Development of Multiplexed Marker Sets to Identify the Most Relevant Poplar Species for Breeding



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
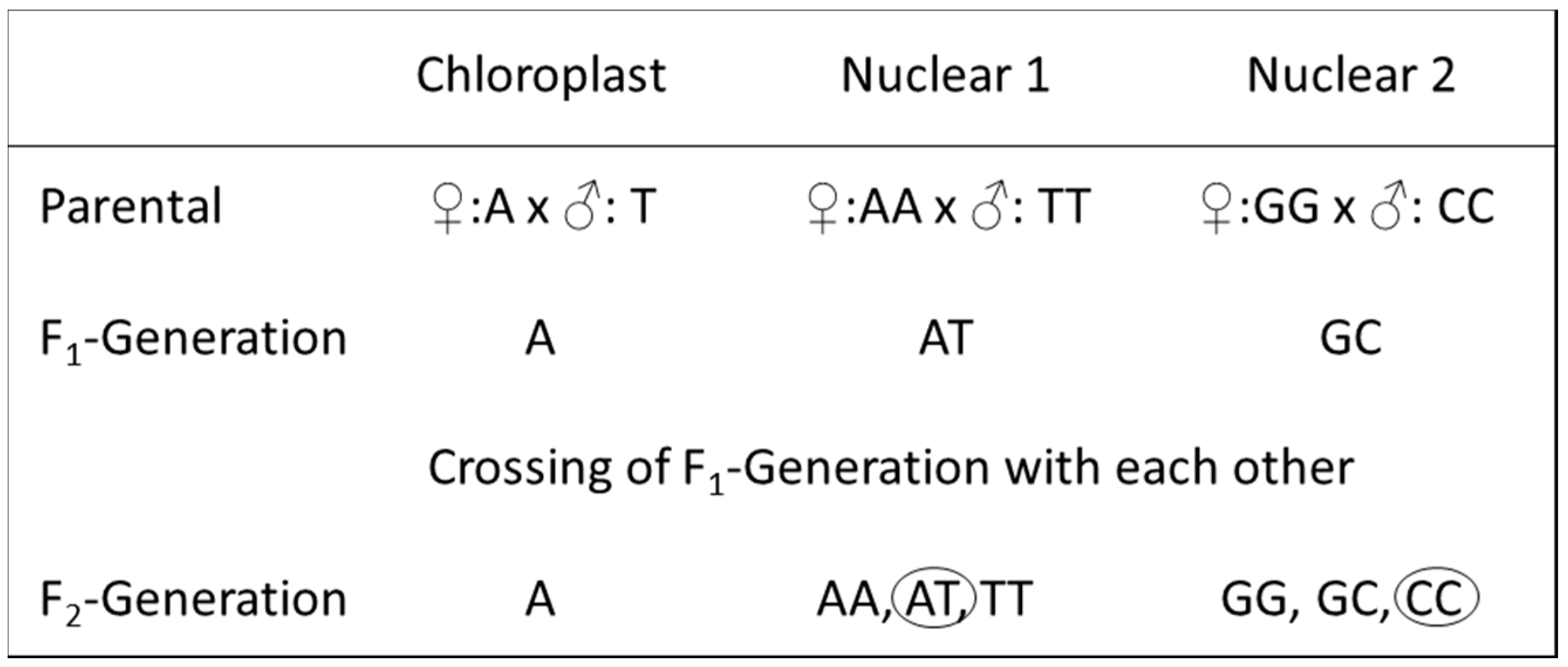
2.2. Tested Chloroplast and Nuclear Regions
2.3. DNA Extraction and PCR Amplification
2.4. Sanger Sequencing and PCR-RFLP
3. Results
3.1. Amount of Variations
3.1.1. Variations within the Chloroplast
3.1.2. Variations within the Nuclear Regions
3.1.3. Variations per Base Pair
3.2. Applicability of the Molecular Markers
3.2.1. Use of Single Markers for Species Identification
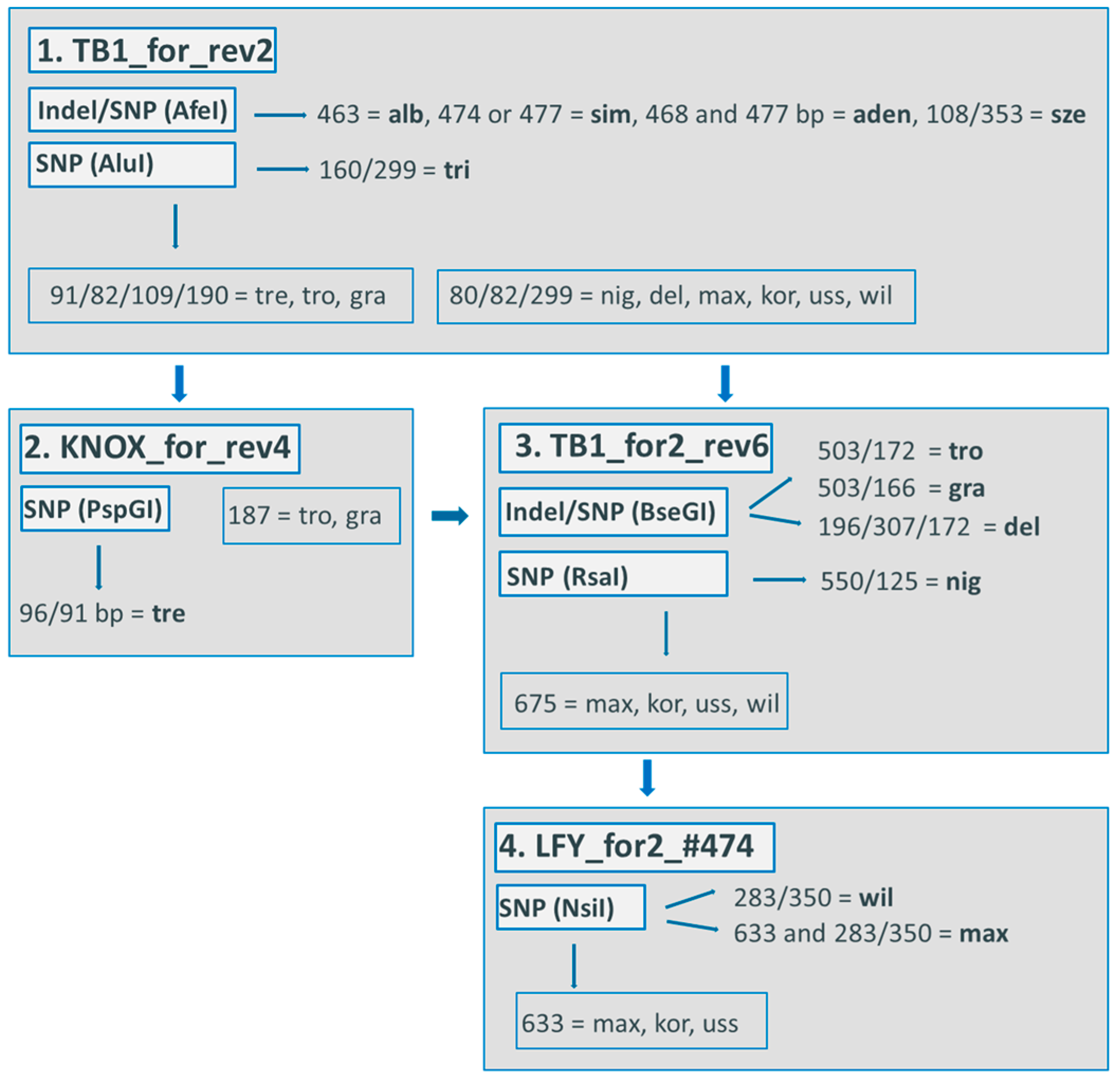
3.2.2. Cp Indel Marker Set
3.2.3. Procedure of Exclusion for Nuclear Markers
3.2.4. Reconstruction of Genealogy
4. Discussion
4.1. Differences between Species
4.2. Comparison of Cp and Nc Genomic Regions
4.3. Applicability
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dickman, D.I.; Stuart, K. The Culture of Poplars in Eastern North America; Department of Forestry, Michigan State University: East Lansing, MI, USA, 1983. [Google Scholar]
- Eckenwalder, J.E. Systematics and evolution of Populus. In Biology of Populus and its Implications for Management and Conservation Part I; Stettler, R.F., Bradshaw, H.D., Jr., Heilman, P.E., Hinckley, T.M., Eds.; NRC Research Press: Ottawa, ON, Canada, 1996; pp. 7–32. [Google Scholar]
- Stettler, R.; Bradshaw, T.; Heilman, P.; Hinckley, T. Biology of Populus and its Implications for Management and Conservation; NRC Research Press: Ottawa, ON, Canada, 1996; p. 542. [Google Scholar]
- Rahman, M.H.; Rajora, O.P. Microsatellite DNA fingerprinting, differentiation, and genetic relationships of clones, cultivars, and varieties of six poplar species from three sections of the genus Populus. Genome 2002, 45, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Rajora, O.P.; Rahman, M.H. Microsatellite DNA and RAPD fingerprinting, identification and genetic relationships of hybrid poplar (Populus × canadensis). Theor. Appl. Genet. 2003, 106, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Orlović, S.; Galović, V.; Zorić, M.; Kovačović, B.; Pilipović, A.; Galić, Z. Evaluation of interspecific DNA variability in poplars using AFLP and SSR markers. Afr. J. Biotechnol. 2009, 8, 5241–5247. [Google Scholar] [CrossRef]
- Schroeder, H.; Fladung, M. SSR and SNP markers for identification of clones, hybrids and species within the genus Populus. Silv. Genet. 2010, 59, 257–263. [Google Scholar] [CrossRef]
- Bruegmann, T.; Fladung, M. Potentials and limitations of the cross-species transfer of nuclear microsatellite marker in six species belonging to three sections of the genus Populus L. Tree Gen. Genom. 2013, 9, 1413–1421. [Google Scholar] [CrossRef]
- Ingvarsson, P.K. Nucleotide polymorphism and linkage disequilibrium within and among natural populations of European aspen (Populus tremula L., Salicaceae). Genetics 2005, 169, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, E.J.; Haughn, G.W.; Ying, C.C.; Otto, S.P.; Zhuang, J.; Cheung, D.; Hamberger, B.; Aboutorabi, F.; Kalynyak, T.; Johnson, L.; et al. Use of Ecotilling as an efficient SNP discovery tool to survey genetic variation in wild populations of Populus trichocarpa. Mol. Ecol. 2006, 15, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Meirmans, P.G.; Lamothe, M.; Périnet, P.; Isabel, N. Species-specific single nucleotide polymorphism markers for detecting hybridization and introgression in poplar. Can. J. Bot. 2007, 85, 1082–1091. [Google Scholar] [CrossRef]
- Fladung, M.; Buschbom, J. Identification of single nucleotide polymorphisms in different Populus species. Trees 2009, 23, 1199–1212. [Google Scholar] [CrossRef]
- Kersten, B.; Voss, M.-M.; Fladung, M. Development of mitochondrial SNP markers to differentiate Populus species. Trees 2015, 29, 575–582. [Google Scholar] [CrossRef]
- Tuskan, G.A.; DiFazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- Okumura, S.; Sawada, S.; Park, Y.W.; Hayashi, T.; Shimamura, M.; Takase, H. Transformation of poplar (Populus alba) plastids and expression of foreign proteins in tree chloroplasts. Transgenic Res. 2006, 15, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.I.; Hefer, C.A.; Kolosova, N.; Douglas, C.J.; Cronk, Q.C.B. Whole plastome sequencing reveals deep plastid divergence and cytonuclear discordance between closely related balsam poplars, Populus balsamifera and P. trichocarpa (Salicaceae). New Phytol. 2014, 204, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.J.; Gao, L.Z. The complete chloroplast genome sequence of desert poplar (Populus euphratica). Mitochondrial DNA Part A 2016, 27, 721–723. [Google Scholar] [CrossRef] [PubMed]
- Kersten, B.; Rampant, P.F.; Mader, M.; Le Paslier, M.-C.; Bounon, R.; Berard, A.; Vettori, C.; Schroeder, H.; Leplé, J.-C.; Fladung, M. Genome sequences of Populus tremula chloroplast and mitochondrium: Implications for holistic poplar breeding. PLoS ONE 2016, 11, e0147209. [Google Scholar] [CrossRef] [PubMed]
- Shang, C.; Du, F.K.; Yin, K.; Zhang, Z. The complete chloroplast genome of Cathay polar: Populus cathayana Rehder. Mitochondrial DNA Part B 2016, 1, 86–87. [Google Scholar] [CrossRef]
- Zhang, L.; Shang, C.; Du, F.K.; Zhao, F.; Ziong, B.; Zhang, Z. Chloroplast phylogenomic analyses maternal relationships among sections in the genus Populus. Biochem. Syst. Ecol. 2017, 70, 132–140. [Google Scholar] [CrossRef]
- Birky, C.W. Uniparental inheritance of mitochondrial and chloroplast genes—Mechanisms and evolution. Proc. Natl. Acad. Sci. USA 1995, 92, 11331–11338. [Google Scholar] [CrossRef] [PubMed]
- Barcode of Life. Available online: http://www.barcodeoflife.org/ (accessed on 16 August 2017).
- Chase, M.W.; Salamin, N.; Wilkinson, M.; Dunwell, J.M.; Kesanakurthi, R.P.; Haidar, N.; Savolainen, V. Land plants and DNA barcodes: Short-term and long-term goals. Philos. Trans. R. Soc. Lond. Ser. B 2005, 360, 1889–1895. [Google Scholar] [CrossRef] [PubMed]
- Lahaye, R.; Van der Bank, M.; Bogarin, D.; Warner, J.; Pupulin, F.; Gigot, G.; Maurin, O.; Duthoit, S.; Barraclough, T.G.; Savolainen, V. DNA barcoding the floras of biodiversity hotspots. Proc. Natl. Acad. Sci. USA 2008, 105, 2923–2928. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, M.L.; Clark, A.A.; Forrest, L.L.; Richardson, J.; Pennington, R.T.; Long, D.G.; Cowan, R.; Chase, M.W.; Gaudeul, M.; Hollingsworth, P.M. Selecting barcoding loci for plants: Evaluation of seven candidate loci with species-level sampling in three divergent groups of land plants. Mol. Ecol. Resour. 2009, 9, 439–457. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, H.; Hoeltken, A.M.; Fladung, M. Differentiation of Populus species using chloroplast SNP-markers—Essential for comprehensible and reliable poplar breeding. Plant Biol. 2012, 14, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Jiang, D.; Shang, H.; Dong, M.; Wang, G.; He, X.; Zhao, C.; Mao, K. Barcoding poplars (Populus L.) from Western China. PLoS ONE 2013, 8, e71710. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, H.; Fladung, M. Differentiation of Populus species by chloroplast SNP markers for barcoding and breeding approaches. IFOREST 2014, 8, 544–546. [Google Scholar] [CrossRef]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Chloroplast DNA phylogeny, reticulate evolution, and biogeography of Paeonia (Paeoniaceae). Am. J. Bot. 1997, 84, 1120–1136. [Google Scholar] [CrossRef] [PubMed]
- Dumolin, S.; Demesure, B.; Petit, R.J. Inheritance of chloroplast and mitochondrial genomes in pedunculated oak investigated with efficient PCR method. Theor. Appl. Genet. 1995, 91, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Fan, L.; Milne, R.I.; Zhang, L.; Wang, Y.; Mao, K. Species delimitation and lineage separation history of a species complex of aspen in China. Front. Plant Sci. 2017, 8, 375. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Liu, J.; Yu, J.; Wang, L.; Zhou, S. Highly variable chloroplast markers for evaluating plant phylogeny at low taxonomic levels and for DNA barcoding. PLoS ONE 2012, 7, e35071. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Du, S.; Dayanandan, S.; Wang, D.; Zeng, Y.; Zhang, J. Phylogeny reconstruction and hybrid analysis of Populus (Salicaceae) based on nuclear sequences of multiple single-copy nuclear genes and plastid fragments. PLoS ONE 2014, 9, e103645. [Google Scholar] [CrossRef] [PubMed]
- Simeone, M.C.; Piredda, R.; Papini, A.; Vessella, F.; Schirone, B. Application of plastid and nuclear markers to DNA barcoding of Euro-Mediterranean oaks (Quercus, Fagaceae): Problems, prospects and phylogenetic implications. Bot. J. Linn. Soc. 2013, 172, 478–499. [Google Scholar] [CrossRef]
- Mort, M.E.; Archibald, J.K.; Randle, C.P.; Levsen, N.D.; O’Leary, T.R.; Topalov, K.; Wiegand, C.M.; Crawford, D.J. Inferring phylogeny at low taxonomic levels: Utility of rapidly evolving cpDNA and nuclear ITS loci. Am. J. Bot. 2007, 94, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Petit, R.J.; Excoffier, L. Gene flow and species delimitation. Trends Ecol. Evol. 2009, 24, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Lexer, C.; Fay, M.F.; Joseph, J.A.; Nica, M.-S.; Heinze, B. Barrier to gene flow between two ecologically divergent Populus species, P. alba (white poplar) and P. tremula (European aspen): The role of ecology and life history in gene introgression. Mol. Ecol. 2005, 14, 1045–1057. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, H.; Ceulemans, R.; Davis, J.; Stettler, R. Emerging model systems in plant biology: Poplar (Populus) as a model forest tree. J. Plant Growth Regul. 2000, 19, 306–313. [Google Scholar] [CrossRef]
- Vanden Broeck, A.; Cox, K.; Quataert, P.; Van Bockstaele, E.; Van Slycken, J. Flowering phenology of Populus nigra L., P. nigra cv. italica and P. × canadensis Moench. And the potential for natural hybridisation in Belgium. Silv. Genet. 2003, 52, 280–283. [Google Scholar]
- DeWoody, J.; Trewin, H.; Taylor, G. Genetic and morphological differentiation in Populus nigra L.: Isolation by colonization or isolation by adaptation? Mol. Ecol. 2015, 24, 2641–2655. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Region | Primer Name | Sequence (5′-3′) or Reference | Length | Ta (°C) |
|---|---|---|---|---|
| Chloroplast | ||||
| trnH_psbA | trnHf | [29] | ||
| psbAr | [29] | 367 | 58 | |
| psbK_psbI | 4a_f | [26] | ||
| 4b_r | [26] | 434 | 55 | |
| rpoC2_rpoC1 | 8a_f | [26] | ||
| 8b_r | [26] | 533 | 60 | |
| trnV_atpE | 12a_f | [26] | ||
| 12b_r | [26] | 740 | 58 | |
| psbA_matK | 1a_f | [26] | ||
| 1b_r | [26] | 866 | 58 | |
| rps4_trnL | rps4f | GAAACGAGGCCCTCGGTAACGTG | ||
| trnLr | GTCCGTAGCGTCTACCAATTTCG | 821 | 60 | |
| rps2_rpoC2 | 7a_f | [26] | ||
| 7b_r | [26] | 524 | 58 | |
| trnC_petN | trnCf | CCAGTTCAAATCCGGGTGTCGC | ||
| petNr | GCAGCCCAAGCGAGACTTACTAT | 444 | 58 | |
| psaA_ycf13 | psaAf | GTGGTAGTATCAGGTCCTTTAGCT | ||
| ycf13r | GAAGCGCAGAATTGGAAGATC | 782 | 58 | |
| psbE_petL | indel3_f | TTCTTTGAATGAAGTTATACAACAC | ||
| indel3_r | GATACGAAGATAAGCAAAAATACC | 283 | 55 | |
| Nuclear | ||||
| GA20ox | #471_f | ATAGATTGCATCAAAACC | ||
| #743_r | TTAATTACCAGATCTATGGACATAAAT | 1688 | 52 | |
| GA20_for4 | TAATGAGGCTCAACTACTACC | |||
| GA20_rev5 | CTTACCATGAAGGTGTCACC | 202 | 54 | |
| KNOX | KNOX_int2for4 | TGGAGGACTACAATCAAATGAG | ||
| KNOX3_int2rev2 | TGAGATCATTACAGGCGTG | 982 | 52 | |
| KNOX3_int2f | CCAAACAATTAACCAGCAGC | |||
| KNOX3_int2rev4 | GCTACTGAAATGAATTTAATTTGA | 187 | 52 | |
| LEAFY | LFY_for2 | TACTCTTGATGCTCTCTCCCAAG | ||
| LFY_rev2 | CTTGAACTAAATTCCATATATAAG | 263 | 53 | |
| #474_r | TTGCTGTACTGGCTCCTCAGA | 633 | 53 | |
| TB1 | TB1_f | CTAACCATACCATGATTGCA | ||
| TB1_rev2 | TATGGATCTTGCTGTGCCT | 461 | 53 | |
| TB1_rev6 | GGTCATGAGAATTGAAATGGAG | 1369 | 53 | |
| TB1_for2 (combined rev6) | GTTTCAGGCATCAAGCTG | 675 | 53 |
| Species | SNPs/Indels | Species-Specific SNPs/Indels | Restriction Enzymes | [N] Number of Individuals |
|---|---|---|---|---|
| P. alba a | 22/3 | 2/2 | 2 | 5–13 |
| P. tremula a | 28/6 | 9/3 | 1 | 5 |
| P. tremuloides a | 40/9 | 17/7 | 4 | 5–21 |
| P. adenopoda a (P. × tomentosa) | 19/5 | 1/3 | 1 | 5 |
| P. grandidentata a | 26/12 | 4/7 | 3 | 32 |
| P. nigra b | 28/5 | 5/2 | 2 | 5–12 |
| P. deltoides b | 12/3 | 2/1 | 0 | 5–14 |
| P. fremontii b | 14/7 | 3/5 | 1 | 5–16 |
| P. trichocarpa c | - | 4/1 | 1 | 5 |
| P. maximowiczii c | 13/4 | 0/2 | 0 | 3–9 |
| P. cathayana c | 12/3 | 0/0 | 0 | 1 |
| P. koreana c | 12/3 | 0/0 | 0 | 3 |
| P. ussuriensis c | 12/3 | 0/0 | 0 | 1 |
| P. simonii c | 25/2 | 1/0 | 0 | 5 |
| P. szechuanica c | 24/3 | 0/0 | 0 | 1 |
| P. balsamifera c | 11/5 | 1/2 | 0 | 18 |
| P. wilsonii d | 17/2 | 3/2 | 0 | 1 |
| P. lasiocarpa d | 21/3 | 0/0 | 0 | 2 |
| P. euphratica e | 22/7 | 7/4 | 0 | 1 |
| Species | SNPs/Indels | Species-Specific SNPs/Indels | Restriction Enzymes | [N] Number of Individuals |
|---|---|---|---|---|
| P. alba a | 119/14 | 13/4 | 2 | 6 |
| P. tremula a | 107/12 | 8/0 | 1 | 5 |
| P. tremuloides a | 113/10 | 8/0 | 0 | 4–6 |
| P. adenopoda 2,a (P. × tomentosa) | 35/3 | 1/0 | 0 | 3–5 |
| P. grandidentata 2,a | 36/4 | 3/2 | 1 | 5–32 |
| P. nigra b | 53/12 | 17/1 | 4 | 4–6 |
| P. deltoide s b | 48/11 | 18/0 | 3 | 4–5 |
| P. fremontii 1,b | 23/0 | 6/0 | 0 | 10 |
| P. trichocarpa c | - | 7/0 | 2 | 5–7 |
| P. maximowiczii c | 52/11 | 4/0 | 0 | 4–6 |
| P. cathayana c | 17/0 | 0/0 | 0 | 1 |
| P. koreana c | 36/6 | 1/0 | 0 | 3 |
| P. ussuriensis c | 28/6 | 0/0 | 0 | 1 |
| P. simonii c | 49/10 | 7/3 | 2 | 3 |
| P. szechuanica c | 32/4 | 2/0 | 1 | 1 |
| P. balsamifera c | 27/7 | 1/0 | 0 | 3–18 |
| P. wilsonii d | 35/4 | 3/0 | 1 | 1 |
| P. lasiocarpa 2,d | 9/0 | 1/0 | 0 | 2 |
| P. euphratica 2,e | 14/2 | 6/1 | 2 | 1 |
| Species | Overall cp Variations | Species-Specific cp Variations | Overall nc Variations | Species-Specific nc Variations | ||||
|---|---|---|---|---|---|---|---|---|
| SNP | Indel | SNP | Indel | SNP | Indel | SNP | Indel | |
| P. alba a | 0.40 | 0.05 | 0.04 | 0.04 | 2.54 | 0.30 | 0.28 | 0.09 |
| P. tremula a | 0.51 | 0.11 | 0.16 | 0.05 | 2.29 | 0.26 | 0.17 | 0.00 |
| P. tremuloides a | 0.73 | 0.16 | 0.31 | 0.13 | 2.42 | 0.21 | 0.17 | 0.00 |
| P. adenopoda 2,a (P. × tomentosa) | 0.34 | 0.09 | 0.02 | 0.05 | 0.75 | 0.06 | 0.02 | 0.00 |
| P. grandidentata 2,a | 0.47 | 0.22 | 0.07 | 0.13 | 0.77 | 0.09 | 0.06 | 0.04 |
| P. nigra b | 0.51 | 0.09 | 0.09 | 0.04 | 1.13 | 0.26 | 0.36 | 0.02 |
| P. deltoids b | 0.22 | 0.05 | 0.04 | 0.02 | 1.03 | 0.24 | 0.38 | 0.00 |
| P. fremontii 1,b | 0.25 | 0.13 | 0.05 | 0.09 | 0.49 | 0.00 | 0.13 | 0.00 |
| P. trichocarpa c | - | - | 0.07 | 0.02 | - | - | 0.15 | 0.00 |
| P. maximowiczii c | 0.24 | 0.07 | 0.00 | 0.04 | 1.11 | 0.24 | 0.09 | 0.00 |
| P. cathayana c | 0.22 | 0.05 | 0.00 | 0.00 | 0.36 | 0.00 | 0.00 | 0.00 |
| P. koreana c | 0.22 | 0.05 | 0.00 | 0.00 | 0.77 | 0.13 | 0.02 | 0.00 |
| P. ussuriensis c | 0.22 | 0.05 | 0.00 | 0.00 | 0.60 | 0.13 | 0.00 | 0.00 |
| P. simonii c | 0.45 | 0.04 | 0.02 | 0.00 | 1.05 | 0.21 | 0.15 | 0.06 |
| P. szechuanica c | 0.44 | 0.05 | 0.00 | 0.00 | 0.68 | 0.09 | 0.04 | 0.00 |
| P. balsamifera c | 0.20 | 0.09 | 0.02 | 0.04 | 0.58 | 0.15 | 0.02 | 0.00 |
| P. wilsonii d | 0.31 | 0.04 | 0.05 | 0.04 | 0.75 | 0.09 | 0.06 | 0.00 |
| P. lasiocarpa 2,d | 0.38 | 0.05 | 0.00 | 0.00 | 0.19 | 0.00 | 0.02 | 0.00 |
| P. euphratica 2,e | 0.40 | 0.13 | 0.13 | 0.07 | 0.30 | 0.04 | 0.13 | 0.02 |
| Average (species) | 0.36 | 0.09 | 0.06 | 0.04 | 0.99 | 0.14 | 0.12 | 0.01 |
| Sum over regions and species | 6.50 | 1.54 | 1.07 | 0.74 | 17.81 | 2.48 | 2.27 | 0.24 |
| Species | Region | Marker Type | Restriction Enzyme | Length [bp] | Source |
|---|---|---|---|---|---|
| P. alba | trnH_psbA | cp SNP | Alw61I | 139/228 | [26] |
| P. tremula | rpoC2_rpoC1 psbE_petL | cp SNP indel | MlsI / | 409/124 283 (−99) | [26] new |
| P. tremuloides | trnH_psbA | cp SNP | DraI | 129/238 | [26] |
| P. adenopoda | psbK_psbI | cp SNP | AseI | 144/290 | new |
| P. grandidentata | psbK_psbI | cp SNP + indel | DraI | 264/174 | new |
| P. nigra | trnV_atpE | cp SNP + indel | ApoI | 404/62/260 | new |
| P. deltoides | TB1 | nc SNP | BseGI | 194/481 | new |
| P. fremontii | trnH_psbA | cp SNP | BsmI | 102/265 | new |
| P. trichocarpa | psbK_psbI | cp SNP | TaqI | 78/209/49/43/214 | new |
| P. maximowiczii | psaA_ycf13 | cp indel | / | (ca. 790) + 22 | new |
| P. simonii | GA20ox | nc SNP | CviKI | 8/27/167 | new |
| P. szechuanica | TB1 | nc SNP | AfeI | 108/353 | new |
| P. wilsonii | trnH_psbA | cp indel | / | (367) + 4 | new |
| P. euphratica | KNOX_int2 | nc SNP | BfaI | 64/223 | new |
| PCR 1 | Concentration | PCR 2 | Concentration |
|---|---|---|---|
| BD PCR-Puffer 10× | 1× | BD PCR-Puffer 10× | 1× |
| MgCl 25 mM | 1.5 mM | MgCl 25 mM | 1.75 mM |
| dNTPs 25 mM | 0.2 mM | dNTPs 25 mM | 0.2 mM |
| rps4_2_VIC 10 μM | 0.18 μM | psbA_2_PET 10 μM | 0.2 μM |
| trnL_2_R 10 μM | 0.18 μM | matK_2_R 10 μM | 0.2 μM |
| rps2-mod_FAM 10 μM | 0.12 μM | Enhancer DMSO | |
| rpoc2_R 10 μM | 0.12 μM | Taq 5 U/μL | 0.6 U |
| trnC_FAM 10 μM | 0.04 μM | DNA 10 ng/μL | 30 ng |
| petN_R 10 μM | 0.04 μM | ||
| Enhancer DMSO | |||
| Taq 5 U/μL | 0.6 U | ||
| DNA 10 ng/μL | 30 ng |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schroeder, H.; Kersten, B.; Fladung, M. Development of Multiplexed Marker Sets to Identify the Most Relevant Poplar Species for Breeding. Forests 2017, 8, 492. https://doi.org/10.3390/f8120492
Schroeder H, Kersten B, Fladung M. Development of Multiplexed Marker Sets to Identify the Most Relevant Poplar Species for Breeding. Forests. 2017; 8(12):492. https://doi.org/10.3390/f8120492
Chicago/Turabian StyleSchroeder, Hilke, Birgit Kersten, and Matthias Fladung. 2017. "Development of Multiplexed Marker Sets to Identify the Most Relevant Poplar Species for Breeding" Forests 8, no. 12: 492. https://doi.org/10.3390/f8120492
APA StyleSchroeder, H., Kersten, B., & Fladung, M. (2017). Development of Multiplexed Marker Sets to Identify the Most Relevant Poplar Species for Breeding. Forests, 8(12), 492. https://doi.org/10.3390/f8120492