Product and Residue Biomass Equations for Individual Trees in Rotation Age Pinus radiata Stands under Three Thinning Regimes in New South Wales, Australia


Abstract
:1. Introduction
2. Material and Methods
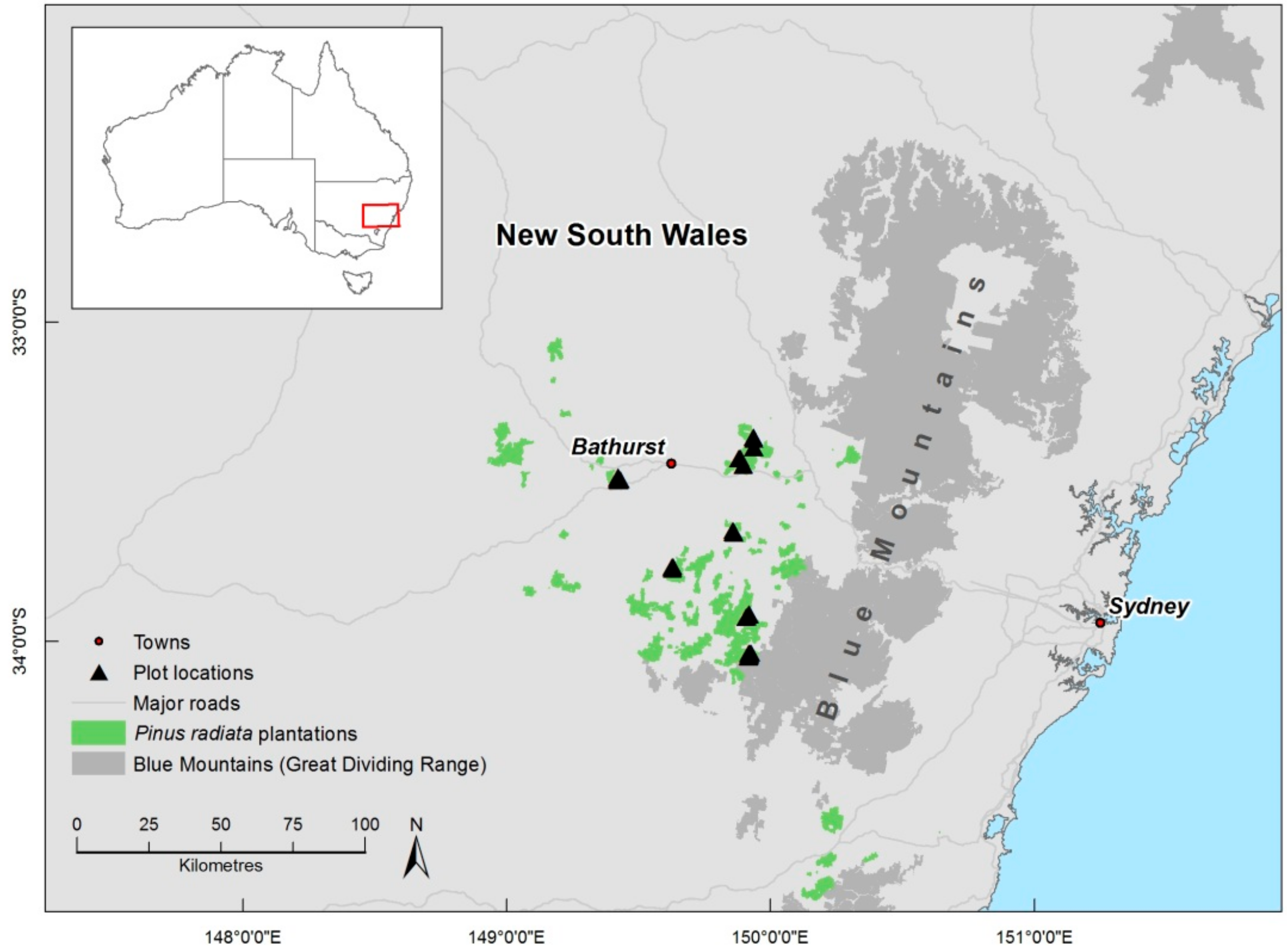
2.1. Study Area
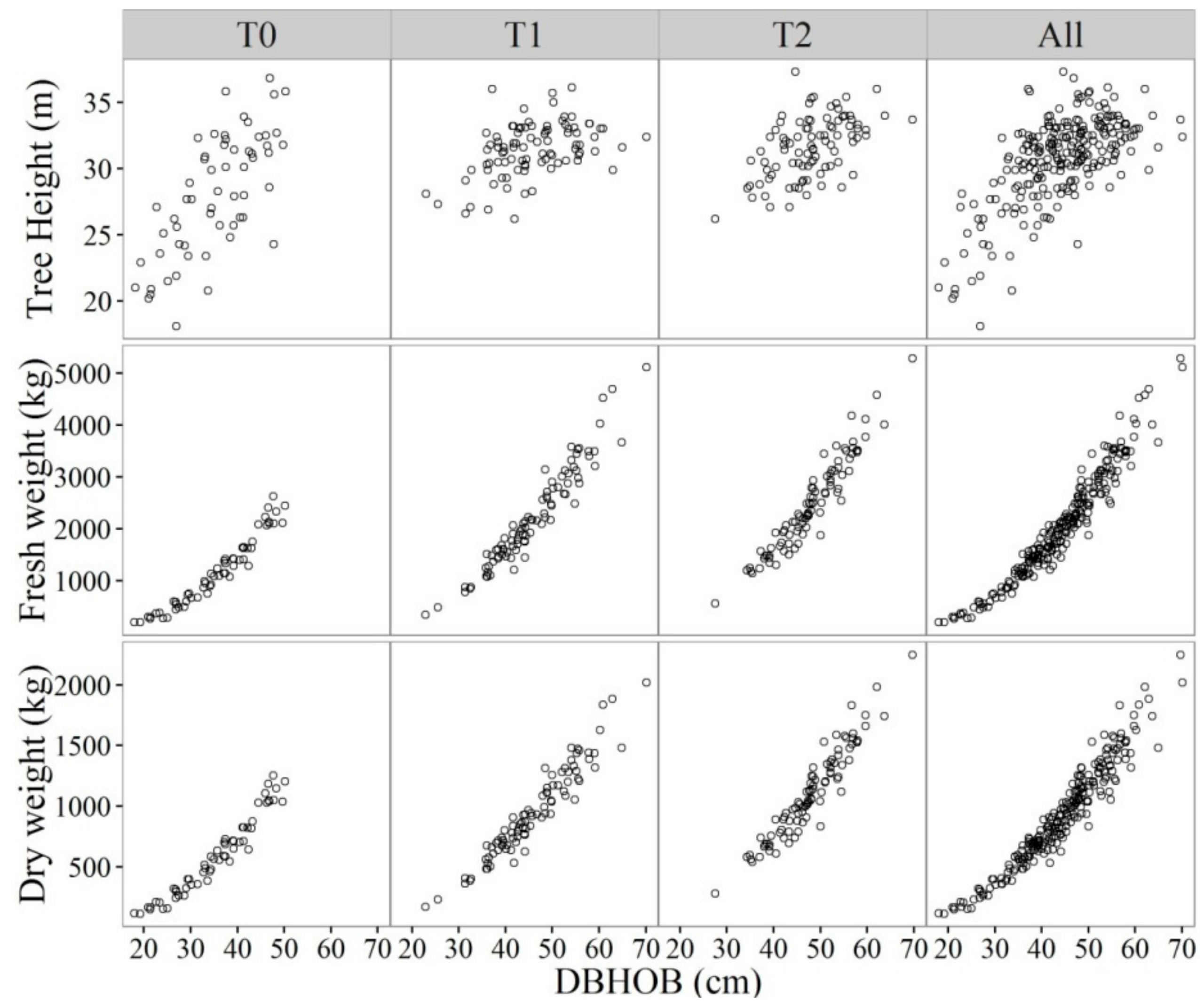
2.2. Plot Measurements and Selection of Sample Trees
2.3. Destructive Sampling in the Field
2.4. Sample Processing and Oven Drying
2.5. Calculating Stump Fresh Weight
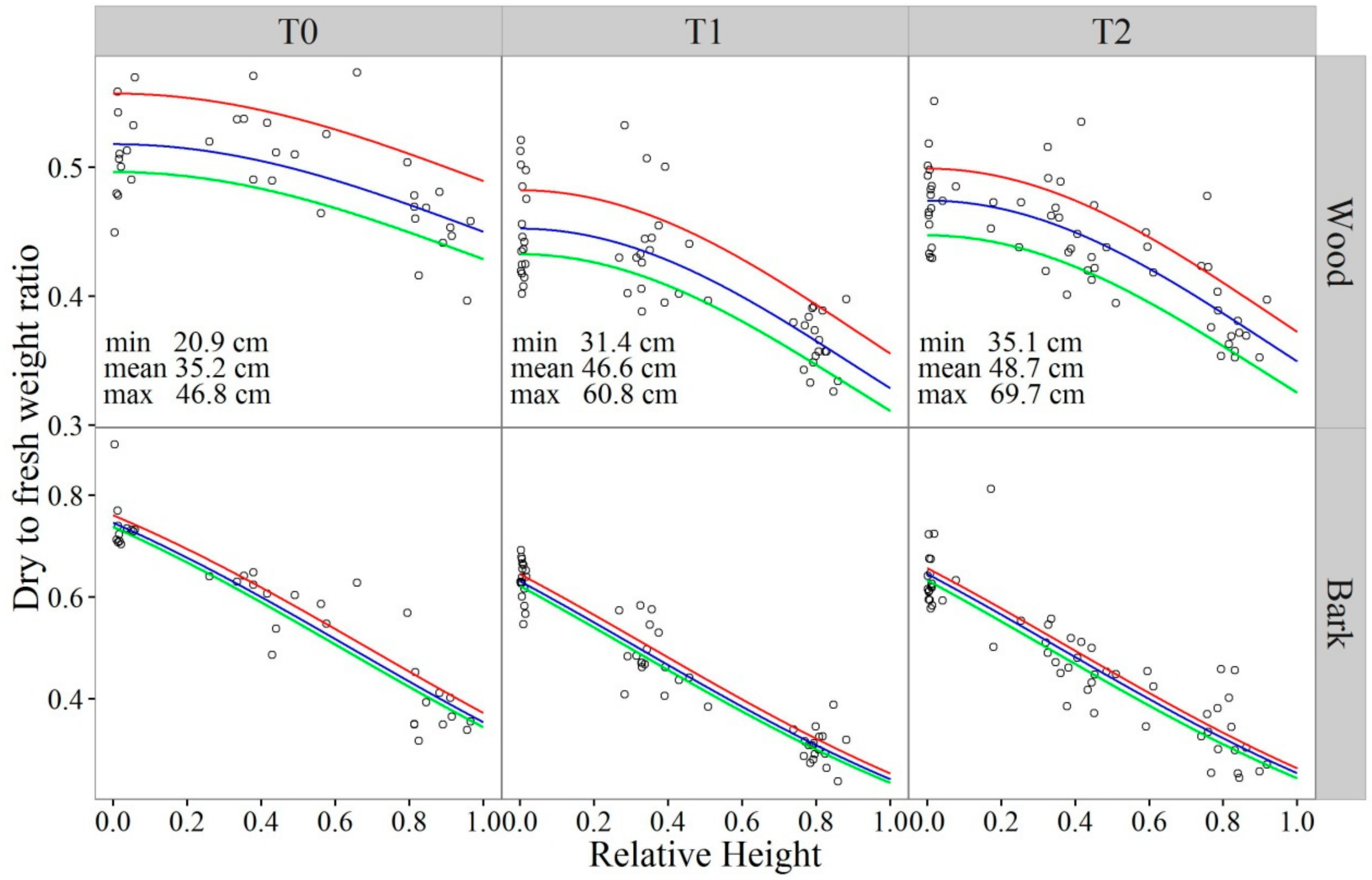
2.6. Estimating Dry to Fresh Weight Ratios of Stemwood and Bark by Beta Regression
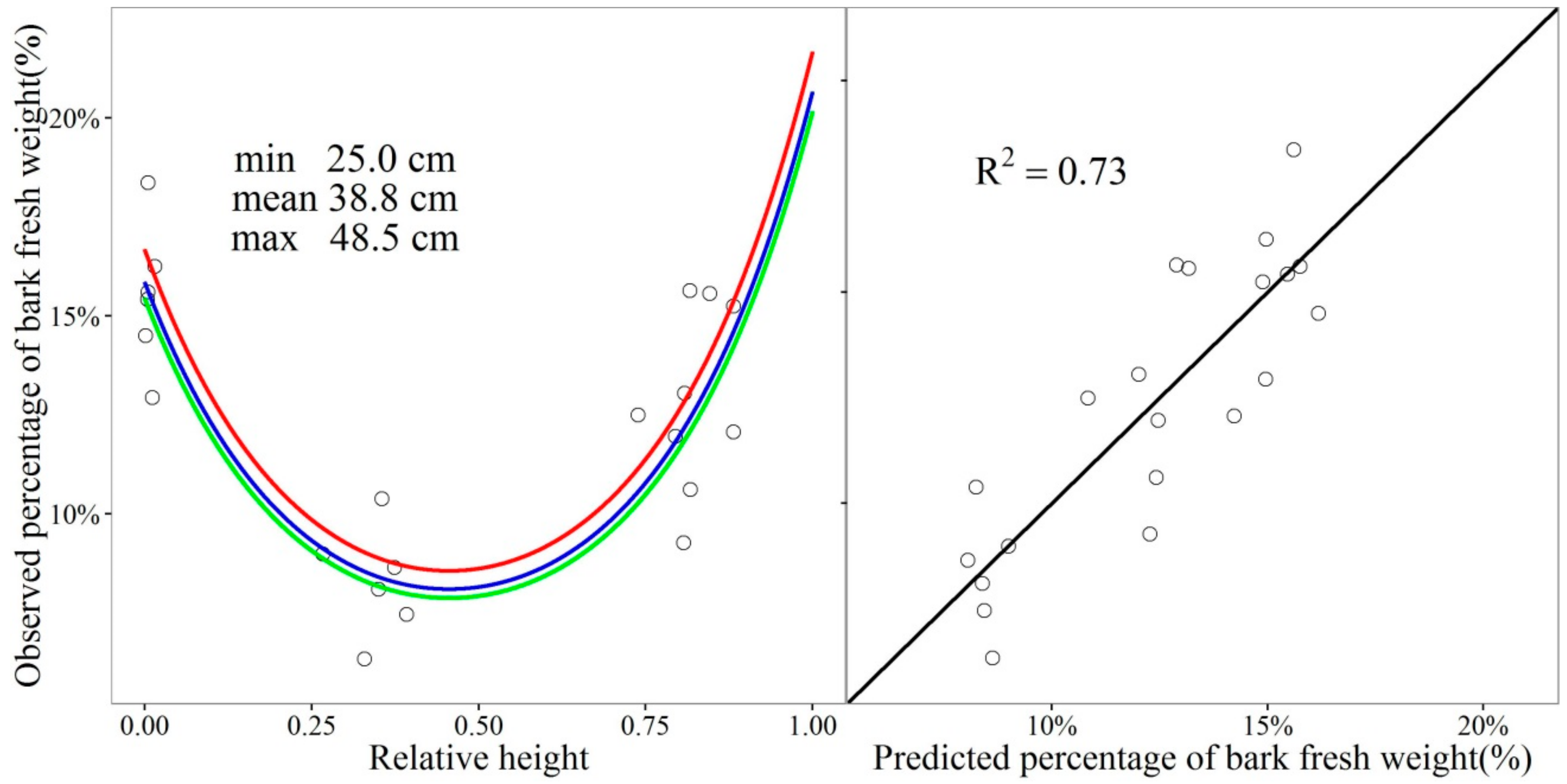
2.7. Estimating the Percentage of Bark Fresh Weight of Stem Sections
2.8. Converting Fresh Weight to Dry Weight
2.9. Systems of Additive and Allocative Biomass Equations
2.9.1. Additive Biomass Equations
2.9.2. Allocative Biomass Equations
2.9.3. Residual Variance and Approximate Confidence Band of Residuals
2.9.4. Evaluating Prediction Accuracy
3. Results
3.1. Dry to Fresh Weight Ratios
3.2. The Percentage of Bark in the Total Fresh Weight of a Stem Cross-Section
3.3. Converting Fresh Weight to Dry Weight
3.4. Additive Biomass Equations
3.5. Allocative Biomass Equations
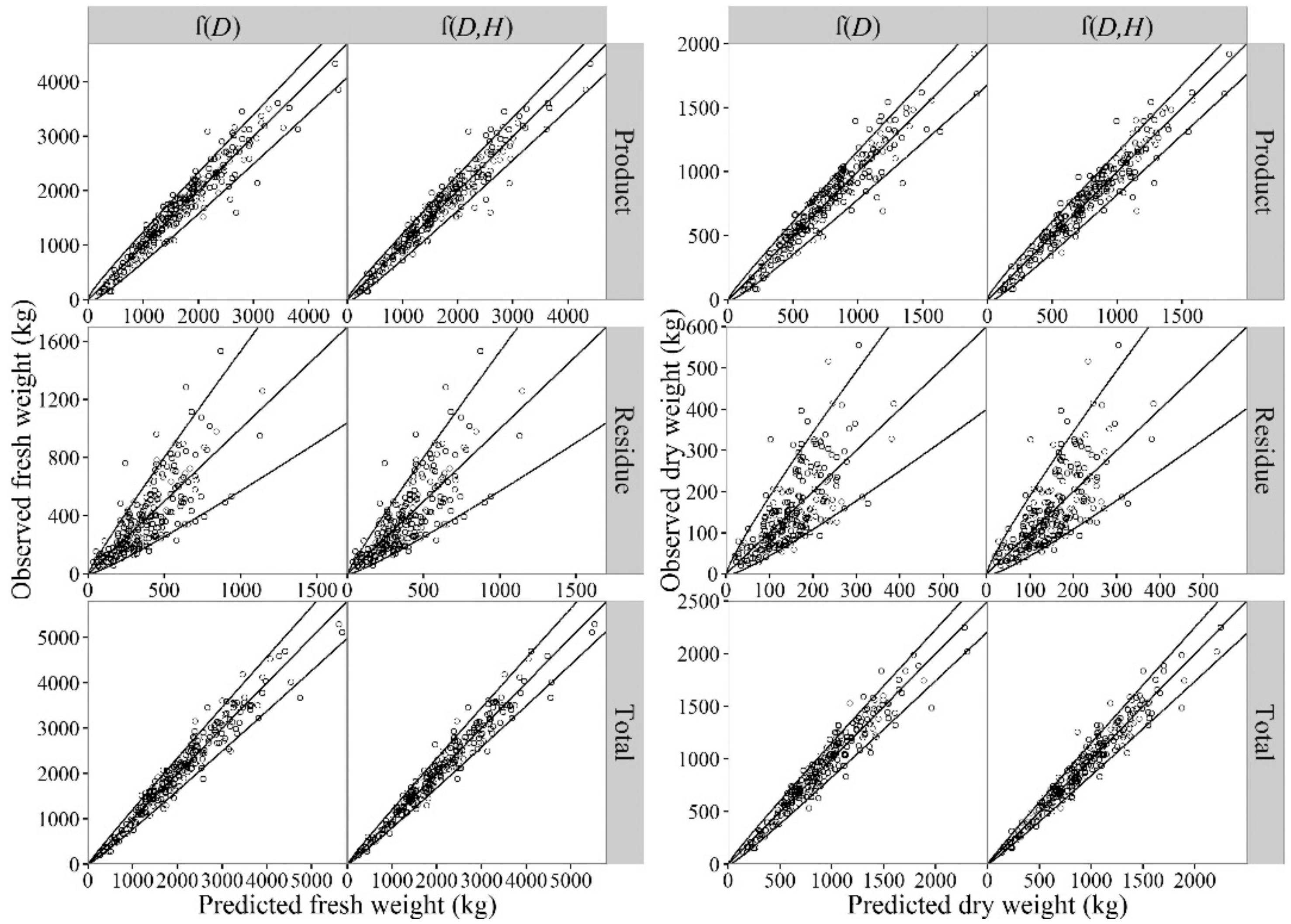
3.6. Prediction Accuracy
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Arnold, J.E.M.; Jongma, J. Fuelwood and charcoal in developing countries. South Asia 1977, 267, 0–38. [Google Scholar]
- Bhattacharya, S.C.; Salam, P.A. Low greenhouse gas biomass options for cooking in the developing countries. Biomass Bioenergy 2002, 22, 305–317. [Google Scholar] [CrossRef]
- Flinn, D.W. Comparison of establishment methods for Pinus radiata on a former P. pinaster site. Aust. For. 1978, 41, 167–176. [Google Scholar] [CrossRef]
- Pye, J.M.; Vitousek, P.M. Soil and nutrient removals by erosion and windrowing at a southeastern U.S. Piedmont site. For. Ecol. Manag. 1985, 11, 145–155. [Google Scholar] [CrossRef]
- Dyck, W.J.; Beets, P.N. Managing for long-term site productivity. N. Z. For. Nov. 1987, 32, 23–26. [Google Scholar]
- Creutzig, F.; Ravindranath, N.H.; Berndes, G.; Bolwig, S.; Bright, R.; Cherubini, F.; Chum, H.; Corbera, E.; Delucchi, M.; Faaij, A.; et al. Bioenergy and climate change mitigation: An assessment. Glob. Chang. Biol. Bioenergy 2015, 7, 916–944. [Google Scholar] [CrossRef] [Green Version]
- Röder, M.; Thornley, P. Bioenergy as climate change mitigation option within a 2 °C target—Uncertainties and temporal challenges of bioenergy systems. Energy Sustain. Soc. 2016, 6, 6–12. [Google Scholar] [CrossRef]
- Souza, G.M.; Ballester, M.V.R.; De Brito Cruz, C.H.; Chum, H.; Dale, B.; Dale, V.H.; Fernandes, E.C.M.; Foust, T.; Karp, A.; Lynd, L.; et al. The role of bioenergy in a climate-changing world. Environ. Dev. 2017, 23, 57–64. [Google Scholar] [CrossRef]
- Hoogwijk, M.; Faaij, A.; Van Den Broek, R.; Berndes, G.; Gielen, D.; Turkenburg, W. Exploration of the ranges of the global potential of biomass for energy. Biomass Bioenergy 2003, 25, 119–133. [Google Scholar] [CrossRef]
- Parikka, M. Global biomass fuel resources. Biomass Bioenergy 2004, 27, 613–620. [Google Scholar] [CrossRef]
- Mckechnie, J.; Colombo, S.; Chen, J.; Mabee, W.; Maclean, H.L. Forest bioenergy or forest carbon? Assessing trade-offs in greenhouse gas mitigation with wood-based fuels. Environ. Sci. Technol. 2011, 45, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Repo, A.; Känkänen, R.; Tuovinen, J.P.; Antikainen, R.; Tuomi, M.; Vanhala, P.; Liski, J. Forest bioenergy climate impact can be improved by allocating forest residue removal. Glob. Chang. Biol. Bioenergy 2012, 4, 202–212. [Google Scholar] [CrossRef]
- Gustavsson, L.; Haus, S.; Ortiz, C.A.; Sathre, R.; Truong, N.L. Climate effects of bioenergy from forest residues in comparison to fossil energy. Appl. Energy 2015, 138, 36–50. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, L.; Xie, G. Forest biomass energy resources in China: Quantity and distribution. Forests 2015, 6, 3970–3984. [Google Scholar] [CrossRef]
- Eisenbies, M.H.; Vance, E.D.; Aust, W.M.; Seiler, J.R. Intensive utilization of harvest residues in southern pine plantations: Quantities available and implications for nutrient budgets and sustainable site productivity. Bioenergy Res. 2009, 2, 90–98. [Google Scholar] [CrossRef]
- Kizha, A.R.; Han, H.-S. Forest residues recovered from whole-tree timber harvesting operations. Eur. J. For. Eng. 2015, 1, 46–55. [Google Scholar]
- Parresol, B.R. Assessing tree and stand biomass: A review with examples and critical comparisons. For. Sci. 1999, 45, 573–593. [Google Scholar]
- Fielding, J.M. The moisture content of the trunks of Monterey pine trees. Aust. For. 1952, 16, 3–21. [Google Scholar] [CrossRef]
- Moreno Chan, J.; Walker, J.C.F.; Raymond, C.A. Variation in green density and moisture content of radiata pine trees in the Hume region of New South Wales. Aust. For. 2012, 75, 31–42. [Google Scholar] [CrossRef]
- Perez-Garcia, J.; Lippke, B.; Comnick, J.; Manriquez, C. An assessment of carbon pools, storage, and wood products market substitution using life-cycle analysis results. Wood Fiber Sci. J. Soc. Wood Sci. Technol. 2005, 37, 140–148. [Google Scholar]
- Mckendry, P. Energy production from biomass. (Part 1): Overview of biomass. Bioresour. Technol. 2002, 83, 37–46. [Google Scholar] [CrossRef]
- Smith, C.T.; Lowe, A.T.; Skinner, M.F.; Beets, P.N.; Schoenholtz, S.H.; Fang, S. Response of radiata pine forests to residue management and fertilisation across a fertility gradient in New Zealand. For. Ecol. Manag. 2000, 138, 203–223. [Google Scholar] [CrossRef]
- Jones, H.S.; Beets, P.N.; Kimberley, M.O.; Garrett, L.G. Harvest residue management and fertilisation effects on soil carbon and nitrogen in a 15-year-old Pinus radiata plantation forest. For. Ecol. Manag. 2011, 262, 339–347. [Google Scholar] [CrossRef]
- Lavery, P.B.; Mead, D.J. Pinus radiata: A narrow endemic from North America takes on the world. In Ecology and Biogeography of Pinus; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Rogers, D.L. In situ genetic conservation of a naturally restricted and commercially widespread species, Pinus radiata. For. Ecol. Manag. 2004, 197, 311–322. [Google Scholar] [CrossRef]
- Rogers, D.L.; Matheson, A.C.; Vargas-Hernández, J.J.; Guerra-Santos, J.J. Genetic conservation of insular populations of Monterey pine (Pinus radiata D. Don). Biodivers. Conserv. 2006, 15, 779–798. [Google Scholar] [CrossRef]
- Mead, D.J. Sustainable Management of Pinus Radiata Plantations; Food and Agriculture Organization of the United Nations: Room, Italy, 2013. [Google Scholar]
- Lewis, N.B.; Ferguson, I.S.; Sutton, W.R.J.; Donald, D.G.M.; Lisboa, H.B. Management of Radiata Pine; Inkata Press Pty Ltd., Butterworth-Heinemann: Oxford, UK, 1993. [Google Scholar]
- Toro, J.; Gessel, S.P. Radiata pine plantations in Chile. New For. 1999, 18, 33–44. [Google Scholar] [CrossRef]
- Turner, J.; Gessel, S.P.; Lambert, M.J. Sustainable management of native and exotic plantations in Australia. New For. 1999, 18, 17–32. [Google Scholar] [CrossRef]
- Rogers, D.L. In Situ Genetic Conservation of Monterey Pine (Pinus Radiata D. Don): Information and Recommendations; Report 26; University of California Division of Agriculture and Natural Resources, Genetic Resources Conservation Program: Davis, CA, USA, 2002; pp. 1–80. [Google Scholar]
- Bi, H.; Simpson, J.; Li, R.; Yan, H.; Wu, Z.; Cai, S.; Eldridge, R. Introduction of Pinus radiata for afforestation: A review with reference to Aba, China. J. For. Res. 2003, 14, 311–322. [Google Scholar]
- Yan, H.; Bi, H.; Li, R.; Eldidge, R.; Wu, Z.; Li, Y.; Simpson, J. Assessing climatic suitability of Pinus radiata for summer rainfall environment of southwest China for ecological plantings. For. Ecol. Manag. 2006, 234, 199–208. [Google Scholar] [CrossRef]
- Bi, H.; Simpson, J.; Eldridge, R.; Sullivan, S.; Li, R.; Xiao, Y.; Zhou, J.; Wu, Z.; Yan, H.; Huang, Q.; et al. Survey of damaging pests and preliminary assessment of forest health risks to the long term success of Pinus radiata introduction in Sichuan, southwest China. J. For. Res. 2008, 19, 85–100. [Google Scholar] [CrossRef]
- Bi, H.; Li, R.; Wu, Z.; Huang, Q.; Liu, Q.; Zhou, Y.; Li, Y. Early performance of Pinus radiata provenances in the earthquake-ravaged dry river valley area of Sichuan, southwest China. J. For. Res. 2013, 24, 619–632. [Google Scholar] [CrossRef]
- Downham, R.; Gavran, M. Australian Plantation Statistics 2017 Update; Australian Government Department of Agriculture and Water Resources: Canberra, Australia, 2017; p. 12.
- Ferguson, I.S.; Leech, J.W. Generalized least squares estimation of yield functions. For. Sci. 1978, 24, 27–42. [Google Scholar]
- García, O. New class of growth models for even-aged stands: Pinus radiata in golden downs forest. N. Z. J. For. Sci. 1984, 14, 65–88. [Google Scholar]
- Horne, R.; Robinson, G.L. Development of Basal Area Thinning Prescriptions and Predictive Yield Models for Pinus Radiata Plantations in New South Wales, 1962–1988; Forestry Commission of New South Wales: Sydney, Australia, 1988. [Google Scholar]
- Candy, S.G. Growth and yield models for Pinus radiata in Tasmania. N. Z. J. For. Sci. 1989, 19, 112–133. [Google Scholar]
- Goulding, C.J. Development of growth models for Pinus radiata in New Zealand—Experience with management and process models. For. Ecol. Manag. 1994, 69, 331–343. [Google Scholar] [CrossRef]
- Castedo-Dorado, F.; Diéguez-Aranda, U.; Álvarez-González, J.G. A growth model for Pinus radiata D. Don stands in north-western Spain. Ann. For. Sci. 2007, 64, 453–465. [Google Scholar] [CrossRef]
- Guzmán, G.; Pukkala, T.; Palahí, M.; De-Miguel, S. Predicting the growth and yield of Pinus radiata in Bolivia. Ann. For. Sci. 2012, 69, 335–343. [Google Scholar] [CrossRef]
- Madgwick, H.A.I.; Webber, B. Nutrient removal in harvesting mature Pinus radiata. N. Z. J. For. 1987, 15–18. [Google Scholar]
- Ximenes, F.A.; Gardner, W.D.; Kathuria, A. Proportion of above-ground biomass in commercial logs and residues following the harvest of five commercial forest species in Australia. For. Ecol. Manag. 2008, 256, 335–346. [Google Scholar] [CrossRef]
- Cartes-Rodríguez, E.; Rubilar-Pons, R.; Acuña-Carmona, E.; Cancino-Cancino, J.; Rodríguez-Toro, J.; Burgos-Tornería, Y. Potential of Pinus radiata plantations for use of harvest residues in characteristic soils of south-central Chile. Revista Chapingo Serie Ciencias Forestales y del Ambiente 2016, 22, 221–233. [Google Scholar] [CrossRef]
- Bi, H.; Long, Y.; Turner, J.; Lei, Y.; Snowdon, P.; Li, Y.; Harper, R.; Zerihun, A.; Ximenes, F. Additive prediction of aboveground biomass for Pinus radiata (D. Don) plantations. For. Ecol. Manag. 2010, 259, 2301–2314. [Google Scholar] [CrossRef]
- Johnson, I.G.; Cotterill, I.M.; Raymond, C.A.; Henson, M. Half a century of Pinus radiata tree improvement in New South Wales. N. Z. J. For. 2008, 52, 7–13. [Google Scholar]
- Horne, R. The ‘Philosophy’ and Practice of P. Radiata Plantation Silviculture in New South Wales; Forestry Commission of NSW: Sydney, Australia, 1986. [Google Scholar]
- Raymond, C.A. Influence of prior land use on wood quality of Pinus radiata in New South Wales, Australia. For. Ecol. Manag. 2008, 255, 2626–2633. [Google Scholar] [CrossRef]
- Standards Association of Australia; Standards New Zealand. Timber—Methods of Test; Method 1: Moisture Content; AS/NZS 1080.1:1997; Standards Association of Australia: Sydney, Australia; Standards New Zealand: Wellington, New Zealand, 1997. [Google Scholar]
- Reuter, D.J.; Robinson, J.B.; Peverill, K.I.; Price, G.H.; Lambert, M.J. Guidelines for collecting, handling and analysing plant material. In Plant Analysis: An Interpretation Manual, 2nd ed.; Reuter, D.J., Robinson, J.B., Eds.; CSIRO Publishing: Clayton, Australia, 1997; pp. 53–80. [Google Scholar]
- Azur, M.J.; Stuart, E.A.; Frangakis, C.; Leaf, P.J. Multiple imputation by chained equations: What is it and how does it work? Int. J. Methods Psychiatr. Res. 2011, 20, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Van Buuren, S.; Groothuis-Oudshoorn, K. Mice: Multivariate imputation by chained equations in R. J. Stat. Softw. 2011, 45, 67. [Google Scholar] [CrossRef]
- Bi, H. Trigonometric variable-form taper equations for Australian eucalypts. For. Sci. 2000, 46, 397–409. [Google Scholar]
- Bi, H.; Long, Y. Flexible taper equation for site-specific management of Pinus radiata in New South Wales, Australia. For. Ecol. Manag. 2001, 148, 79–91. [Google Scholar] [CrossRef]
- Cribari-Neto, F.; Zeileis, A. Beta regression in R. J. Stat. Softw. 2010, 34, 24. [Google Scholar] [CrossRef]
- Paolino, P. Maximum likelihood estimation of models with beta-distributed dependent variables. Political Anal. 2001, 9, 325–346. [Google Scholar] [CrossRef]
- Ferrari, S.L.P.; Cribari-Neto, F. Beta regression for modelling rates and proportions. J. Appl. Stat. 2004, 31, 799–815. [Google Scholar] [CrossRef]
- Ramalho, E.A.; Ramalho, J.J.S.; Murteira, J.M.R. Alternative estimating and testing empirical strategies for fractional regression models. J. Econ. Surv. 2011, 25, 19–68. [Google Scholar] [CrossRef]
- Schmid, M.; Wickler, F.; Maloney, K.O.; Mitchell, R.; Fenske, N.; Mayr, A. Boosted beta regression. PLoS ONE 2013, 8, e61623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cepeda-Cuervo, E. Beta regression models: Joint mean and variance modeling. J. Stat. Theory Pract. 2015, 9, 134–145. [Google Scholar] [CrossRef]
- Barreto-Souza, W.; Simas, A.B. Improving estimation for beta regression models via em-algorithm and related diagnostic tools. J. Stat. Comput. Simul. 2017, 87, 2847–2867. [Google Scholar] [CrossRef]
- Parresol, B.R. Additivity of nonlinear biomass equations. Can. J. For. Res. 2001, 31, 865–878. [Google Scholar] [CrossRef]
- Bi, H.; Turner, J.; Lambert, M.J. Additive biomass equations for native eucalypt forest trees of temperate Australia. Trees 2004, 18, 467–479. [Google Scholar] [CrossRef]
- Bi, H.; Murphy, S.; Volkova, L.; Weston, C.; Fairman, T.; Li, Y.; Law, R.; Norris, J.; Lei, X.; Caccamo, G. Additive biomass equations based on complete weighing of sample trees for open eucalypt forest species in south-eastern Australia. For. Ecol. Manag. 2015, 349, 106–121. [Google Scholar] [CrossRef]
- Zeng, W.S.; Zhang, H.R.; Tang, S.Z. Using the dummy variable model approach to construct compatible single-tree biomass equations at different scales—A case study for Masson pine (Pinus massoniana) in southern China. Can. J. For. Res. 2011, 41, 1547–1554. [Google Scholar] [CrossRef]
- Fu, L.; Zeng, W.; Tang, S.; Sharma, R.P.; Li, H. Using linear mixed model and dummy variable model approaches to construct compatible single-tree biomass equations at different scales—A case study for Masson pine in southern China. J. For. Sci. 2012, 58, 101–115. [Google Scholar]
- Fu, L.; Lei, Y.; Sun, W.; Tang, S.; Zeng, W. Development of compatible biomass models for trees from different stand origin. Acta Ecol. Sin. 2014, 34, 1461–1470. [Google Scholar]
- Greene, W.H. Econometric Analysis: Fourth Edition; Prentice Hall: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- Tang, S.; Zhang, H.; Xu, H. Study on establish and estimate method of compatible biomass model. Sci. Silvae Sin. 2000, 36, 19–27. [Google Scholar]
- Tang, S.; Li, Y.; Wang, Y. Simultaneous equations, error-in-variable models, and model integration in systems ecology. Ecol. Model. 2001, 142, 285–294. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, L.; Li, F. A three-step proportional weighting system of nonlinear biomass equations. For. Sci. 2015, 61, 35–45. [Google Scholar] [CrossRef]
- Tang, S.; Lang, K.; Li, H. Statistics and Computation of Biomathematical Models (ForStat Course); Science Press: Beijing, China, 2008. [Google Scholar]
- Amemiya, T. Advanced Econometrics; Harvard University Press: Cambridge, MA, USA, 1985; p. 536. [Google Scholar]
- Snowdon, P. A ratio estimator for bias correction in logarithmic regressions. Can. J. For. Res. 1991, 21, 720–724. [Google Scholar] [CrossRef]
- Huang, S.; Yang, Y.; Wang, Y.; Amaro, A.; Reed, D.; Soares, P. A Critical Look at Procedures for Validating Growth and Yield Models. In Modelling Forest Systems. Workshop on the Interface between Reality, Modelling and the Parameter Estimation Processes, Sesimbra, Portugal, 2–5 June 2002; CABI Publishing: Wallingford, UK, 2003; p. 293. [Google Scholar]
- Chalk, L.; Bigg, J.M. The distribution of moisture in the living stem in Sitka Spruce and Douglas fir. For. Int. J. For. Res. 1956, 29, 5–21. [Google Scholar] [CrossRef]
- Stewart, C.M. Moisture content of living trees. Nature 1967, 214, 138–140. [Google Scholar] [CrossRef]
- Nakada, R. Within-stem water distribution in living trees of some conifers. IAWA J. 2006, 27, 313–327. [Google Scholar]
- Paul, K.I.; Roxburgh, S.H.; Larmour, J.S. Moisture content correction: Implications of measurement errors on tree- and site-based estimates of biomass. For. Ecol. Manag. 2017, 392, 164–175. [Google Scholar] [CrossRef]
- Moreno Chan, J. Moisture Content in Radiata Pine Wood: Implications for Wood Quality and Water-Stress Response; University of Canterbury: Christchurch, New Zealand, 2007. [Google Scholar]
- Tian, X.; Cown, D.J.; McConchie, D.L. Modelling of Pinus radiata wood properties—Part 2: Basic density. N. Z. J. For. Sci. 1995, 25, 214–230. [Google Scholar]
- Madgwick, H.A.I. Pinus Radiata—Biomass, Form and Growth; Madgwick: Rotorua, New Zealand, 1994. [Google Scholar]
- Young, G.D.; McConchie, D.L.; McKinley, R.B. Utilisation of 25-year-old Pinus radiata—Part 1: Wood properties. N. Z. J. For. Sci. 1991, 21, 217–227. [Google Scholar]
- Murphy, G.; Cown, D. Within-tree, between-tree, and geospatial variation in estimated Pinus radiata bark volume and weight in New Zealand. N. Z. J. For. Sci. 2015, 45, 18. [Google Scholar] [CrossRef]
- Biørn, E. Regression systems for unbalanced panel data: A stepwise maximum likelihood procedure. J. Econ. 2004, 122, 281–291. [Google Scholar] [CrossRef]
- Biørn, E. Estimating SUR system with random coefficients: The unbalanced panel data case. Empir. Econ. 2014, 47, 451–468. [Google Scholar] [CrossRef]
- Lu, K.; Bi, H.; Watt, D.; Strandgard, M.; Li, Y. Reconstructing the size of individual trees using log data from cut-to-length harvesters in Pinus radiata plantations: A case study in NSW, Australia. J. For. Res. 2017, 1–21. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Thinning Type | Number of Plots | Age (Year) | Basal Area (m2/ha) | Dominant Height (m) | Stand Density (trees/ha) |
|---|---|---|---|---|---|
| T0 | 30 | 33–42 (39) | 50.0–89.2 (64.4) | 26.6–38.5 (31.5) | 424–1188 (740) |
| T1 | 15 | 28–29 (28) | 40.8–52.8 (46.8) | 29.4–34.1 (31.8) | 199–477 (325) |
| T2 | 15 | 28–33 (32) | 30.5–46.6 (38.3) | 29.3–37.3 (32.6) | 163–366 (220) |
| Stemwood | Bark | ||||
|---|---|---|---|---|---|
| Parameter | Estimate | SE | Parameter | Estimate | SE |
| w1 | 0.4497 | 0.2009 | b1 | 1.1161 | 0.3867 |
| w2 | 0.1063 | 0.0595 | b2 | 0.0696 | 0.0438 |
| w3 | 0.4290 | 0.0607 | b3 | 0.4982 | 0.0584 |
| w4 | 0.5233 | 0.0540 | b4 | −1.6781 | 0.0591 |
| w5 | −0.0054 | 0.0759 | b5 | −0.1512 | 0.1008 |
| w6 | −0.2514 | 0.0785 | |||
| w7 | −0.3031 | 0.0513 | |||
| pseudo-R2 | 0.74 | pseudo-R2 | 0.88 | ||
| Parameter | Estimate | SE | Pseudo-R2 |
|---|---|---|---|
| a | −1.1667 | 0.7349 | 0.73 |
| b | −3.3327 | 0.4540 | |
| c | 3.6565 | 0.5137 | |
| d | −0.1374 | 0.1981 |
| Tree | Parameter | Fresh Weight | Dry Weight | ||||
|---|---|---|---|---|---|---|---|
| Component | Estimate | SE | R2 | Estimate | SE | R2 | |
| Equation (6), , | |||||||
| Product () | 0.2587 | 0.0283 | 0.92 | 0.2761 | 0.0455 | 0.90 | |
| 2.2997 | 0.0288 | 2.0817 | 0.0433 | ||||
| Residue ( | 0.0255 | 0.0122 | 0.60 | 0.0386 | 0.0191 | 0.51 | |
| 2.5203 | 0.1257 | 2.1674 | 0.1295 | ||||
| Total () | 0.94 | 0.93 | |||||
| Equation (8), , | |||||||
| Product () | 0.0370 | 0.0117 | 0.93 | 0.0378 | 0.0103 | 0.92 | |
| 2.1122 | 0.0400 | 1.8930 | 0.0505 | ||||
| 0.7740 | 0.1086 | 0.7880 | 0.1012 | ||||
| Residue ( | 0.0248 | 0.0103 | 0.60 | 0.0372 | 0.0169 | 0.51 | |
| 2.5275 | 0.1077 | 2.1755 | 0.1190 | ||||
| Total () | 0.95 | 0.94 | |||||
| Equation (9), , | |||||||
| Product () | 0.3543 | 0.0406 | 0.93 | 0.2535 | 0.0443 | 0.92 | |
| 2.2140 | 0.0297 | 2.0922 | 0.0461 | ||||
| 0.0148 | 0.0034 | 0.0223 | 0.0042 | ||||
| −0.0175 | 0.0062 | 0.0171 | 0.0051 | ||||
| Residue ( | 0.0310 | 0.0163 | 0.62 | 0.0392 | 0.0176 | 0.51 | |
| 2.4914 | 0.1353 | 2.1606 | 0.1163 | ||||
| −0.0359 | 0.0136 | ||||||
| −0.0404 | 0.0242 | ||||||
| Total () | 0.95 | 0.94 | |||||
| Equation (10), , | |||||||
| Product () | 0.0516 | 0.0168 | 0.94 | 0.0326 | 0.0086 | 0.93 | |
| 2.0562 | 0.0449 | 1.9163 | 0.0495 | ||||
| 0.7339 | 0.1198 | 0.7900 | 0.0958 | ||||
| 0.0148 | 0.0036 | 0.0228 | 0.0037 | ||||
| −0.0114 | 0.0068 | 0.0262 | 0.0051 | ||||
| Residue ( | 0.0295 | 0.0136 | 0.62 | 0.0374 | 0.0148 | 0.51 | |
| 2.5040 | 0.1186 | 2.1722 | 0.1019 | ||||
| −0.0359 | 0.0123 | ||||||
| −0.0399 | 0.0241 | ||||||
| Total () | 0.95 | 0.95 | |||||
| Tree Component | Predictor | p5 | p95 | |||
|---|---|---|---|---|---|---|
| b | ||||||
| Fresh weight | ||||||
| D | 15.3344 | 0.8764 | 5.1923 | −1.6947 | 1.3512 | |
| Residue | D | 0.3872 | 1.6312 | 3.5267 | −1.3098 | 1.6301 |
| Total | D | 0.1950 | 1.4607 | 3.9601 | −1.6560 | 1.5318 |
| Dry weight | ||||||
| Product | D | 1.3190 | 1.1296 | 4.7699 | −1.7473 | 1.3031 |
| Residue | D | 0.9646 | 1.4208 | 3.4076 | −1.1757 | 1.8415 |
| Total | D | 1.1391 | 1.1534 | 3.8866 | −1.5161 | 1.3727 |
| Fresh weight | ||||||
| Product | D, H | 10.7596 | 0.9171 | 4.5895 | −1.5781 | 1.2667 |
| Residue | D, H | 0.4047 | 1.6220 | 3.5703 | −1.3150 | 1.6298 |
| Total | D, H | 0.2604 | 1.4009 | 4.0150 | −1.4839 | 1.6155 |
| Dry weight | ||||||
| Product | D, H | 7.6822 | 0.8563 | 4.3787 | −1.5936 | 1.3505 |
| Residue | D, H | 1.0377 | 1.4068 | 3.4324 | −1.1673 | 1.8546 |
| Total | D, H | 0.0919 | 1.4749 | 4.2767 | −1.5239 | 1.4914 |
| Tree Component | Stand | θs | p5 | p95 | ||
|---|---|---|---|---|---|---|
| Type | b | |||||
| Fresh weight | ||||||
| Product | T0 | 6.7815 | 0.9458 | 4.4141 | −1.6618 | 1.5271 |
| T1 | 9.7562 | 0.9458 | 5.6492 | −1.5033 | 1.3586 | |
| T2 | 7.3155 | 0.9458 | 4.6826 | −1.3650 | 1.5189 | |
| Residue | T0 | 0.3675 | 1.7025 | 2.6791 | −1.0969 | 1.6872 |
| T1 | 0.1773 | 1.7025 | 4.3380 | −1.3152 | 1.6022 | |
| T2 | 0.2290 | 1.7025 | 3.7288 | −1.2699 | 1.8052 | |
| Total | T0 | 0.0588 | 1.6768 | 2.3100 | −1.8942 | 1.3194 |
| T1 | 0.0475 | 1.6768 | 3.1247 | −1.4882 | 1.4235 | |
| T2 | 0.0328 | 1.6768 | 3.6697 | −1.8579 | 1.7139 | |
| Dry weight | ||||||
| Product | T0 | 3.4018 | 0.9605 | 3.6655 | −1.8418 | 1.3759 |
| T1 | 3.8532 | 0.9605 | 5.5436 | −1.6111 | 1.1995 | |
| T2 | 3.1352 | 0.9605 | 4.1343 | −1.2882 | 1.6791 | |
| Residue | T0, T1, T2 | 1.2607 | 1.3621 | 3.5896 | −1.1511 | 1.8664 |
| Total | T0 | 0.1218 | 1.5100 | 2.5433 | −1.9707 | 1.3969 |
| T1 | 0.1134 | 1.5100 | 3.0902 | −1.4408 | 1.4230 | |
| T2 | 0.0760 | 1.5100 | 3.8888 | −1.7484 | 1.6971 | |
| Tree Component | Stand | p5 | p95 | |||
|---|---|---|---|---|---|---|
| Type | b | |||||
| Fresh weight | ||||||
| Product | T0 | 3.3324 | 1.1226 | 3.3611 | −1.6399 | 1.2789 |
| T1 | 1.9472 | 1.1226 | 5.6202 | −1.7525 | 1.2699 | |
| T2 | 1.8473 | 1.1226 | 4.0394 | −1.4824 | 1.4159 | |
| Residue | T0 | 0.4529 | 1.6645 | 2.6948 | −1.0964 | 1.6950 |
| T1 | 0.2260 | 1.6645 | 4.3267 | −1.3056 | 1.5984 | |
| T2 | 0.2765 | 1.6645 | 3.9167 | −1.2788 | 1.8020 | |
| Total | T0 | 0.0042 | 2.0262 | 3.0365 | −1.8946 | 1.3323 |
| T1 | 0.0023 | 2.0262 | 3.2479 | −1.8056 | 1.6948 | |
| T2 | 0.0021 | 2.0262 | 2.9956 | −1.5580 | 1.6932 | |
| Dry weight | ||||||
| Product | T0 | 5.7190 | 0.9349 | 3.5573 | −1.8767 | 1.0616 |
| T1 | 3.0913 | 0.9349 | 5.9595 | −1.6808 | 1.1194 | |
| T2 | 3.5089 | 0.9349 | 3.5960 | −1.3641 | 1.5202 | |
| Residue | T0, T1, T2 | 1.2744 | 1.3586 | 3.6255 | −1.1495 | 1.8714 |
| Total | T0 | 0.0078 | 1.9280 | 2.9966 | −2.0387 | 1.3830 |
| T1 | 0.0040 | 1.9280 | 3.4928 | −1.8257 | 1.6743 | |
| T2 | 0.0035 | 1.9280 | 3.3492 | −1.4458 | 1.6745 | |

| DBHOB (cm) | Product | Residue | |||
|---|---|---|---|---|---|
| Sawlog | Pulpwood | Stump | Branch | Waste | |
| Fresh weight | |||||
| 15 | 0.8283 | 0.1717 | 0.1074 | 0.1797 | 0.7129 |
| 20 | 0.8329 | 0.1671 | 0.1112 | 0.2467 | 0.6421 |
| 25 | 0.8363 | 0.1637 | 0.1117 | 0.3088 | 0.5794 |
| 30 | 0.8391 | 0.1609 | 0.1104 | 0.3651 | 0.5244 |
| 35 | 0.8414 | 0.1586 | 0.1081 | 0.4157 | 0.4763 |
| 40 | 0.8434 | 0.1566 | 0.1051 | 0.4608 | 0.4341 |
| 45 | 0.8452 | 0.1548 | 0.1017 | 0.5011 | 0.3972 |
| 50 | 0.8467 | 0.1533 | 0.0983 | 0.5370 | 0.3647 |
| 55 | 0.8481 | 0.1519 | 0.0949 | 0.5690 | 0.3361 |
| 60 | 0.8494 | 0.1506 | 0.0915 | 0.5978 | 0.3108 |
| 65 | 0.8505 | 0.1495 | 0.0882 | 0.6235 | 0.2883 |
| 70 | 0.8516 | 0.1484 | 0.0851 | 0.6467 | 0.2682 |
| Dry weight | |||||
| 15 | 0.8209 | 0.1791 | 0.1225 | 0.1758 | 0.7017 |
| 20 | 0.8313 | 0.1687 | 0.1288 | 0.2314 | 0.6398 |
| 25 | 0.8390 | 0.1610 | 0.1318 | 0.2820 | 0.5862 |
| 30 | 0.8450 | 0.1550 | 0.1328 | 0.3276 | 0.5396 |
| 35 | 0.8500 | 0.1500 | 0.1325 | 0.3687 | 0.4988 |
| 40 | 0.8542 | 0.1458 | 0.1313 | 0.4058 | 0.4629 |
| 45 | 0.8579 | 0.1421 | 0.1297 | 0.4393 | 0.4311 |
| 50 | 0.8611 | 0.1389 | 0.1276 | 0.4696 | 0.4028 |
| 55 | 0.8639 | 0.1361 | 0.1254 | 0.4971 | 0.3775 |
| 60 | 0.8664 | 0.1336 | 0.1231 | 0.5221 | 0.3548 |
| 65 | 0.8687 | 0.1313 | 0.1207 | 0.5450 | 0.3344 |
| 70 | 0.8708 | 0.1292 | 0.1182 | 0.5659 | 0.3158 |
| Tree | Fresh Weight | Dry Weight | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Component | MEP (kg) | MPEP (%) | MAEP (kg) | MPAEP (%) | MSEP | MEP (kg) | MPEP (%) | MAEP (kg) | MPAEP (%) | MSEP | ||
| Equation (6), one-variable model, without dummy variable | ||||||||||||
| Product | −2.4 | −1.5 | 167.7 | 11 | 57,964 | 0.91 | −0.3 | −1.3 | 79.7 | 11 | 13,146 | 0.89 |
| Residue | 0.1 | 1.8 | 120.6 | 33 | 27,043 | 0.58 | 0.1 | 1.8 | 49.4 | 35 | 4439 | 0.49 |
| Total | −2.3 | −1.1 | 181.0 | 9 | 61,220 | 0.94 | −0.2 | −0.9 | 82.5 | 10 | 12,908 | 0.92 |
| Sawlog | 9.6 | −1.8 | 216.4 | 18 | 83,420 | 0.84 | 4.2 | −1.5 | 98.9 | 17 | 17,690 | 0.82 |
| Pulpwood | 4.4 | 6.2 | 140.6 | 58 | 41,820 | 0.25 | 2.1 | 6.0 | 62.9 | 61 | 8269 | 0.18 |
| Stump | −0.3 | −1.9 | 9.0 | 24 | 188 | 0.60 | −0.1 | −1.2 | 4.6 | 25 | 46 | 0.57 |
| Branch | −3.4 | 1.0 | 63.7 | 33 | 8529 | 0.63 | −1.4 | 1.3 | 20.5 | 32 | 838 | 0.62 |
| Waste | 3.8 | 4.6 | 100.1 | 68 | 18,990 | 0.13 | 1.6 | 4.3 | 44.1 | 71 | 3645 | 0.09 |
| Equation (8), two-variable model, without dummy variable | ||||||||||||
| Product | −1.5 | −1.1 | 155.8 | 10 | 48,356 | 0.93 | 0.0 | −0.8 | 74.9 | 11 | 11,005 | 0.91 |
| Residue | −0.5 | 1.8 | 120.5 | 33 | 27,000 | 0.58 | 0.6 | 2.2 | 49.3 | 35 | 4435 | 0.49 |
| Total | −2.1 | −0.6 | 166.7 | 9 | 51,909 | 0.95 | 0.6 | −0.3 | 74.9 | 9 | 10,724 | 0.94 |
| Sawlog | 10.4 | −1.5 | 203.7 | 17 | 75,537 | 0.85 | 4.4 | −1.2 | 93.0 | 16 | 15,845 | 0.84 |
| Pulpwood | 4.7 | 8.0 | 142.3 | 60 | 41,976 | 0.25 | 2.2 | 7.9 | 63.7 | 63 | 8315 | 0.18 |
| Stump | −0.4 | −1. 9 | 9.0 | 24 | 188 | 0.60 | −0.1 | −0.8 | 4.5 | 25 | 46 | 0.57 |
| Branch | −3.8 | 1.0 | 63.7 | 33 | 8511 | 0.64 | −1.1 | 1.7 | 20.4 | 32 | 836 | 0.62 |
| Waste | 3.6 | 4.6 | 100.1 | 68 | 18,986 | 0.13 | 1.8 | 4.7 | 44.1 | 72 | 3645 | 0.09 |
| Equation (9), one-variable model, with dummy variables | ||||||||||||
| Product | −2.3 | −1.3 | 163.0 | 11 | 53,112 | 0.92 | −2.8 | −1.4 | 74.4 | 11 | 10,924 | 0.91 |
| Residue | −0.1 | 2.2 | 118.9 | 33 | 26,172 | 0.60 | 1.3 | 2.6 | 49.3 | 35 | 4466 | 0.49 |
| Total | −2.4 | −0.9 | 179.6 | 9 | 60,194 | 0.94 | −1.5 | −0.9 | 79.5 | 9 | 11,409 | 0.93 |
| Sawlog | 10.6 | −1.5 | 208.1 | 17 | 78,969 | 0.84 | 2.5 | −1.6 | 95.8 | 17 | 16,226 | 0.83 |
| Pulpwood | 4.7 | 6.4 | 140.3 | 58 | 41,393 | 0.26 | 1.8 | 6.0 | 62.7 | 61 | 8164 | 0.19 |
| Stump | −0.3 | −1.3 | 9.4 | 25 | 197 | 0.59 | −0.1 | −1.4 | 4.6 | 25 | 46 | 0.57 |
| Branch | −3.3 | 0.9 | 60.5 | 33 | 7388 | 0.68 | 0.1 | 2.5 | 20.3 | 32 | 832 | 0.63 |
| Waste | 3.6 | 5.6 | 101.6 | 69 | 19,361 | 0.11 | 1.3 | 4 | 44.2 | 72 | 3636 | 0.09 |
| Equation (10), two-variable model, with dummy variables | ||||||||||||
| Product | −3.7 | −1.0 | 152.2 | 10 | 45,046 | 0.93 | −3.9 | −1.2 | 68.4 | 10 | 9036 | 0.92 |
| Residue | 0.6 | 2.6 | 118.8 | 33 | 26,159 | 0.60 | 1.9 | 3.1 | 49.2 | 35 | 4462 | 0.49 |
| Total | −3.1 | −0.6 | 166.2 | 9 | 51,575 | 0.95 | −1.9 | −0.6 | 71.6 | 8 | 9395 | 0.94 |
| Sawlog | 9.5 | −1.3 | 199.5 | 17 | 73,535 | 0.85 | 1.6 | −1.6 | 90.8 | 16 | 14,813 | 0.85 |
| Pulpwood | 4.6 | 8.0 | 141.5 | 60 | 41,439 | 0.26 | 1.7 | 7.4 | 63.5 | 63 | 8198 | 0.19 |
| Stump | −0.3 | −1.0 | 9.4 | 25 | 197 | 0.59 | 0.0 | −1.0 | 4.6 | 25 | 46 | 0.57 |
| Branch | −3.0 | 1.3 | 60.4 | 33 | 7366 | 0.68 | 0.4 | 3.0 | 20.3 | 32 | 831 | 0.63 |
| Waste | 3.9 | 6.0 | 101.5 | 70 | 19,368 | 0.11 | 1.6 | 4.8 | 44.2 | 72 | 3635 | 0.09 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Bi, H.; Ximenes, F.; Ramos, J.; Li, Y. Product and Residue Biomass Equations for Individual Trees in Rotation Age Pinus radiata Stands under Three Thinning Regimes in New South Wales, Australia. Forests 2017, 8, 439. https://doi.org/10.3390/f8110439
Wang X, Bi H, Ximenes F, Ramos J, Li Y. Product and Residue Biomass Equations for Individual Trees in Rotation Age Pinus radiata Stands under Three Thinning Regimes in New South Wales, Australia. Forests. 2017; 8(11):439. https://doi.org/10.3390/f8110439
Chicago/Turabian StyleWang, Xin, Huiquan Bi, Fabiano Ximenes, Jorge Ramos, and Yun Li. 2017. "Product and Residue Biomass Equations for Individual Trees in Rotation Age Pinus radiata Stands under Three Thinning Regimes in New South Wales, Australia" Forests 8, no. 11: 439. https://doi.org/10.3390/f8110439
APA StyleWang, X., Bi, H., Ximenes, F., Ramos, J., & Li, Y. (2017). Product and Residue Biomass Equations for Individual Trees in Rotation Age Pinus radiata Stands under Three Thinning Regimes in New South Wales, Australia. Forests, 8(11), 439. https://doi.org/10.3390/f8110439