
Estimating Carbon Dynamics in an Intact Lowland Mixed Dipterocarp Forest Using a Forest Carbon Model

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Estimation of Biomass C Stock and Growth Rate
2.3. Simulation of Forest C Dynamics with the FBDC Model
3. Results
3.1. Initial C Stocks
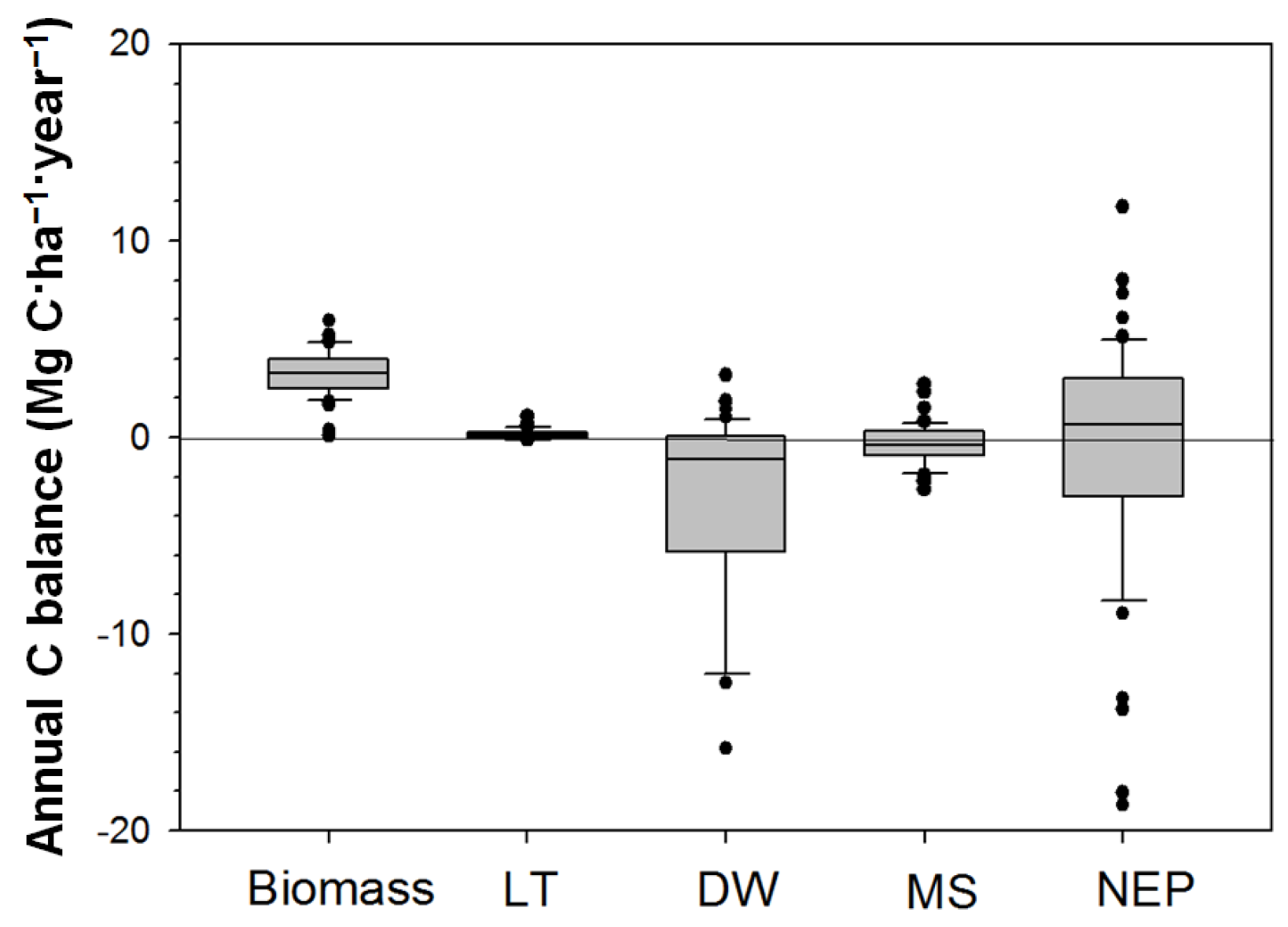
3.2. Changes in C Stocks and NEP
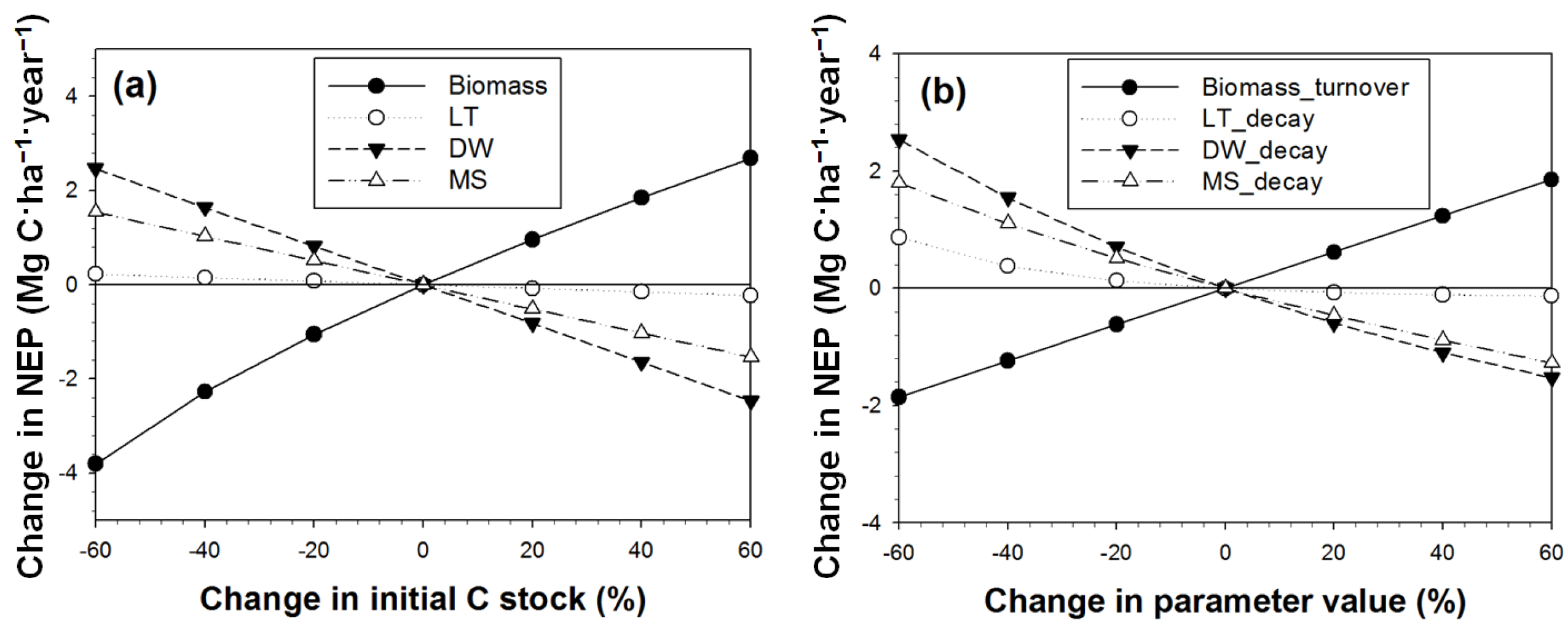
3.3. Sensitivity Analysis
4. Discussion
4.1. C Dynamics in the Intact Tropical Forests
4.2. Implications
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gautam, S.; Pietsch, S.A. Carbon pools of an intact forest in Gabon. Afr. J. Ecol. 2012, 50, 414–427. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.J. The carbon sink in intact tropical forests. Glob. Chang. Biol. 2013, 19, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Miah, M.D.; Akther, S.; Shin, M.; Koike, M. Scaling up REDD+ strategies in Bangladesh: A forest dependence study in the Chittagong Hill Tracts. For. Sci. Technol. 2014, 10, 148–156. [Google Scholar] [CrossRef]
- Lee, S.; Lee, D.; Yoon, T.K.; Salim, K.A.; Han, S.; Yun, H.M.; Yoon, M.; Kim, E.; Lee, W.-K.; Davies, S.J.; et al. Carbon stocks and its variations with topography in an intact lowland mixed dipterocarp forest in Brunei. J. Ecol. Environ. 2015, 38, 75–84. [Google Scholar] [CrossRef]
- Yamakura, T.; Hagihara, A.; Sukardjo, S.; Ogawa, H. Aboveground biomass of tropical rain forest stands in Indonesian Borneo. Vegetatio 1986, 68, 71–82. [Google Scholar]
- Basuki, T.M.; Van Laake, P.E.; Skidmore, A.K.; Hussin, Y.A. Allometric equations for estimating the above-ground biomass in tropical lowland Dipterocarp forests. For. Ecol. Manag. 2009, 257, 1684–1694. [Google Scholar] [CrossRef]
- Kralicek, K.; Poudel, K.P.; Temesgen, T.; Salas, C. Simultaneous estimation of above- and below-ground biomass in tropical forests of Viet Nam. For. Ecol. Manag. 2017, 390, 147–156. [Google Scholar] [CrossRef]
- Chapagain, T.R.; Sharma, R.P.; Bhandari, S.K. Modeling above-ground biomass for three tropical tree species at their juvenile stage. For. Sci. Technol. 2014, 10, 51–60. [Google Scholar] [CrossRef]
- Kangkuso, A.; Jamili, J.; Septiana, A.; Raya, R.; Sahidin, I.; Rianse, U.; Rahim, S.; Alfirman, A.; Sharma, S.; Nadaoka, K. Allometric models and aboveground biomass of Lumnitzera racemosa Willd. forest in Rawa Aopa Watumohai National Park, Southeast Sulawesi, Indonesia. For. Sci. Technol. 2016, 12, 43–50. [Google Scholar]
- Kosugi, Y.; Takanashi, S.; Ohkubo, S.; Matsuo, N.; Tani, M.; Mitani, T.; Tsutsumi, D.; Nik, A.R. CO2 exchange of a tropical rainforest at Pasoh in Peninsular Malaysia. Agric. For. Meteorol. 2008, 148, 439–452. [Google Scholar] [CrossRef]
- Malhi, Y.; Aragão, L.E.O.C.; Metcalfe, D.B.; Paiva, R.; Quesada, C.A.; Almeida, S.; Anderson, L.; Brando, P.; Chambers, J.Q.; Da Costa, A.C.L.; et al. Comprehensive assessment of carbon productivity, allocation and storage in three Amazonian forests. Glob. Chang. Biol. 2009, 15, 1255–1274. [Google Scholar] [CrossRef]
- Saitoh, T.M.; Kumagai, T.; Sato, Y.; Suzuki, M. Carbon dioxide exchange over a Bornean tropical rainforest. J. Agric. Meteorol. 2005, 60, 553–556. [Google Scholar] [CrossRef]
- Baldocchi, D.D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: Past, present and future. Glob. Chang. Biol. 2003, 9, 479–492. [Google Scholar] [CrossRef]
- Kim, H.; Kim, Y.-H.; Kim, R.; Park, H. Reviews of forest carbon dynamics models that use empirical yield curves: CBM-CFS3, CO2FIX, CASMOFOR, EFISCEN. For. Sci. Technol. 2015, 11, 212–222. [Google Scholar] [CrossRef]
- Sierra, C.A.; Harmon, M.E.; Moreno, F.H.; Orrego, S.A.; Del Valle, J.I. Spatial and temporal variability of net ecosystem production in a tropical forest: Testing the hypothesis of a significant carbon sink. Glob. Chang. Biol. 2007, 13, 838–853. [Google Scholar] [CrossRef]
- Huang, M.; Asner, G.P.; Keller, M.; Berry, J.A. An ecosystem model for tropical forest disturbance and selective logging. J. Geophys. Res. 2008, 113, G01002. [Google Scholar] [CrossRef]
- Hashimoto, H.; Melton, F.; Ichii, K.; Milesi, C.; Wang, W.; Nemani, R.R. Evaluating the impacts of climate and elevated carbon dioxide on tropical rainforests of the western Amazon basin using ecosystem models and satellite data. Glob. Chang. Biol. 2010, 16, 255–271. [Google Scholar] [CrossRef]
- Kim, Y.; Knox, R.G.; Longo, M.; Medvigy, D.; Hutyra, L.R.; Pyle, E.H.; Wofsy, S.C.; Bras, R.; Moorcroft, P.R. Seasonal carbon dynamics and water fluxes in an Amazon rainforest. Glob. Chang. Biol. 2012, 18, 1322–1334. [Google Scholar] [CrossRef]
- Lee, J.; Yoon, T.K.; Han, S.; Kim, S.; Yi, M.J.; Park, G.S.; Kim, C.; Son, Y.M.; Kim, R.; Son, Y. Estimating the carbon dynamics of South Korean forests from 1954 to 2012. Biogeosciences 2014, 11, 4637–4650. [Google Scholar] [CrossRef]
- Lee, J.; Tolunay, D.; Makineci, E.; Çömez, A.; Son, Y.M.; Kim, R.; Son, Y. Estimating the age-dependent changes in carbon stocks of Scots pine (Pinus sylvestris L.) stands in Turkey. Ann. For. Sci. 2016, 73, 523–531. [Google Scholar] [CrossRef]
- Small, A.; Martin, T.G.; Kitching, R.L.; Wong, K.M. Contribution of tree species to the biodiversity of a 1 ha old world rainforest in Brunei, Borneo. Biodivers. Conserv. 2004, 13, 2067–2088. [Google Scholar] [CrossRef]
- Ashton, P.S.; Hall, P. Comparisons of structure among mixed dipterocarp forests of north-western Borneo. J. Ecol. 1992, 90, 459–481. [Google Scholar] [CrossRef]
- Kenzo, T.; Ichie, T.; Hattori, D.; Itioka, T.; Handa, C.; Ohkubo, T.; Kendawang, J.J.; Nakamura, M.; Sakaguchi, M.; Takahashi, N.; et al. Development of allometric relationships for accurate estimation of above- and below-ground biomass in tropical secondary forests in Sarawak, Malaysia. J. Trop. Ecol. 2009, 25, 371–386. [Google Scholar] [CrossRef]
- Huy, B.; Poudel, K.P.; Kralicek, K.; Hung, N.D.; Khoa, P.V.; Phương, V.T.; Temesgen, H. Allometric equations for estimating tree aboveground biomass in tropical dipterocarp forests of Vietnam. Forests 2016, 7, 180. [Google Scholar] [CrossRef]
- Kenzo, T.; Furutani, R.; Hattori, D.; Tanaka, S.; Sakurai, K.; Ninomiya, I.; Kendawang, J.J. Aboveground and belowground biomass in logged-over tropical rain forests under different soil conditions in Borneo. J. For. Res. 2015, 20, 197–205. [Google Scholar] [CrossRef]
- Niiyama, K.; Kajimoto, T.; Matsuura, Y.; Yamashita, T.; Matsuo, N.; Yashiro, Y.; Ripin, A.; Kassim, R.; Noor, N.S. Estimation of root biomass based on excavation of individual root systems in a primary dipterocarp forest in Pasoh Forest Reserve, Peninsular Malaysia. J. Trop. Ecol. 2010, 26, 271–284. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Good Practice Guidance for Land Use, Land-Use Change and Forestry; IPCC: Kanagawa, Japan, 2003. [Google Scholar]
- The Statistical Analysis System Institute (SAS Institute). SAS 9.4 Output Delivery System: User’s Guide; SAS Institute: Cary, NC, USA, 2014. [Google Scholar]
- Pransiska, W.; Triadiati, T.; Tjitrosoedirjo, S.; Hertel, D.; Kotowska, M.M. Forest conversion impacts on the fine and coarse root system, and soil organic matter in tropical lowlands of Sumatera (Indonesia). For. Ecol. Manag. 2016, 379, 288–298. [Google Scholar] [CrossRef]
- Brearley, F.Q.; Prajadinata, S.; Kidd, P.S.; Proctor, J.; Suriantata. Structure and floristics of an old secondary rain forest in Central Kalimantan, Indonesia, and a comparison with adjacent primary forest. For. Ecol. Manag. 2004, 195, 385–397. [Google Scholar] [CrossRef]
- Gill, R.A.; Jackson, R.B. Global patterns of root turnover for terrestrial ecosystems. New Phytol. 2000, 147, 13–31. [Google Scholar] [CrossRef]
- Dent, D.H.; Bagchi, R.; Robinson, D.; Majalap-Lee, N.; Burslem, D.F.P.B. Nutrient fluxes via litterfall and leaf litter decomposition vary across a gradient of soil nutrient supply in a lowland tropical rain forest. Plant Soil 2006, 288, 197–215. [Google Scholar] [CrossRef]
- Silver, W.; Thompson, A.W.; McGroddy, M.E.; Varner, R.K.; Dias, J.D.; Silva, H.; Crill, P.M.; Keller, C. Fine root dynamics and trace gas fluxes in two lowland tropical soils. Glob. Chang. Biol. 2005, 11, 290–306. [Google Scholar] [CrossRef]
- Liski, J.; Palosou, T.; Peltoniemi, M.; Sievänen, R. Carbon and decomposition model Yasso for forest soils. Ecol. Model. 2005, 189, 168–182. [Google Scholar] [CrossRef]
- Zhou, T.; Shi, P.; Hui, D.; Luo, Y. Global pattern of temperature sensitivity of soil heterotrophic respiration (Q10) and its implications for carbon-climate feedback. J. Geophys. Res. 2009, 114, G02016. [Google Scholar] [CrossRef]
- Rüger, N.; Berger, U.; Hubbell, S.P.; Vieilledent, G.; Condit, R. Growth strategies of tropical species: Disentangling light and size effects. PLoS ONE 2011, 6, e25330. [Google Scholar]
- Stephenson, N.L.; Das, A.J.; Condit, R.; Russo, S.E.; Baker, P.J.; Beckman, N.G.; Coomes, D.A.; Lines, E.R.; Morris, W.K.; Rüger, N.; et al. Rate of tree carbon accumulation increases continuously with tree size. Nature 2014, 507, 90–93. [Google Scholar] [CrossRef] [PubMed]
- King, D.A.; Davies, S.J.; Md. Noor, N.S. Growth and mortality are related to adult tree size in a Malaysian mixed dipterocarp forest. For. Ecol. Manag. 2006, 223, 152–158. [Google Scholar] [CrossRef]
- Pyle, E.H.; Santoni, G.W.; Nascimento, H.E.M.; Hutyra, L.R.; Vieira, S.; Curran, D.J.; van Haren, J.; Saleska, S.R.; Chow, V.Y.; Carmago, P.B.; et al. Dynamics of carbon, biomass, and structure in two Amazonian forests. J. Geophys. Res. 2008, 113, G00B08. [Google Scholar] [CrossRef]
- Saner, P.; Loh, Y.Y.; Ong, R.C.; Hector, A. Carbon stocks and fluxes in tropical lowland dipterocarp rainforests in Sabah, Malaysian Borneo. PLoS ONE 2012, 7, e29642. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Mean Annual Temperature (°C) a | Annual Precipitation (mm) b | Number of Stems (N·ha−1) | |||
|---|---|---|---|---|---|
| DBH < 20 cm | 20 cm ≤ DBH < 50 cm | 50 cm ≤ DBH | |||
| Maximum | 24.9 | 5677.4 | 7606 | 236 | 58 |
| Average | 24.6 | 4582.0 | 6504 | 204 | 42 |
| Minimum | 24.0 | 3236.9 | 5467 | 167 | 28 |
| Category | Pools | Value | Reference |
|---|---|---|---|
| Turnover rate (year−1) | Stem | 0.0075 | [30] |
| Branches | 0.04 | Lee et al. (unpublished data) | |
| Foliage | 1.458 | Lee et al. (unpublished data) | |
| Coarse roots | 0.04 | Assumption: equal to the Branch | |
| Fine roots | 0.6635 | [31] | |
| Decay constant (k; year−1) | AWDS | 0.209 | Lee et al. (unpublished data) |
| AWDB | 1.07 | [32] | |
| ALT | 2.27 | [32] | |
| BWD | 1.07 | Assumption: equal to the AWDB | |
| BLT | 0.96 | [33] | |
| AHUM and BHUM | 0.02 | [34,35] | |
| SOC | 0.0017 | [34,35] |
| Variables | Biomass | Litter | Dead Wood | Mineral Soil | Total |
|---|---|---|---|---|---|
| C stock (Mg C·ha−1) | 213.1 ± 104.8 | 2.0 ± 0.8 | 31.3 ± 38.8 | 80.7 ± 15.5 | 327.0 ± 102.2 |
| Relative contribution (%) | 65.2 | 0.5 | 9.6 | 24.7 | 100 |
| NEP | Biomass C | LT C | DW C | MS C | |
|---|---|---|---|---|---|
| ΔBio | 0.57 * | 0.94 * | 0.05 | −0.20 | −0.13 |
| ΔLT | 0.55 * | 0.90 * | −0.41 * | −0.23 | 0.08 |
| ΔDW | 0.95 * | 0.32 * | −0.11 | −1.00 * | 0.05 |
| ΔMS | −0.42 * | 0.37 * | 0.19 | 0.73 * | −0.50 * |
| NEP | - | 0.59 * | −0.09 | −0.92 * | −0.06 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.; Lee, S.; Han, S.H.; Kim, S.; Roh, Y.; Salim, K.A.; Pietsch, S.A.; Son, Y. Estimating Carbon Dynamics in an Intact Lowland Mixed Dipterocarp Forest Using a Forest Carbon Model. Forests 2017, 8, 114. https://doi.org/10.3390/f8040114
Lee J, Lee S, Han SH, Kim S, Roh Y, Salim KA, Pietsch SA, Son Y. Estimating Carbon Dynamics in an Intact Lowland Mixed Dipterocarp Forest Using a Forest Carbon Model. Forests. 2017; 8(4):114. https://doi.org/10.3390/f8040114
Chicago/Turabian StyleLee, Jongyeol, Sohye Lee, Seung Hyun Han, Seongjun Kim, Yujin Roh, Kamariah Abu Salim, Stephan A. Pietsch, and Yowhan Son. 2017. "Estimating Carbon Dynamics in an Intact Lowland Mixed Dipterocarp Forest Using a Forest Carbon Model" Forests 8, no. 4: 114. https://doi.org/10.3390/f8040114
APA StyleLee, J., Lee, S., Han, S. H., Kim, S., Roh, Y., Salim, K. A., Pietsch, S. A., & Son, Y. (2017). Estimating Carbon Dynamics in an Intact Lowland Mixed Dipterocarp Forest Using a Forest Carbon Model. Forests, 8(4), 114. https://doi.org/10.3390/f8040114