fraver, S.; Palik, B.J. Effects of thinning on drought vulnerability and climate response in north temperate forest ecosystems. Ecol. Appl. 2013, 23, 1735–1742. [Google Scholar] [CrossRef] [PubMed]
- Maloney, P.E.; Smith, T.F.; Jensen, C.E.; Innes, J.; Rizzo, D.M.; North, M.P. Initial tree mortality and insect and pathogen response to fire and thinning restoration treatments in an old-growth mixed-conifer forest of the Sierra Nevada, California. Can. J. For. Res. 2008, 38, 3011–3020. [Google Scholar] [CrossRef]
- Van Mantgem, P.J.; Stephenson, N.L. Apparent climatically induced increase of tree mortality rates in a temperate forest. Ecol. Lett. 2007, 10, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 129. [Google Scholar] [CrossRef]
- Bigler, C. Trade-Offs between Growth Rate, Tree Size and Lifespan of Mountain Pine (Pinus montana) in the Swiss National Park. PLoS ONE 2016. [Google Scholar] [CrossRef] [PubMed]




| Name | Elev (m) | Size (ha) | BA (m) | Est | Dominant Species |
|---|---|---|---|---|---|
| YOHOPIPO | 1500 | 1 | 67.1 | 1991 | ABCO, CADE, PILA |
| BBBPIPO | 1609 | 1 | 68.8 | 1992 | CADE, QUKE |
| CCRPIPO | 1637 | 1.1 | 70.6 | 1991 | CADE, ABCO |
| CRCRPIPO | 1637 | 1 | 71.1 | 1993 | ABCO, CADE |
| FFS7CONTROL | 1941 | 1 | 80.2 | 2001 | ABCO, PILA, CADE |
| FFS6BURN | 2018 | 1 | 50.6 | 2001 | ABCO, PILA |
| FFS5BURN | 2030 | 1 | 78.8 | 2001 | ABCO, CADE |
| SURIP | 2033 | 1.4 | 90.7 | 1982 | ABCO |
| SUABCO | 2035 | 0.9 | 64 | 1983 | ABCO, CADE |
| SUPILA | 2059 | 1.1 | 74.1 | 1983 | ABCO |
| FRPIJE | 2106 | 1 | 19.4 | 1983 | PIJE, QUKE |
| FFS2BURN | 2128 | 1 | 75.9 | 2001 | ABCO, ABMA |
| LMCC | 2128 | 2 | 319.3 | 1982 | ABCO, ABMA |
| LOTHAR | 2167 | 1.1 | 85.9 | 1984 | ABCO |
| LOGSEGI | 2170 | 2.5 | 362.1 | 1983 | ABCO |
| UPTHAR | 2202 | 1 | 81.2 | 1984 | ABCO |
| LOLOG | 2207 | 1.1 | 70 | 1985 | ABCO, ABMA |
| UPLOG | 2210 | 1 | 54.1 | 1985 | ABCO |
| LOGPIJE | 2405 | 1 | 18.3 | 1985 | ABCO, PIJE |
| SFTRABMA | 2484 | 1 | 100.6 | 1992 | ABMA |
| WTABMA | 2521 | 1 | 56.9 | 1993 | ABMA |
| POFLABMA | 2542 | 1 | 105.9 | 1994 | ABMA |
| PGABMA | 2576 | 1 | 96.9 | 1992 | ABMA |
| EMSALIX | 2838 | 1 | 2.5 | 1983 | PIMO, PICO |
| EMSLOPE | 2950 | 1 | 19.7 | 1983 | PIMO |
| EMRIDGE | 3097 | 1.1 | 16.2 | 1984 | PIMO |
| Species Name | Common Name | Code | Number of Trees | Annual Growth |
|---|---|---|---|---|
| Rate (cm) | ||||
| Abies concolor | White fir | ABCO | 6919 | 0.22 |
| Abies magnifica | Red fir | ABMA | 4663 | 0.17 |
| Pinus contorta | Lodgepole pine | PICO | 68 | 0.20 |
| Pinus jeffreyi | Jeffrey pine | PIJE | 282 | 0.20 |
| Pinus monticola | Western white pine | PIMO | 382 | 0.28 |
| Pinus lambertiana | Sugar pine | PILA | 2798 | 0.19 |
| Pinus ponderosa | Ponderosa pine | PIPO | 564 | 0.17 |
| Calocedrus decurrens | Incense cedar | CADE | 4821 | 0.15 |
| Quercus chysolepis | Canyon live oak | QUCH | 75 | 0.082 |
| Quercus kelloggii | Black oak | QUKE | 1274 | 0.12 |
| Climate Variable Name | Description | Unit | Mean Value | Standard Deviation |
|---|---|---|---|---|
| AvTemp | average temperature | 7.15 | 2.15 | |
| JanMin | minimum temperature in January | −5.22 | 2.79 | |
| JulMax | maximum temperature in July | 19.26 | 3.28 | |
| precip | total annual rain precipitation | mm | 1190.72 | 481.82 |
| snow | total annual snowfall | mm | 797.77 | 415.99 |
| ASpck | April snowpack depth | mm | 403.79 | 469.44 |
| CWD | climatic water deficit | mm | 492.42 | 157.42 |
| AET | actual evapotranspiration | mm | 363.69 | 90.66 |
| Parameter | |||
|---|---|---|---|
| Parameter value | 0.28 | 0.002 | −0.01 |
| Parameter estimation | 0.287 | 0.00183 | −0.01020 |
| [0.276; 0.294] | [0.00178; 0.00189] | [−0.01110; −0.00898] | |
| Parameter value | 0.28 | 0.002 | −0.001 |
| Parameter estimation | 0.286 | 0.00183 | −1.07e−03 |
| [0.277; 0.296] | [0.00175; 0.00188] | [−2.26e−03; 6.62e−05] | |
| Parameter value | 0.28 | 0.002 | −0.0001 |
| Parameter estimation | 0.286 | 0.00182 | −5.37e−05 |
| [0.277; 0.297] | [0.00176; 0.00188] | [−1.26e− 03;9.76e−04] |
| PIMO | PICO | ABMA | PIJE | ABCO | QUKE | QUCH | PILA | CADE | PIPO | |
|---|---|---|---|---|---|---|---|---|---|---|
| AvTemp | ↓ | ↑ | ↑ | ↑ | ↑ | ↓ | ↑ | ↓ | ||
| JanMin | ↓ | ↑ | ↑ | ↑ | ↓ | ↓ | ↓ | |||
| JulMax | ↓ | ↑ | ↑ | ↑ | ||||||
| precip | ↓ | ↑ | ||||||||
| snow | ↓ | ↓ | ↑ | ↑ | ||||||
| CWD | ↑ | ↓ | ↑ | ↑ | ↑ | ↓ | ||||
| AET | ↓ | ↑ | ↑ | ↓ | ||||||
| ASpck | ↓ | ↓ | ↑ | |||||||
| AvTemp | ↑ | ↑ | ↓ | ↓ | ||||||
| JanMin | ↓ | ↓ | ↑ | ↑ | ↓ | |||||
| JulMax | ↑ | ↓ | ||||||||
| precip | ↑ | |||||||||
| snow | ↑ | ↑ | ↑ | ↑ | ||||||
| CWD | ↑ | ↑ | ↓ | ↑ | ||||||
| AET | ↑ | ↓ | ||||||||
| ASpck | ↑ | ↑ | ↓ | |||||||
| AvTemp | ↑ | ↓ | ↑ | ↑ | ↑ | ↑ | ↓ | ↓ | ||
| JanMin | ↓ | ↑ | ↑ | ↑ | ↑ | ↓ | ↓ | |||
| JulMax | ↓ | ↑ | ↑ | ↑ | ||||||
| precip | ↑ | ↓ | ↑ | |||||||
| snow | ↓ | ↓ | ↑ | ↑ | ||||||
| CWD | ↑ | ↓ | ↑ | ↑ | ↑ | |||||
| AET | ↓ | ↑ | ↑ | ↓ | ||||||
| ASpck | ↓ | ↓ | ↑ | |||||||
| AvTemp | ↑ | ↓ | ↑ | ↓ | ||||||
| JanMin | ↓ | ↓ | ↑ | ↑ | ↓ | |||||
| JulMax | ↓ | ↑ | ↓ | |||||||
| precip | ↑ | ↑ | ||||||||
| snow | ↑ | ↑ | ↑ | |||||||
| CWD | ↑ | ↓ | ↑ | ↓ | ↓ | |||||
| AET | ↓ | ↓ | ↑ | ↑ | ||||||
| ASpck | ↑ | ↑ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aubry-Kientz, M.; Moran, E.V. Climate Impacts on Tree Growth in the Sierra Nevada. Forests 2017, 8, 414. https://doi.org/10.3390/f8110414
Aubry-Kientz M, Moran EV. Climate Impacts on Tree Growth in the Sierra Nevada. Forests. 2017; 8(11):414. https://doi.org/10.3390/f8110414
Chicago/Turabian StyleAubry-Kientz, Mélaine, and Emily V. Moran. 2017. "Climate Impacts on Tree Growth in the Sierra Nevada" Forests 8, no. 11: 414. https://doi.org/10.3390/f8110414
APA StyleAubry-Kientz, M., & Moran, E. V. (2017). Climate Impacts on Tree Growth in the Sierra Nevada. Forests, 8(11), 414. https://doi.org/10.3390/f8110414