Genetic Differentiation and Population Genetic Structure of the Chinese Endemic Dipteronia Oliv. Revealed by cpDNA and AFLP Data

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
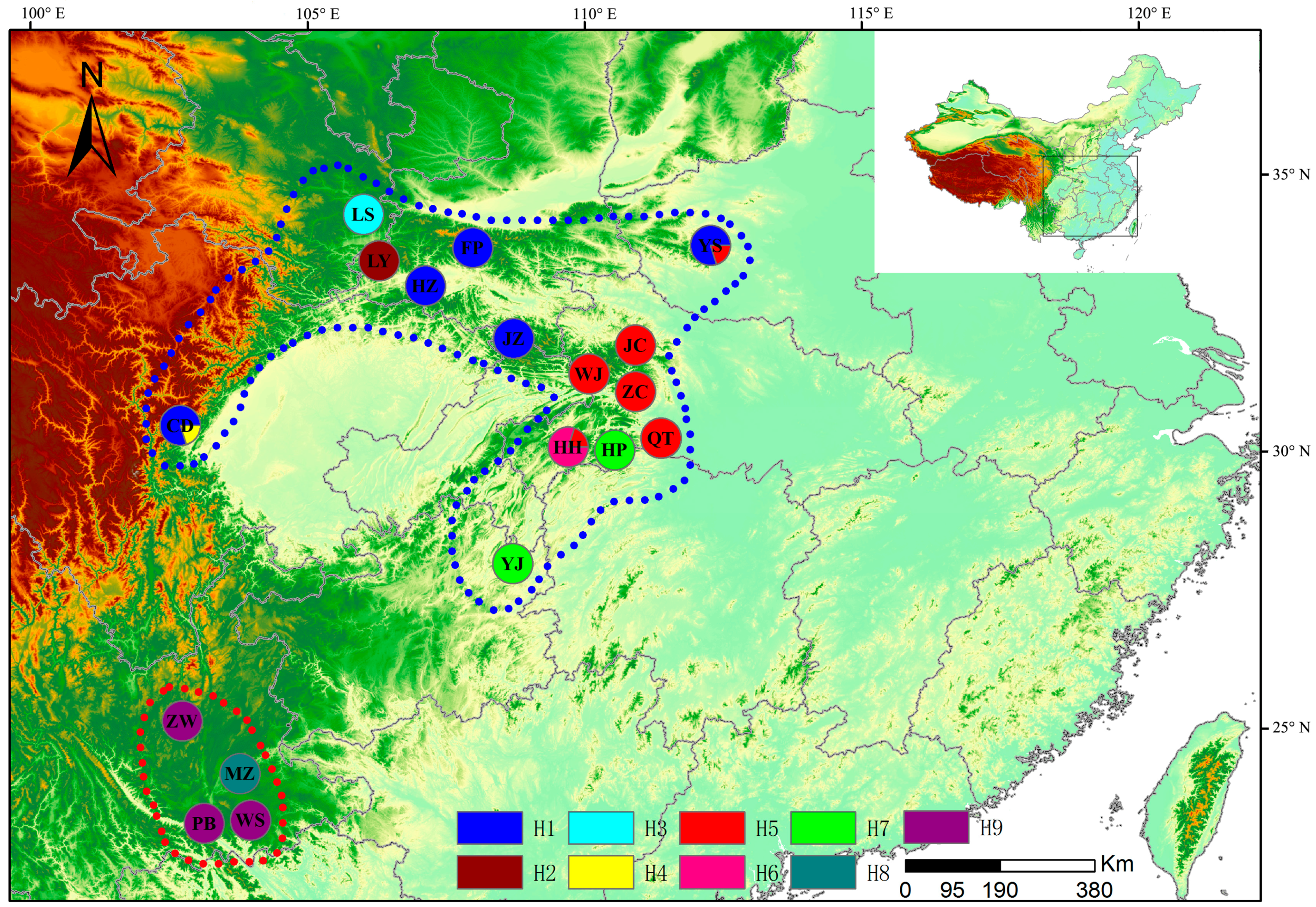
2.1. Sample Collection
2.2. DNA Extraction and PCR Amplification
2.3. Data Analysis
2.3.1. Chloroplast Data Analysis
2.3.2. Analysis of AFLP Datasets
2.3.3. Species Distribution Models
3. Results
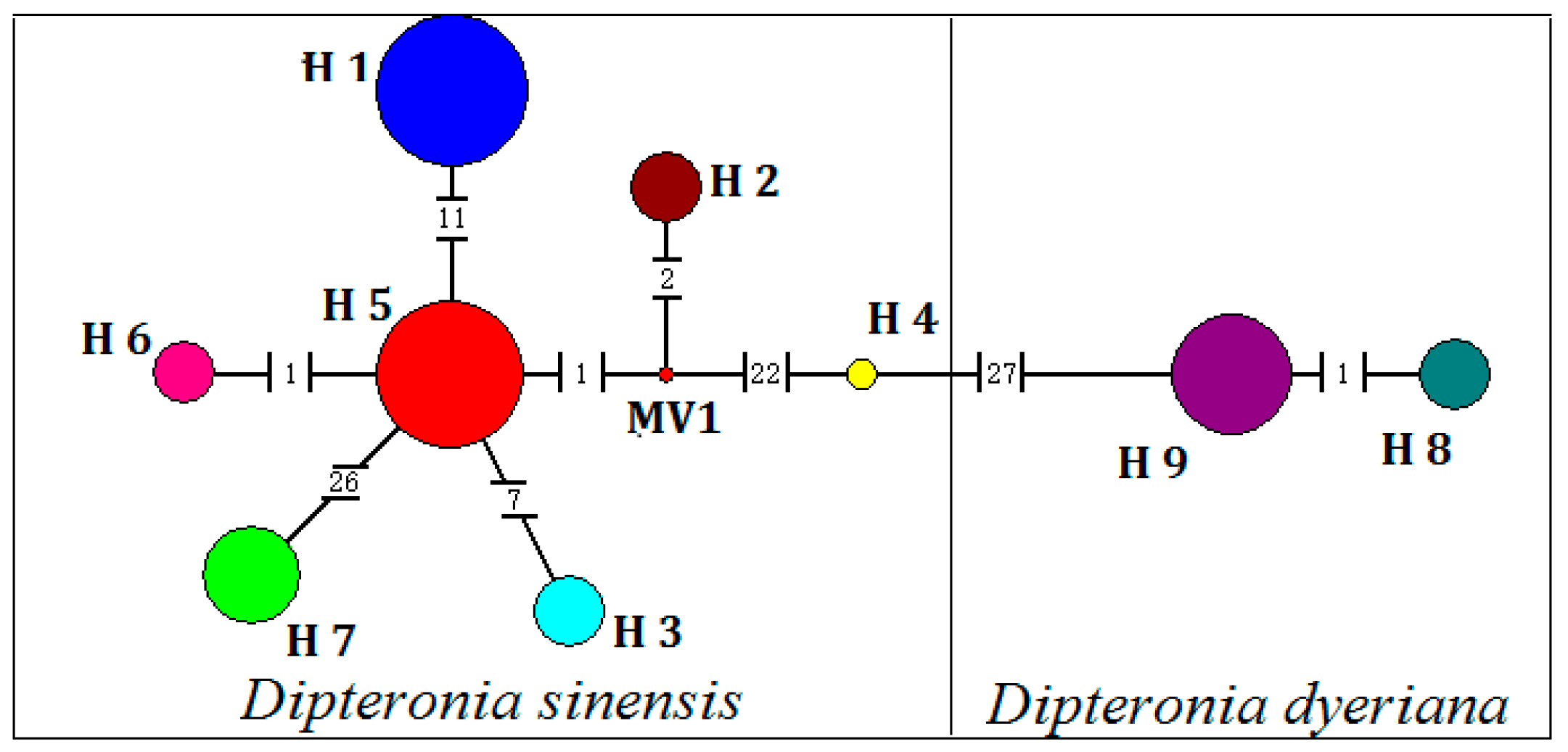
3.1. Chloroplast DNA Polymorphism
3.2. Genetic Diversity and Population Differentiation
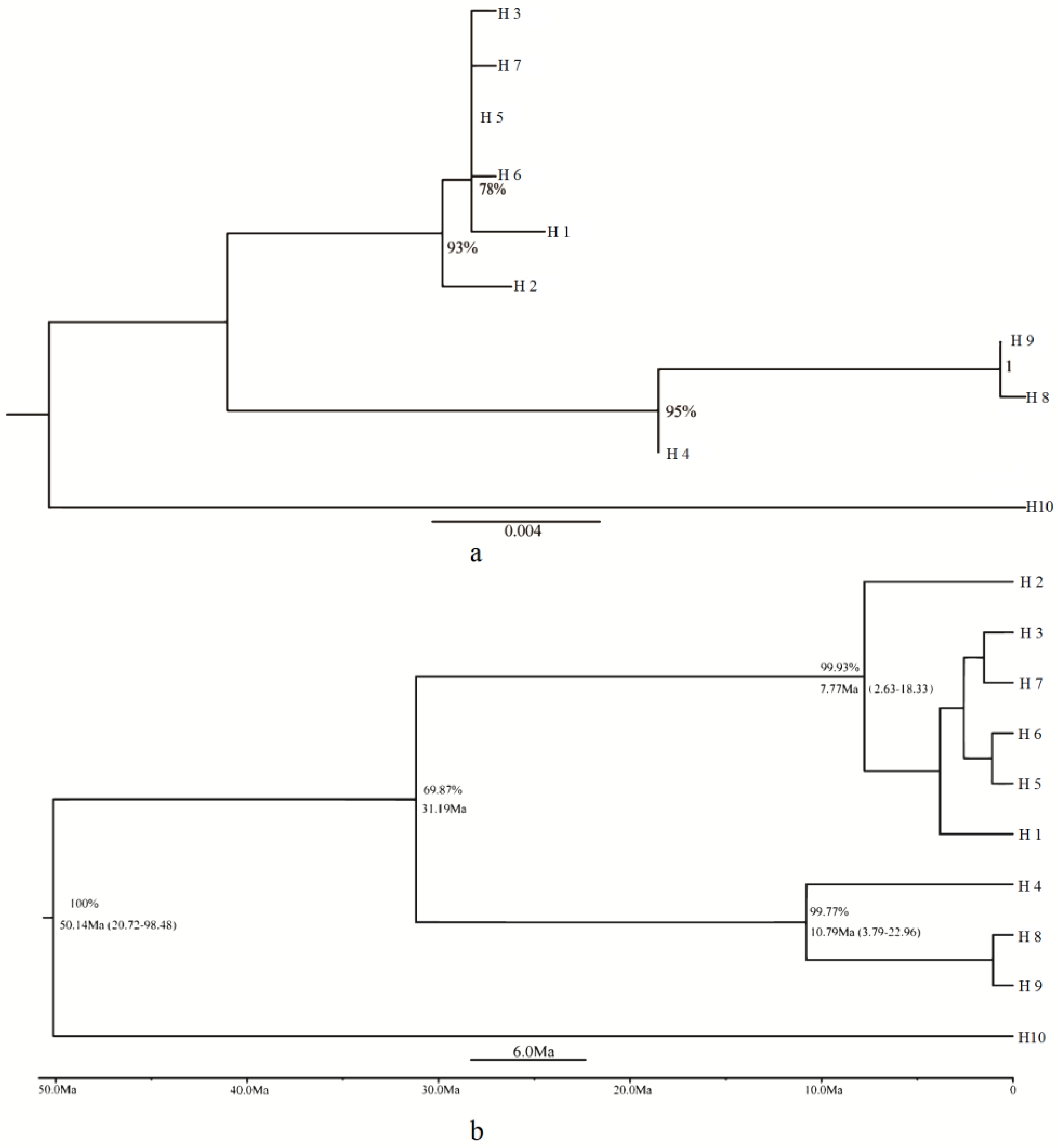
3.3. Phylogeny and Molecular Dating Based on cpDNA Data
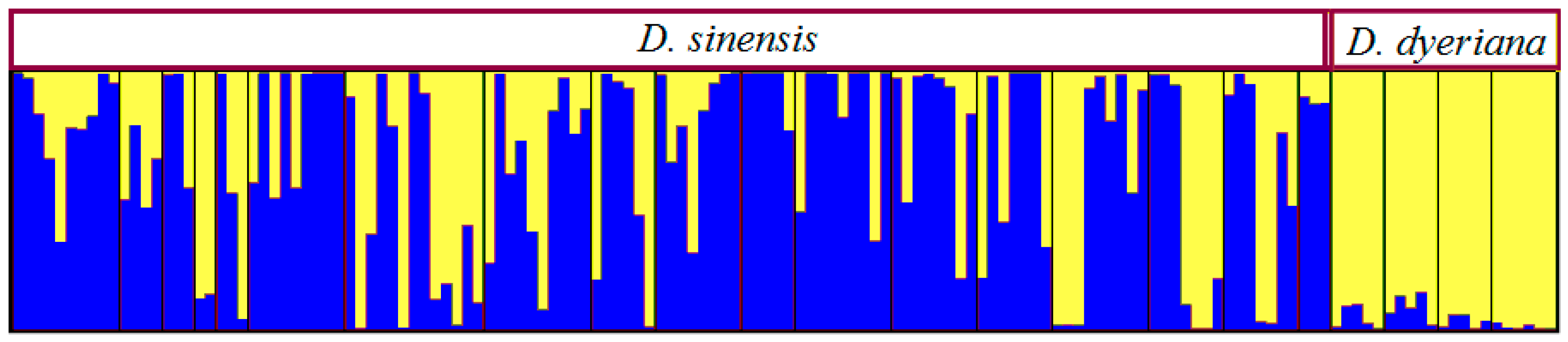
3.4. Population Sturcture and Interspecific Gene Flow Based on AFLP Data
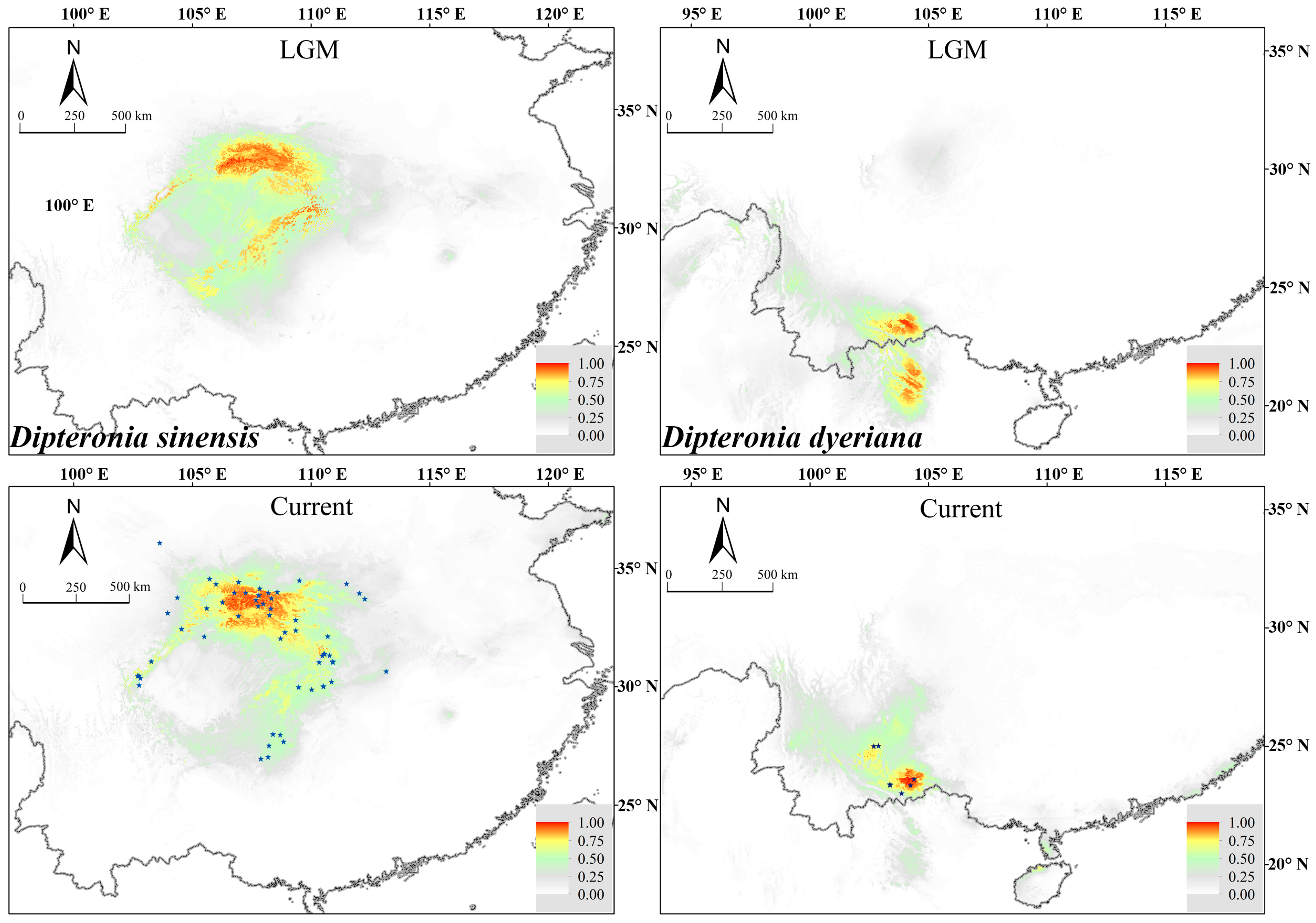
3.5. Present and Past Ecological Niche Models
4. Discussion
4.1. Genetic Diversity, Genetic Differentiation, and Glacial Refuge
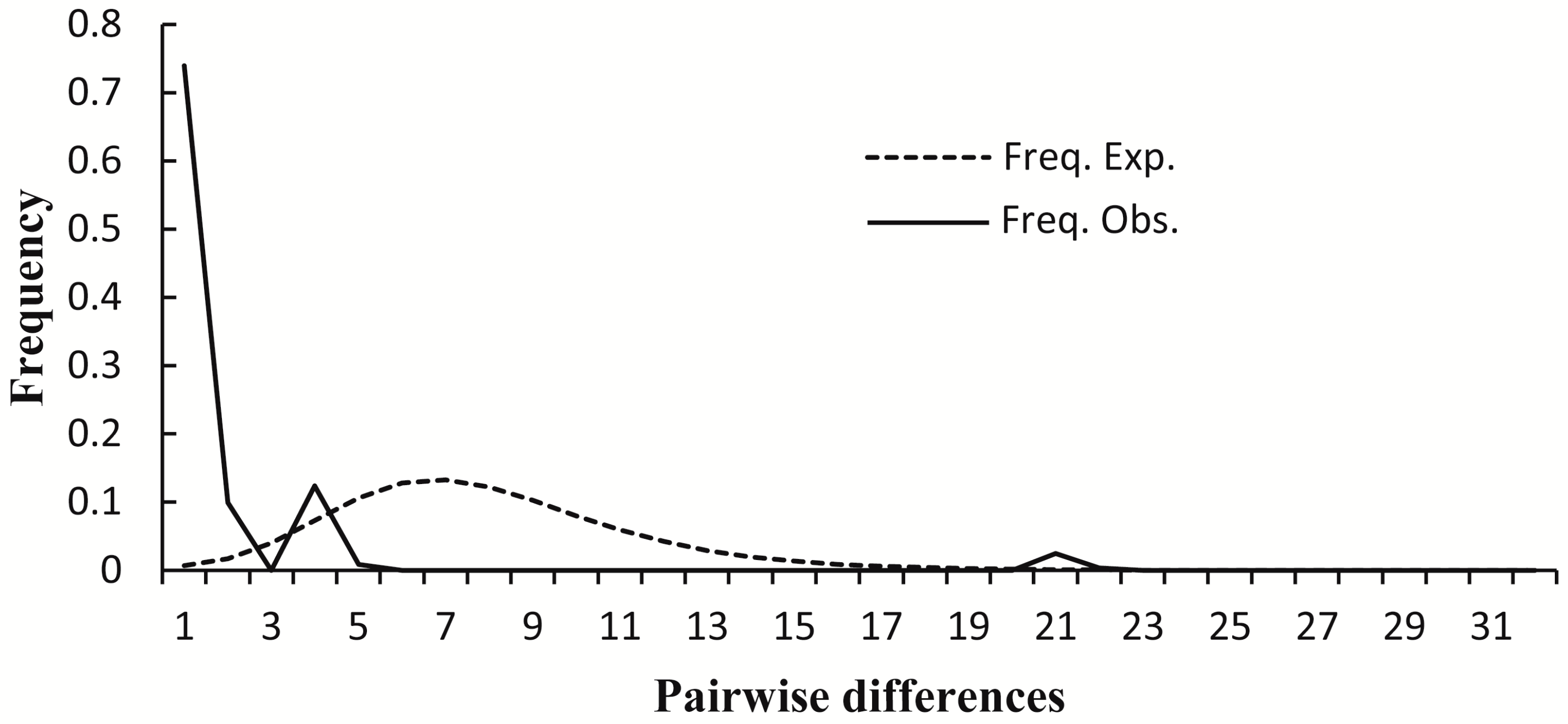
4.2. Demographic History of Dipteronia
4.3. Recommendations for Species Conservation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Avise, J.C. Molecular Markers, Natural History, and Evolution, 2nd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2004; p. 684. [Google Scholar]
- Hewitt, G. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907. [Google Scholar] [CrossRef] [PubMed]
- Abbott, R.J.; Smith, L.C.; Milne, R.I.; Crawfod, R.M.; Wolff, K.; Balfour, J. Molecular analysis of plant migration and refugia in the Arctic. Science 2000, 289, 1343–1346. [Google Scholar] [CrossRef] [PubMed]
- Abbott, R.J.; Brochmann, C. History and evolution of the arctic flora: In the footsteps of Eric Hultén. Mol. Ecol. 2003, 12, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Kropf, M.; Kadereit, J.W.; Comes, H.P. Differential cycles of range contraction and expansion in European high mountain plants during the Late Quaternary: Insights from Pritzelago alpina (L.) O. Kuntze (Brassicaceae). Mol. Ecol. 2003, 12, 931–949. [Google Scholar] [CrossRef] [PubMed]
- Petit, R.J.; Aguinagalde, I.; de Beaulieu, J.L.; Bittkau, C.; Brewer, S.; Cheddadi, R.; Ennos, R.; Fineschi, S.; Grivet, D.; Lasscoux, M.; et al. Glacial refugia: Hotspots but not melting pots of genetic diversity. Science 2003, 300, 1563–1565. [Google Scholar] [CrossRef] [PubMed]
- Petit, R.J.; Duminil, J.; Fineschi, S.; Hampe, A.; Salvoni, D.; Vendramin, G.G. Comparative organization of chloroplast, mitochondrial and nuclear diversity in plant populations. Mol. Ecol. 2005, 14, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chiang, T.Y.; George, M.; Liu, J.Q.; Abbott, R.J. Phylogeography of the Qinghai-Tibetan Plateau enfemic Juniperus przewalskii (Cupressaceae) inferred from chloroplast DNA sequence variation. Mol. Ecol. 2005, 14, 3513–3524. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.M.; Abbott, R.J.; Milne, R.I.; Tian, X.M.; Liu, J.Q. Phylogegraphy of Pinus tabulaefrmis Carr (Pinaceae): A dominant species of coniferous forest in northern China. Mol. Ecol. 2008, 17, 4276–4288. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.X.; Fu, C.X.; Comes, H.P. Plant molecular phylogeography in china adjacent regions, Tracing the genetic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora. Mol. Phylogenet. Evol. 2011, 59, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yan, H.F.; Ge, X.J. Phylogeographic analysis and environmental niche modeling of widespread shrub Rhododendron simsii in China reveals multiple glacial refugia during the last glacial maximum. J. Syst. Evol. 2012, 50, 362–373. [Google Scholar] [CrossRef]
- Hewitt, G.M. Genetic consequences of climatic oscillations in the Quaternary. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.W.; Ge, S. Phylogeography of the endangered Cathaya argyrophylla (Pinaceae) inferred from sequence variation of mitochondrial and nuclear DNA. Mol. Ecol. 2006, 15, 4109–4122. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, H.; Setoguchi, H. Phylogeography and refugia of the Japanese endemic alpine plant, Phyllodoce nipponica Makino (Ericaceae). J. Biogeogr. 2007, 34, 169–174. [Google Scholar] [CrossRef]
- Heuertz, M.; Fineschi, S.; Anzidei, M.; Pastorelli, R.; Salvini, D.; Paule, L.; Rascaria-Lacoste, N.; Hardy, O.J.; Vekemans, X.; Vendramin, G.G. Chloroplast DNA variation and postglacial recolonization of common ash (Fraxinus excelsior L.) in Europe. Mol. Ecol. 2004, 13, 3437–3452. [Google Scholar] [CrossRef] [PubMed]
- Waugh, R.; Bonar, N.; Baird, E.; Thomas, B.; Graner, A.; Hayes, P.; Powell, W. Homology of AFLP products in three mapping populations of barley. Mo. Genet. Genomics 1997, 255, 311–321. [Google Scholar]
- Schut, J.W.; Qi, X.; Stam, P. Association between relationship measures based on AFLP markers, pedigree data and morphologicak traits in barley. Theor. Appl. Genet. 1997, 95, 1161–1168. [Google Scholar] [CrossRef]
- Yuan, L.X.; Fu, J.H.; Warburton, M.; Li, X.H.; Zhang, S.H. Comparison of genetic diversity among maize inbred lines based on RFLPs, SSRs, AFLPs and RAPDs. Acta Gene Sin 2000, 27, 725–733. [Google Scholar]
- Amy, M.M.; Steven, R.M. Dipteronia (Sapindaceae) from the tertiary of North America and implications for the phytogeographic history of the Aceroideae. Am. J. Bot. 2001, 88, 1316–1325. [Google Scholar]
- Ying, J.S.; Zhang, Y.L. Endemic Genera of Chinese Seed Plants; Science Press: Beijing, China, 1994; pp. 57–59. [Google Scholar]
- Tian, X.; Xu, T.Z.; Li, D.Z. New data of some Aceraceae plants in Yunnan, China. Acta Bot. Yunnanica 2001, 23, 291–292. [Google Scholar]
- Wang, S.; Xie, Y. China Species Red List; Science Press: Beijing, China, 2004; p. 357. [Google Scholar]
- Chen, C.; Lu, R.S.; Zhu, S.S.; Tamaki, L.; Qiu, Y.X. Population structure and historical demography of Dipteronia dyeriana (Sapindaceae), an extremely narrow palaeoendemic plant from China: Implications for conservation in a biodiversity hot spot. Heredity 2017, 119, 95–106. [Google Scholar] [CrossRef] [PubMed]
- The Writing Group of Cenozoic Plants of China (WGCPC). Cenozoic Plants from China, Fossil Plants of China; Science Press: Beijing, China, 1978. [Google Scholar]
- Manchester, S.R. Biogeographical relationships of North American Tertiary floras. Ann. Mo. Bot. Gard. 1999, 86, 472–522. [Google Scholar] [CrossRef]
- Liu, D.S. Quaternary Environment; Science Press: Beijing, China, 1997; pp. 14–38. [Google Scholar]
- Xia, Z.K. Quaternary Environmental Science; Beijing University Press: Beijing, China, 1997; pp. 105–118. [Google Scholar]
- Li, S.G. Quaternary Glaciation of China; Science Press: Beijing, China, 1975; pp. 118–120. [Google Scholar]
- Li, W.Q. The Chinese Quaternary Vegetation and Environment; Science Press: Beijing, China, 1998; pp. 183–210. [Google Scholar]
- Zhao, X.G.; Xiao, L.; Mao, F.C. Pollen morphology of Aceraceae. Acta Bot. Boreal.-Occident. Sin. 1998, 18, 252–255. [Google Scholar]
- Tian, X.; Jin, Q.J.; Li, D.Z. Pollen morphology of Aceraceae and its systematic implication. Acta Bot. Yunnanica 2001, 23, 457–465. [Google Scholar]
- Qi, Y.Z.; Du, Y.J.; Li, L.M. The rare and endangered plants of Shaanxi Province for ex situ conservation in Xi’an Botanical Garden. J. Northwest For. Coll. 2001, 16, 33–36. [Google Scholar]
- Li, S.; Cai, Y.L.; Xu, L.; Zhao, G.F. Morphological differentiation of samaras and seeds of Dipteronia dyeriana, Aceraceae. Acta Bot. Yunnanica 2003, 25, 589–595. [Google Scholar]
- Li, S.; Qian, Z.Q.; Cai, Y.L.; Zhao, G.F. A comparative study on the genetic diversity of Dipteronia sinensis and Dipteronia dyeriana. Acta Phytoecol. Sin. 2005, 29, 785–792. [Google Scholar]
- Yang, J.; Qian, Z.Q.; Liu, Z.L.; Li, S.; Sun, G.L.; Zhao, G.F. Genetic diversity and geographical differentiation of Dipteronia Oliv.(Aceraceae) endemic to China as revealed by AFLP analysis. Biochem. Syst. Ecol. 2007, 35, 593–599. [Google Scholar] [CrossRef]
- Wang, G.L.; Fang, H.J. Principles and Techniques of Plant Gene Engineering; Science Press: Beijing, China, 1998; pp. 370–375. [Google Scholar]
- Yang, J.; Li, S.; Cao, D.W.; Liu, Z.L.; Zhao, G.F. Construction and analysis of the AFLP reaction system of Dipteronia Oliv. Acta Bot. Boreal.-Occident. Sin. 2005, 25, 2173–2177. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The ClustalX windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 24, 4876–4882. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987; pp. 10–88. [Google Scholar]
- Rozas, J.; Sánchez-DelBarrio, J.C.; Messeguer, X.; Rozas, R. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 2003, 19, 2496–2497. [Google Scholar] [CrossRef] [PubMed]
- Pons, O.; Petit, R.J. Measuring and testing genetic differentiation with ordered versus unordered alleles. Genetics 1996, 144, 1237–1245. [Google Scholar] [PubMed]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. Modeltest: testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.Y.; Thomas, D.T.; Xiang, Q.P. Resolving and dating the phylogeny of Cornales-Effects of taxon sampling, data partitions, and fossil calibrations. Mol. Phylogenet. Evol. 2011, 59, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.D.; Wang, H.F.; Bao, L.; Wang, T.M.; Bai, W.N.; Ye, J.W.; Ge, J.P. Evolutionary history of a widespread tree species Acer mono in East Asia. Ecol. Evol. 2014, 4, 4332–4345. [Google Scholar] [PubMed]
- Rambaut, A.; Drummond, A.J. Tracerv1.4. Available online: http://beast.bio.ed.ac.uk/Tracer/ (accessed on 23 April 2017).
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.N.; Comes, H.P.; Sakaguchi, S.; Chen, L.Y.; Qiu, Y.X. Evolution of East Asia’s Arcto-Tertiary relict Euptelea (Eupteleaceae) shaped by Late Neogene vicariance and Quaternary climate change. BMC Evol. Biol. 2016, 16, 66. [Google Scholar] [CrossRef] [PubMed]
- Beerli, P. Comparison of Bayesian and maximum-likelihood inference of population genetic parameters. Bioinformatics 2006, 22, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Beerli, P.; Felsenstein, J. Maximum-likelihood estimation of migration rates and effective population numbers in two populations using a coalescent approach. Genetics 1999, 152, 763–773. [Google Scholar] [PubMed]
- Bai, W.N.; Liao, W.J.; Zhang, D.Y. Nuclear and chloroplast DNA phylogeography reveal two refuge areas with asymmetrical gene flow in a temperate walnut tree from East Asia. New Phytol. 2010, 188, 892–901. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- The Chinese Virtual Herbarium. Available online: http://www.cvh.org.cn (accessed on 12 June 2015).
- The National Specimen Information Infrastructure of China. Available online: http://www.nsii.org.cn (accessed on 19 October 2015).
- The WorldClim Database. Available online: http://www.worldclim.org (accessed on 13 January 2017).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolate climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Peterson, A.T.; Nakazawa, Y. Environmental data sets matter in ecological niche modelling: An example with Solenopsis invicta and Solenopsis richteri. Glob. Ecol. Biogeogr. 2008, 17, 135–144. [Google Scholar] [CrossRef]
- The Community Climate System Model. Available online: http://pmip2.lsce.ipsl.fr/ (accessed on 27 March 2017).
- Collins, W.D.; Bitz, C.M.; Blackmon, M.L.; Bonan, G.B.; Bretherton, C.S.; Carton, J.A.; Chang, P.; Doney, S.C.; Hack, J.J.; Henderson, T.B. The Community Climate System Model version 3 (CCSM3). J. Clim. 2006, 19, 2122–2143. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Sakurai, S.; Yamasaki, M.; Isagi, Y. How did the exposed seafloor function in postglacial northward range expansion of Kalopanax septemlobus? Evidence from ecological niche modelling. Ecol. Res. 2010, 25, 1183–1195. [Google Scholar] [CrossRef]
- Fawcett, T. An introduction to ROC analysis. Pattern Recogn. Lett. 2006, 27, 861–874. [Google Scholar] [CrossRef]
- Chiang, T.Y.; Schaal, B.A. Phylogeography of North American populations of the moss species Hylocomium splendens, based on the nucleotide sequence of internal transcribed spacer 2 of nuclear ribosomal DNA. Mol. Ecol. 1999, 8, 1037–1042. [Google Scholar] [CrossRef]
- Huang, S.; Chiang, Y.C.; Schaal, B.A.; Chou, C.H.; Chiang, T.Y. Organelle DNA phylogeography of Cycas taitungensis, a relict species in Taiwan. Mol. Ecol. 2001, 10, 2669–2681. [Google Scholar] [CrossRef] [PubMed]
- Crandall, K.A.; Templeton, A.R. Empirical tests of some predictions from coalescent theory with applications to intraspecific phylogeny reconstruction. Genetics 1993, 134, 959–969. [Google Scholar] [PubMed]
- Wang, H.S. Floristic Geography; Science Press: Beijing, China, 1992. [Google Scholar]
- Ying, T.S. Species diversity and distribution pattern of seed plants in China. Biodivers. Sci. 2001, 9, 393–398. [Google Scholar]
- Axelrod, D.I.; Al-Shehba, I.; Raven, P.H. History of the modern flora of China. In Floristic Characteristics and Diversity of East Asian Plants; Zhang, A.L., Wu, S.G., Eds.; Springer: New York, NY, USA, 1996; pp. 43–55. [Google Scholar]
- Harrison, S.P.; Yu, G.; Takahara, H.; Prentice, I.C. Palaeovegetation: Diversity of temperate plants in East Asia. Nature 2001, 413, 129–130. [Google Scholar] [CrossRef] [PubMed]
- Ray, N.; Adams, J.M. A GIS-based Vegetation Map of the Worldat the Last Glacial Maximum (25,000–15,000 BP). Internetarchaeology 2001, 11. Available online: http://www.ncdc.noaa.gov/ (accessed on 14 February 2017).
- Moritz, C. Defining significant units for conservation. Trends Ecol. Evol. 1994, 9, 373–375. [Google Scholar] [CrossRef]
- Rita, J.; Cursach, J. Creating new populations of Apium bermejoi (Apiaceae), a critically endangered endemic plant on Menorca (Balearic Islands). Anales Del Jardín Botánico De Madrid 2013, 70, 27–38. [Google Scholar] [CrossRef]
- Fernández-Mazuecos, M.; Jiménez-Mejías, P.; Rotllan-Puig, X.; Vargas, P. Narrow endemics to Mediterranean islands: Moderate genetic diversity but narrow climatic niche of the ancient, critically endangered Naufraga (Apiaceae). Perspect. Plant Ecol. 2014, 16, 190–202. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Sample Location | Latitude | Longitude | Altitude (m) | Number | Haplotype (Sample Number) |
|---|---|---|---|---|---|---|
| D. sinensis | ||||||
| FP | Fo-ping | 33°40′ | 107°41′ | 1154 | 15 | H1 (15) |
| HZ | Han-zhong | 32°08′ | 105°30′ | 1603 | 15 | H1 (15) |
| LY | Lue-yang | 33°35′ | 106°16′ | 1346 | 15 | H2 (15) |
| LS | Long-shan | 34°21′ | 106°00′ | 1526 | 15 | H3 (15) |
| CD | Chuan-dongzi | 30°28′ | 102°42′ | 1468 | 15 | H1 (12), H4 (3) |
| JZ | Jian-zhu | 32°03′ | 108°43′ | 1479 | 15 | H1 (15) |
| YS | Yao-shan | 33°43′ | 112°16′ | 1138 | 15 | H1 (12), H5 (3) |
| HH | Hou-he | 31°19′ | 110°29′ | 1297 | 15 | H5 (3), H6 (12) |
| JC | Jiu-chongshan | 31°24′ | 110°33′ | 870 | 15 | H5 (15) |
| QT | Qing-tanwan | 31°03′ | 110°55′ | 1685 | 15 | H5 (15) |
| WJ | Wan-jiagou | 31°24′ | 110°33′ | 811 | 15 | H5 (15) |
| ZC | Zhu-caogou | 31°05′ | 110°55′ | 1735 | 15 | H5 (15) |
| HP | Hu-pingshan | 30°01′ | 110°31′ | 1500 | 15 | H7 (15) |
| YJ | Yin-jiang | 27°59’ | 108°42′ | 1098 | 15 | H7 (15) |
| D. dyeriana | ||||||
| MZ | Meng-zi | 23°24′ | 103°23′ | 1902 | 15 | H8 (15) |
| PB | Ping-bian | 23°01′ | 103°52′ | 2019 | 15 | H9 (15) |
| WS | Wen-shan | 23°37′ | 104°24′ | 2217 | 15 | H9 (15) |
| ZW | Zhi-wuyuan | 25°02′ | 102°54′ | 1923 | 15 | H9 (15) |
| Region | Size Range (bp) | Total Number of Mutations | N° Polymorphic Sites | Parsimony Informative Sites (Two Variants) |
|---|---|---|---|---|
| trnV intron | 408 | 6 | 6 | 6 |
| rps18-rpl20 | 415 (404–415) | 6 | 6 | 6 |
| rpl20-rps12 | 560 (559–560) | 10 | 10 | 10 |
| trnH-psbA | 356 (356–399) | 15 | 14 | 14 |
| Species | π × 10−3 | Hd | HT | HS | GST | NST | Fu’s Fs (p-Value) | Tajima’s D (p-Value) |
|---|---|---|---|---|---|---|---|---|
| D. sinensis | 0.63 | 0.260 | 0.812 | 0.086 | 0.894 | 0.912 | 19.053 (p = 0.998) | –2.365 (p = 0.000) |
| D. dyeriana | 0.23 | 0.395 | 0.500 | 0 | 1 | 1 | 0.976 (p = 0.534) | 0.723 (p = 0.851) |
| Markers | Source of Variation | df | Sum of Squares | Variance Components | Percentage of Variation (%) | Fixation Index |
|---|---|---|---|---|---|---|
| Between species | 1 | 6.734 | 0.160 Va | 29.92 | FCT: 0.299 | |
| Among populations within species | 16 | 27.921 | 0.342 Vb | 63.87 | FSC: 0.911 | |
| Within populations | 72 | 2.400 | 0.033 Vc | 6.22 | FST: 0.938 ** | |
| Total | 89 | 37.056 | 0.536 | |||
| D. sinensis | Among populations | 13 | 24.171 | 0.363 Va | 89.45 | FST: 0.894 |
| Within populations | 56 | 2.400 | 0.043 Vb | 10.55 | ||
| Total | 69 | 26.571 | 0.406 | |||
| D. dyeriana | Among populations | 3 | 3.750 | 0.250 Va | 100 | FST: 1.000 |
| Within populations | 16 | 0.000 | 0.000 Vb | 0 | ||
| Total | 19 | 3.750 | 0.250 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, G.; Zhou, T.; Zhang, X.; Chen, X.; Yang, J.; Li, Z.; Zhao, G. Genetic Differentiation and Population Genetic Structure of the Chinese Endemic Dipteronia Oliv. Revealed by cpDNA and AFLP Data. Forests 2017, 8, 424. https://doi.org/10.3390/f8110424
Bai G, Zhou T, Zhang X, Chen X, Yang J, Li Z, Zhao G. Genetic Differentiation and Population Genetic Structure of the Chinese Endemic Dipteronia Oliv. Revealed by cpDNA and AFLP Data. Forests. 2017; 8(11):424. https://doi.org/10.3390/f8110424
Chicago/Turabian StyleBai, Guoqing, Tao Zhou, Xiao Zhang, Xiaodan Chen, Jia Yang, Zhonghu Li, and Guifang Zhao. 2017. "Genetic Differentiation and Population Genetic Structure of the Chinese Endemic Dipteronia Oliv. Revealed by cpDNA and AFLP Data" Forests 8, no. 11: 424. https://doi.org/10.3390/f8110424
APA StyleBai, G., Zhou, T., Zhang, X., Chen, X., Yang, J., Li, Z., & Zhao, G. (2017). Genetic Differentiation and Population Genetic Structure of the Chinese Endemic Dipteronia Oliv. Revealed by cpDNA and AFLP Data. Forests, 8(11), 424. https://doi.org/10.3390/f8110424