Identification of Floral Relicts Based on Spatial Distance of Isolation

Abstract
:1. Introduction
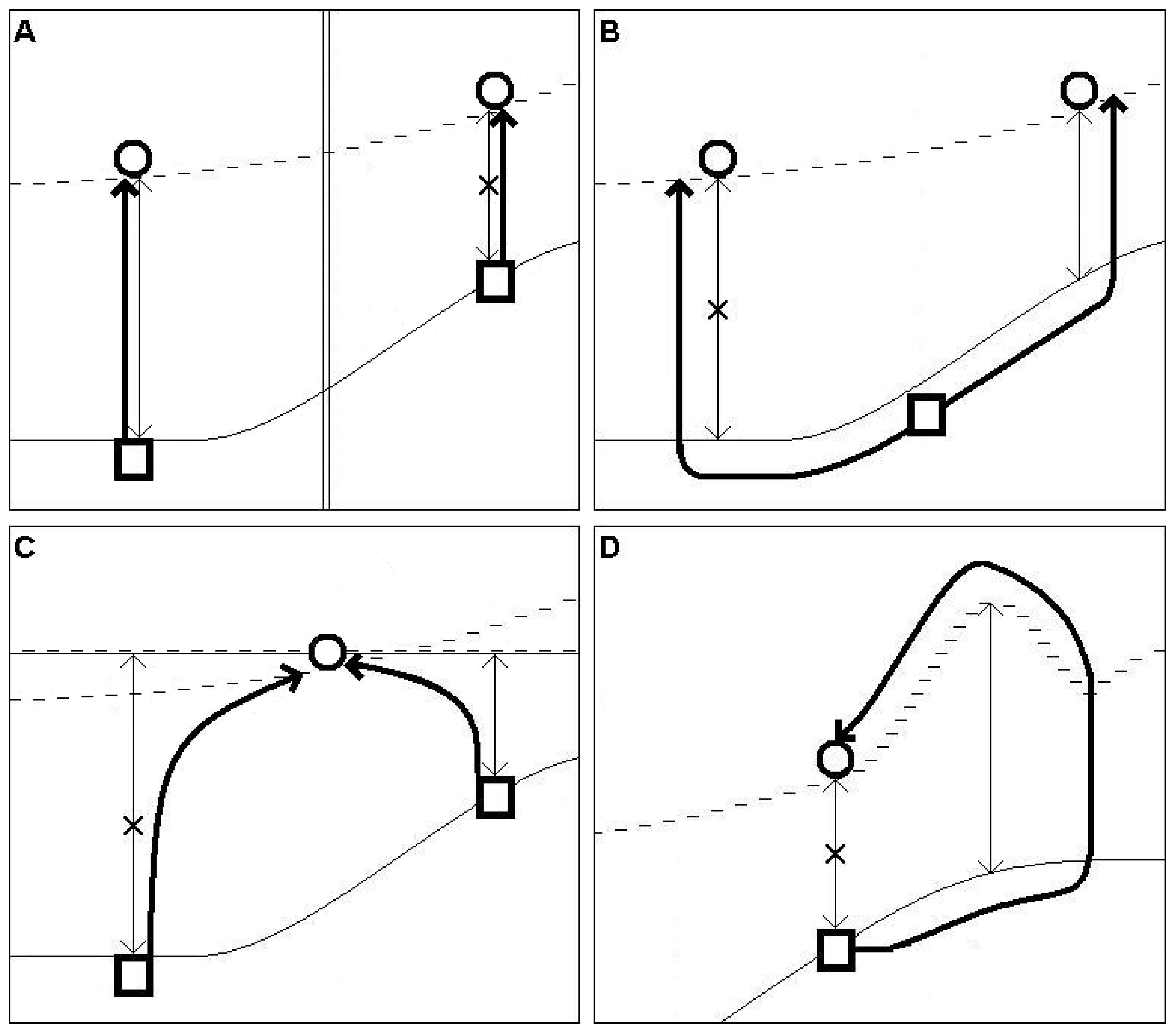
- An inner position in the continuous distribution of a species does not imply relict occurrence.
- An edge position on a continuous distribution can imply relict occurrence, although with a very low probability.
- An isolated occurrence from a large, continuous distribution but close to it can be regarded as a relict with a slightly higher probability.
- An isolated occurrence far from a large, continuous distribution can be regarded as a relict occurrence with much a higher probability than in the former cases.
- (1)
- Which species, as candidates for the status of an HTM (or postglacial) relict, satisfy the criteria of N-SD of 150–350 km in the Pannonian flora? We expect a limited number of such species.
- (2)
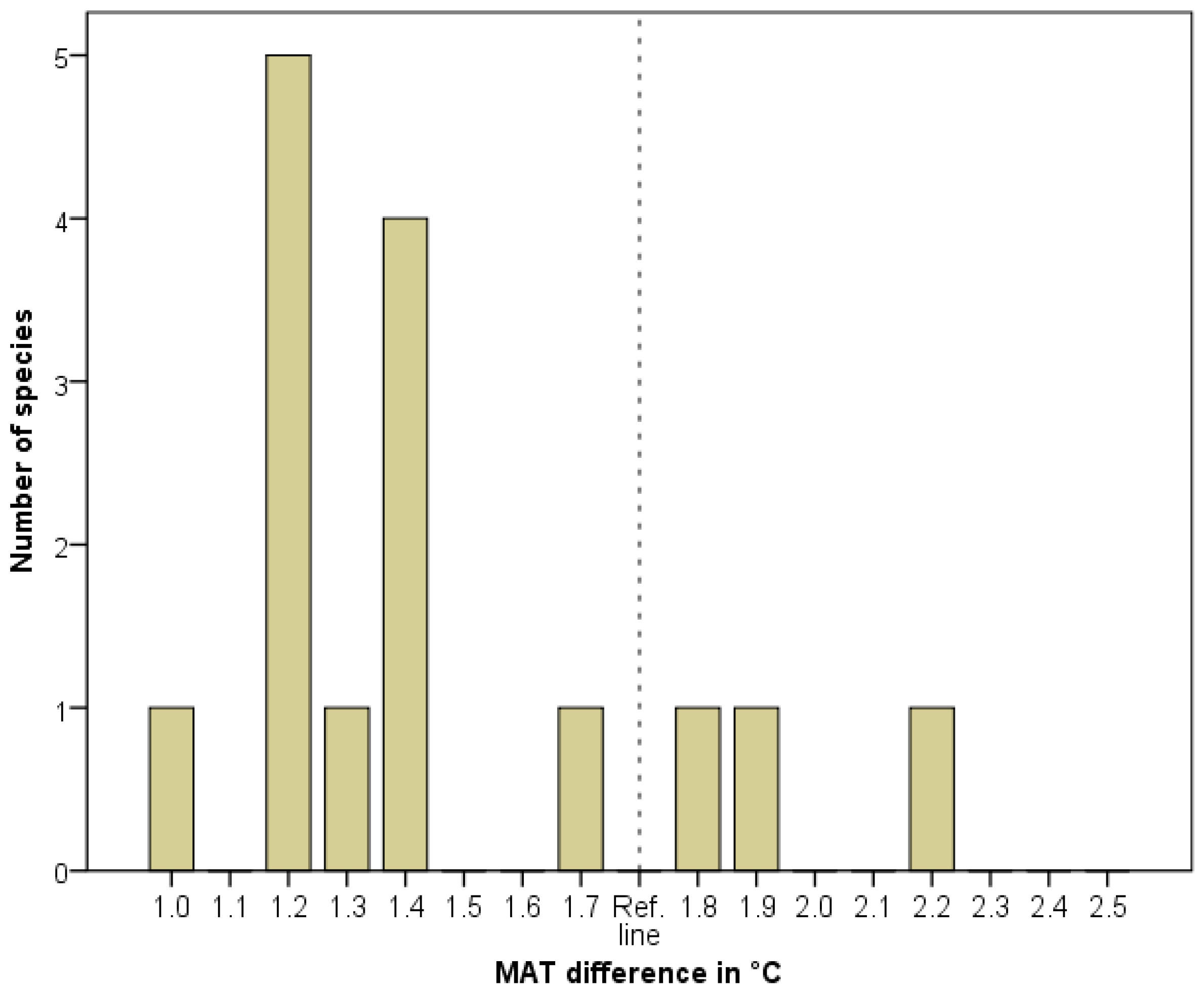
- How is MATD distributed among these taxa? The distribution of N-SD values, if they are not evenly distributed, can be informative with respect to assessing the exact value of MATD within the wider range (of 1.0–2.5 °C) or with regard to judging whether some relicts originated from earlier interglacial periods.
2. Materials and Methods
- The HTM-expansion was not a rapid process (anemochory and zoochory are excluded);
- The area of the gap is flat or hilly, where micro- and mesoclimatic effects are weak, or, if the investigated species depends on habitats influenced by the micro- and mesoclimate, then their effect is evenly distributed;
- The disjunction is not the result of human impacts and re-occupation was not limited by anthropogenic influences.
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hampe, A.; Jump, A.S. Climate relicts: Past, present, future. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 313–333. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.H.; Lomolino, M.V. Biogeography; Cambridge University Press: Sunderland, MA, USA, 1998; p. 300. [Google Scholar]
- Croizat, L. Manual of Phytogeography; Springer-Science + Business Media: Amsterdam, The Netherlands, 1952. [Google Scholar]
- Cox, C.B.; Moore, P.D.; Marquardt, W.C.; Demaree, R.S.; Grieve, R.B. Biogeography: An Ecological and Evolutionary Approach, 6th ed.; Blackwell Publishing: Hoboken, NJ, USA, 2005. [Google Scholar]
- Habel, J.C.; Assmann, T. (Eds.) Relict Species: Phylogeography and Conservation Biology; Springer Science & Business Media: New York, NY, USA, 2009; p. 324. [Google Scholar]
- Hájková, P.; Horsák, M.; Hájek, M.; Jankovská, V.; Jamrichová, E.; Moutelíková, J. Using multi-proxy palaeoecology to test a relict status of refugial populations of calcareous-fen species in the Western Carpathians. Holocene 2015, 25, 702–715. [Google Scholar] [CrossRef]
- Huggett, R.J. Fundamentals of Biogeography, 2nd ed.; Routledge: Abingdon-on-Thames, UK, 2004; pp. 62–63. [Google Scholar]
- Soó, R. The history of the vegetation of Hungary. Collect. Bot. 1968, 7, 1109–1120. [Google Scholar]
- Zahariev, D. Biodiversity of Relict Vascular Plants in Bulgaria. Int. J. Res. Stud. Biosci. 2016, 4, 38–51. [Google Scholar]
- Kaplan, Z. Flora and phytogeography of the Czech Republic. Dedicated to the centenary of the Czech Botanical Society (1912–2012). Preslia 2012, 84, 505–573. [Google Scholar]
- Kaplan, Z.; Danihelka, J.; Štěpánková, J.; Bureš, P.; Zázvorka, J.; Hroudová, Z.; Ducháček, M.; Grulich, V.; Řepka, R.; Dančáket, M.; et al. Distributions of vascular plants in the Czech Republic. Part 1. Preslia 2015, 87, 417–500. [Google Scholar]
- Bartha, D.; Király, G. (Eds.) Distribution Atlas of Vascular Plants of Hungary; University of West Hungary Press: Sopron, Hungary, 2015; pp. 1–329. [Google Scholar]
- Futák, J.; Bertová, L. Flóra Slovenska III; VEDA, Vydavateľstvo Slovenskej Akadémie Vied: Bratislava, Slovakia, 1982; p. 519. [Google Scholar]
- Goliašová, K.; Šípošová, H. (Eds.) Flóra Slovenska VI/1; VEDA Vydavateľstvo Slovenskej Akadémie Vied: Bratislava, Slovakia, 2008; p. 170. [Google Scholar]
- Rull, V. Microrefugia. J. Biogeogr. 2009, 36, 481–484. [Google Scholar] [CrossRef]
- Birks, H.J.B. Some reflections on the refugium concept and its terminology in historical biogeography, contemporary ecology and global-change biology. Biodiversity 2015, 16, 196–212. [Google Scholar] [CrossRef]
- Hájková, P.; Roleček, J.; Hájek, M.; Horsák, M.; Fajmon, K.; Polák, M.; Jamrichová, E. Prehistoric origin of the extremely species-rich semi-dry grasslands in the Bílé Karpaty Mts (Czech Republic and Slovakia). Preslia 2011, 83, 185–204. [Google Scholar]
- Riddle, B.R.; Dawson, M.N.; Hadly, E.A.; Hafner, D.J.; Hickerson, M.J.; Mantooth, S.J.; Yoder, A.D. The role of molecular genetics in sculpting the future of integrative biogeography. Prog. Phys. Geogr. 2008, 32, 173–202. [Google Scholar] [CrossRef]
- Roderick, G.K. Geographic structure of insect populations: Gene flow, phylogeography, and their uses. Annu. Rev. Entomol. 1996, 41, 325–352. [Google Scholar] [CrossRef] [PubMed]
- Avise, J.C. Phylogeography: The History and Formation of Species; Harvard University Press: Cambridge, UK, 2000; pp. 3–36. [Google Scholar]
- Collevatti, R.G.; Rabelo, S.G.; Vieira, R.F. Phylogeography and disjunct distribution in Lychnophora ericoides (Asteraceae), an endangered cerrado shrub species. Ann. Bot.-Lond. 2009, 104, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Gentili, R.; Bacchetta, G.; Fenu, G.; Cogoni, D.; Abeli, T.; Rossi, G.; Salvatore, M.C.; Baroni, C.; Citterio, S. From cold to warm-stage refugia for boreo-alpine plants in southern European and Mediterranean mountains: The last chance to survive or an opportunity for speciation? Biodiversity 2015, 16, 247–261. [Google Scholar] [CrossRef]
- Craw, R.C.; Grehan, J.R.; Heads, M.J. Panbiogeography: Tracking the History of Life; Oxford University Press: Oxford, UK, 1999; pp. 4–6. [Google Scholar]
- Schmitt, T. Molecular biogeography of Europe: Pleistocene cycles and postglacial trends. Front. Zool. 2007, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- Varga, Z. Extra-Mediterranean Refugia, Post-Glacial Vegetation History and Area Dynamics in Eastern Central Europe. In Relict Species: Phylogeography and Conservation Biology; Habel, J.C., Assmann, T., Eds.; Springer Science & Business Media: New York, NY, USA, 2009; pp. 57–87. [Google Scholar]
- Kwiatkowski, P.; Krahulec, F. Disjunct distribution patterns in vascular flora of the Sudetes. Ann. Bot. Fenn. 2016, 53, 91–102. [Google Scholar] [CrossRef]
- Chen, J.; Wan, S.; Henebry, G.; Qi, J.; Gutman, G.; Sun, G.; Kappas, M. (Eds.) Dryland East Asia: Land Dynamics Amid Social and Climate Change; Walter de Gruyter: Berlin, Germany, 2014; p. 251. [Google Scholar]
- Jump, A.; Mátyás, C.; Penuelas, J. The paradox of altitude for latitude comparisons in species range retractions. Trends Ecol. Evol. 2009, 24, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Molnár, A.; Végvári, Z. Reconstruction of early Holocene Thermal Maximum temperatures using present vertical distribution of conifers in the Pannonian region (SE Central Europe). Holocene 2016, 27, 236–245. [Google Scholar] [CrossRef]
- Tóth, M.; Magyari, E.; Buczkó, K.; Braun, M.; Panagiotopoulos, K.; Heiri, O. Chironomid-inferred Holocene temperature changes in the South Carpathians (Romania). Holocene 2015, 25, 569–582. [Google Scholar] [CrossRef] [Green Version]
- Dobbertin, M.; Hilker, N.; Rebetez, M.; Zimmermann, N.E.; Wohlgemuth, T.; Rigling, A. The upward shift in altitude of pine mistletoe (Viscum album ssp. austriacum) in Switzerland—The result of climate warming? Int. J. Biometeorol. 2005, 50, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Rasztovits, E.; Moricz, N.; Berki, I.; Poetzelsberger, E.; Matyas, C. Evaluating the performance of stochastic distribution models for European beech at low-elevation xeric limits. Időjárás 2012, 116, 173–194. [Google Scholar]
- Ruiz-Labourdette, D.; Schmitz, M.F.; Pineda, F.D. Changes in tree species composition in Mediterranean mountains under climate change: Indicators for conservation planning. Ecol. Indic. 2013, 24, 310–323. [Google Scholar] [CrossRef]
- Walter, H.; Lieth, H. Klimadiagramm-Weltatlas; Gustav Fisher Verlag: Vienna, Austria, 1960. [Google Scholar]
- Woodward, F.I. Temperature and the distribution of plant species. Symp. Soc. Exp. Biol. 1988, 42, 59–75. [Google Scholar] [PubMed]
- Hocker, H.W. Certain aspects of climate as related to the distribution of Loblolly Pine. Ecology 1956, 37, 824–834. [Google Scholar] [CrossRef]
- Molnár, A.; Végvári, Z. Bioclimatic constraints of European mistletoe Viscum album at its southern distribution limit at past and present temporal scales, Pannonian Basin, Hungary. Clim. Res. 2017, 71, 237–248. [Google Scholar] [CrossRef]
- Root, T. Energy constraints on avian distributions and abundances. Ecology 1988, 69, 330–339. [Google Scholar] [CrossRef]
- Brown, J.H. Macroecology; The University of Chicago Press: Chicago, IL, USA, 1995. [Google Scholar]
- Feurdean, A.; Perşoiu, A.; Tanţău, I.; Stevens, T.; Magyari, E.K.; Onac, B.P.; Markovićh, S.; Andriči, M.; Connorj, S.; Fărcaş, S.; et al. Climate variability and associated vegetation response throughout Central and Eastern Europe (CEE) between 60 and 8 ka. Quat. Sci. Rev. 2014, 106, 206–224. [Google Scholar] [CrossRef] [Green Version]
- Magyari, E.K.; Jakab, G.; Bálint, M.; Kernd, Z.; Buczkóe, K.; Braunf, M. Rapid vegetation response to Lateglacial and early Holocene climatic fluctuation in the South Carpathian Mountains (Romania). Quat. Sci. Rev. 2012, 35, 116–130. [Google Scholar] [CrossRef]
- Gabler, R.; Petersen, J.; Trapasso, L.; Dorothy, M.S. Physical Geography, 9th ed.; Brooks/Cole, Cengage Learning: Belmont, CA, USA, 2008. [Google Scholar]
- Soó, R. A Magyar Flóra és Vegetáció Rendszertani-Növényföldrajzi Kézikönyve. (Handbook of the Flora and Vegetation of HUNGARY) I–VII; Akadémiai Kiadó: Budapest, Hungary, 1964–1985.
- Savulescu, T. (Ed.) Flora Republicae Popularis Romanicae. (Flora of Romania) Vol. I; Editio Academiae Rei Publicae Socialisticae Romania: Bucharest, Romania, 1952; p. 136.
- Savulescu, T. (Ed.) Flora Republicae Popularis Romanicae. (Flora of Romania) Vol. II; Editio Academiae Rei Publicae Socialisticae Romania: Bucharest, Romania, 1953; p. 155.
- Savulescu, T. (Ed.) Flora Republicae Popularis Romanicae. (Flora of Romania) Vol. III; Editio Academiae Rei Publicae Socialisticae Romania: Bucharest, Romania, 1955; pp. 404–405.
- Savulescu, T. (Ed.) Flora Republicae Popularis Romanicae. (Flora of Romania) Vol. IV; Editio Academiae Rei Publicae Socialisticae Romania: Bucharest, Romania, 1956; pp. 41, 622.
- Savulescu, T. (Ed.) Flora Republicae Popularis Romanicae. (Flora of Romania) Vol. VII; Editio Academiae Rei Publicae Socialisticae Romania: Bucharest, Romania, 1960; pp. 225–226.
- Dihoru, G.; Negran, G. Cartea Roşie a Plantelor Vasculare din România (Red Book of Vascular Plants of Romania); Editura Academiei: Buchureşti, Romania, 2009; pp. 526–527. [Google Scholar]
- Jalas, J.; Suominen, J. (Eds.) Atlas Florae Europaeae. 1. Pteridophyta; Commitee for Mapping the Flora of Europe and Societas Biologica Fennica Vanamo: Helsinki, Finland, 1972; p. 79. [Google Scholar]
- Jalas, J.; Suominen, J. (Eds.) Atlas Florae Europaeae. 3. Salicaceae to Balanophoraceae; Commitee for Mapping the Flora of Europe and Societas Biologica Fennica Vanamo: Helsinki, Finland, 1976; p. 63. [Google Scholar]
- Jalas, J.; Suominen, J. (Eds.) Atlas Florae Europaeae. 7. Caryophyllaceae (Silenoideae); Commitee for Mapping the Flora of Europe and Societas Biologica Fennica Vanamo: Helsinki, Finland, 1986; p. 65. [Google Scholar]
- Jalas, J.; Suominen, J. (Eds.) Atlas Florae Europaeae. 8. Nymphaeaceae to Ranunculaceae; Commitee for Mapping the Flora of Europe and Societas Biologica Fennica Vanamo: Helsinki, Finland, 1989; pp. 135–136, 148. [Google Scholar]
- Kurtto, A.; Lampinen, R.; Junikka, L. Atlas Florae Europaeae. 13. Rosaceae (Spiraea to Fragaria, Excl. Rubus); Cambridge United Kingdom: Helsinki, Finland, 2004; p. 219. [Google Scholar]
- Bogosavljević, S.; Zlatković, B.; Ranđelović, V. Flora klisure Svrljiškog Timoka Stefan Bogosavljević. In Proceedings of the 9th Symposium on Flora of Southeastern Serbia and Neighbouring Regions, Niš, Serbia, 1–3 September 2007; pp. 41–54. [Google Scholar]
- Marković, S.M.; Stankov, J.V.; Mitić, V. Study of oak forests and scrubs of hornbeam vegetation, metals content of Teucrium chamedrys and soils the first year after wildfire on Vidlic Mountain. Saf. Eng. 2015, 5, 61–68. [Google Scholar]
- Petrova, A.; Vladimirov, V. Balkan endemics in the Bulgarian flora. Phytol. Balc. 2010, 16, 293–311. [Google Scholar]
- Jávorka, S. Magyar Flóra (Flora Hungarica); Magyar Nemzeti Múzeum Növénytára: Budapest, Hungary, 1924; pp. 691, 796. [Google Scholar]
- Stupar, V.; Milanović, D.; Brujić, J.; Čarni, A. Formalized classification and nomenclatural revision of thermophilous deciduous forests (Quercetalia pubescentis) of Bosnia and Herzegovina. Tuexenia 2015, 35, 85–130. [Google Scholar]
- Prodanović, D.; Krivošej, Z.; Lazarević, P.; Amidžić, L. Contribution to the knowledge of serpentine flora in Kosovo’s part of the Ibar river valley. Bot. Serb. 2010, 34, 81–86. [Google Scholar]
- Mártonfi, P.; Kolarcik, V.; Somlyay, L. Lectotypification of Onosma viridis and synonymization of O. tornensis with O. viridis (Boraginaceae). Ann. Bot. Fenn. 2014, 51, 201–206. [Google Scholar]
- Tatár, M. A Pannoniai Flóra Endemikus Fajai. Bölcsészdoktori Értekezés. (Endemic Species of the Pannonian Flora. Dissertation); Debreceni Református Kollégium Tanárképző Intézetének kiadása: Debrecen, Hungary, 1938. [Google Scholar]
- Oeyen, J.P.; Funke, S.; Böhme, W.; Wesener, T. The evolutionary history of the rediscovered Austrian population of the giant centipede Scolopendra cingulata Latreille 1829 (Chilopoda, Scolopendromorpha). PLoS ONE 2014, 9, e108650. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
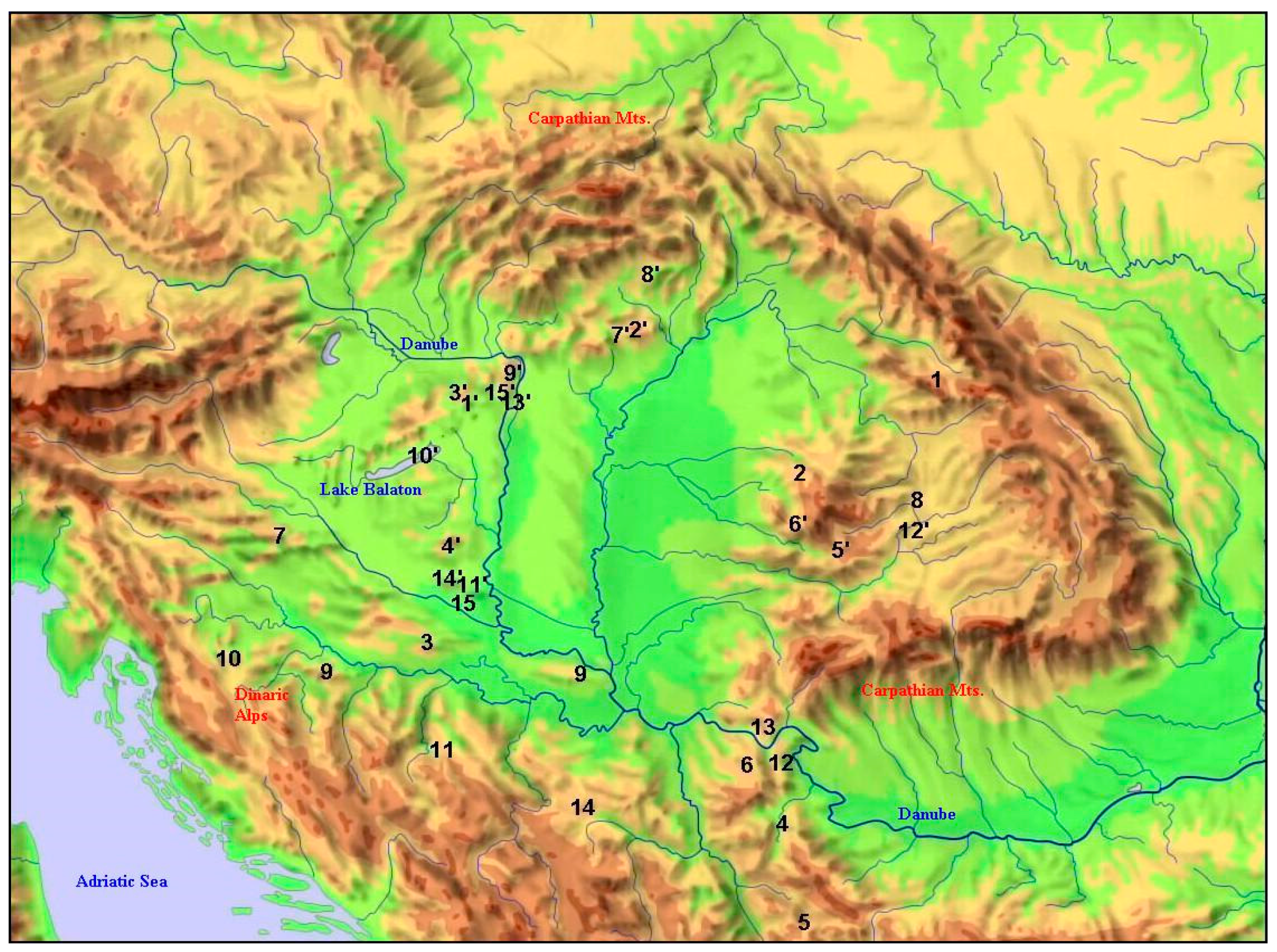
| Species | Southern, Continuous Occurrence 1 (Degree of Latitude) | Northern, Isolated Occurrence 2 (Degree of Latitude) | N-SD (Distance in Degree of Latitude) | Difference in Temperature as Derived from N-SD (°C) |
|---|---|---|---|---|
| 1. Aethionema saxatile | 47.5 | 49.0 | 1.5 | 1.0 |
| 2. Asplenium lepidum | 47.0 | 49.0 | 2.0 | 1.4 |
| 3. Carpinus orientalis | 45.6 | 47.4 | 1.8 | 1.2 |
| 4. Cuscuta approximata | 44.1 | 46.1 | 2.0 | 1.4 |
| 5. Hypericum umbellatum | 43.1 | 46.3 | 3.2 | 2.2 |
| 6. Ilex aquifolium | 44.5 | 46.3 | 1.8 | 1.3 |
| 7. Micromeria thymifolia | 46.1 | 48.1 | 2.0 | 1.4 |
| 8. Onosma viridis | 46.6 | 49.0 | 2.4 | 1.7 |
| 9. Physospermum cornubiense | 45.2 | 47.8 | 2.6 | 1.8 |
| 10. Potentilla pedata | 45.2 | 46.9 | 1.7 | 1.2 |
| 11. Ranunculus psilostachys | 44.1 | 45.8 | 1.7 | 1.2 |
| 12. Saponaria bellidifolia | 44.5 | 46.5 | 2.0 | 1.4 |
| 13. Silene flavescens | 44.8 | 47.5 | 2.7 | 1.9 |
| 14. Trigonella gladiata | 44.1 | 45.8 | 1.7 | 1.2 |
| 15. Vincetoxicum pannonicum | 45.8 | 47.5 | 1.7 | 1.2 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molnár, A.; Végvári, Z.; Tóthmérész, B. Identification of Floral Relicts Based on Spatial Distance of Isolation. Forests 2017, 8, 459. https://doi.org/10.3390/f8110459
Molnár A, Végvári Z, Tóthmérész B. Identification of Floral Relicts Based on Spatial Distance of Isolation. Forests. 2017; 8(11):459. https://doi.org/10.3390/f8110459
Chicago/Turabian StyleMolnár, Attila, Zsolt Végvári, and Béla Tóthmérész. 2017. "Identification of Floral Relicts Based on Spatial Distance of Isolation" Forests 8, no. 11: 459. https://doi.org/10.3390/f8110459
APA StyleMolnár, A., Végvári, Z., & Tóthmérész, B. (2017). Identification of Floral Relicts Based on Spatial Distance of Isolation. Forests, 8(11), 459. https://doi.org/10.3390/f8110459