
Thinning of Beech Forests Stocking on Shallow Calcareous Soil Maintains Soil C and N Stocks in the Long Run


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Characteristics and Experimental Design
2.2. Experimental Design and Soil Sampling
2.3. Soil Analyses
2.4. Statistics
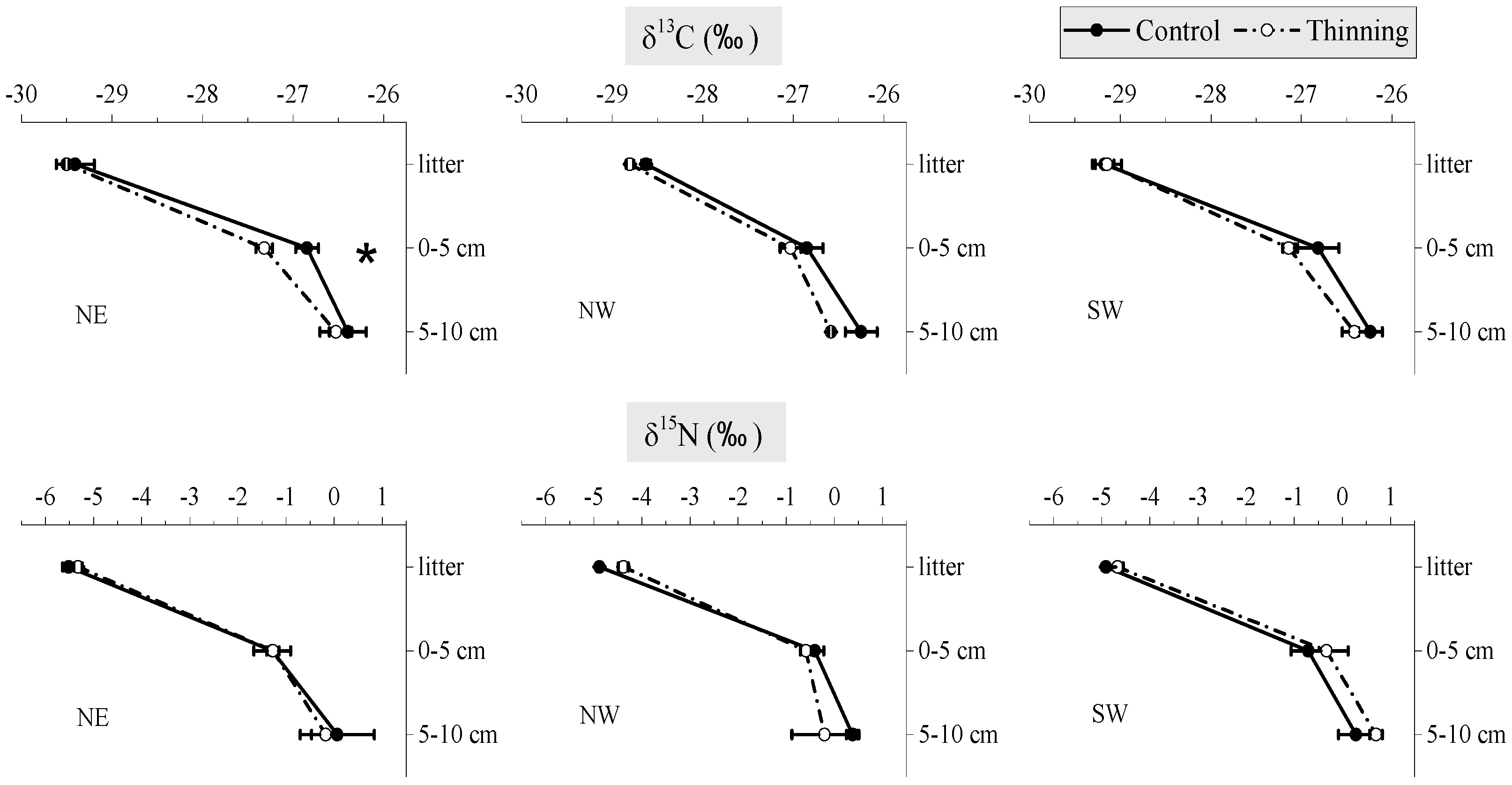
3. Results
Effects of Forest Management
4. Discussion
4.1. Forest Management Affects Soil C Turnover and Soil C Stocks in the Short, but Not in the Long Run
4.2. Forest Management Affects Soil N Turnover and Soil N Stocks in the Short, but Not in the Long Run
4.3. Synthesis of Short-Term and Long-Term Thinning Effects: Microclimate, Understorey Vegetation, and Carbon–Nitrogen Interaction
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zanchi, G.; Pena, N.; Bird, N. Is woody bioenergy carbon neutral? A comparative assessment of emissions from consumption of woody bioenergy and fossil fuel. GCB Bioenerg. 2012, 4, 761–772. [Google Scholar] [CrossRef]
- Tsunetsugu, Y.; Tonosaki, M. Quantitative estimation of carbon removal effects due to wood utilization up to 2050 in Japan: Effects from carbon storage and substitution of fossil fuels by harvested wood products. J. Wood Sci. 2006, 56, 339–344. [Google Scholar] [CrossRef]
- Lineback, N.; Dellinger, T.; Shienvold, L.; Witcher, B.; Reynolds, A.; Brown, L. Industrial greenhouse gas emissions: Does CO2 from combustion of biomass residue for energy really matter? Clim. Res. 1999, 13, 221–229. [Google Scholar] [CrossRef]
- Dixon, R.K.; Solomon, A.M.; Brown, S.; Houghton, R.A.; Trexier, M.C.; Wisniewski, J. Carbon pools and flux of global forest ecosystems. Science 1994, 263, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, P.; Schmidt, I.K.; Raulund-Rasmussen, K. Leaching of nitrate from temperate forests—Effects of air pollution and forest management. Environ. Rev. 2006, 14, 1–57. [Google Scholar] [CrossRef]
- Gregorich, E.; Greer, K.; Anderson, D.; Liang, B. Carbon distribution and losses: Erosion and deposition effects. Soil Tillage Res. 1998, 47, 291–302. [Google Scholar] [CrossRef]
- Lal, R. Restoring soil quality to mitigate soil degradation. Sustainability 2015, 7, 5875–5895. [Google Scholar] [CrossRef]
- Rennenberg, H.; Dannenmann, M.; Gessler, A.; Kreuzwieser, J.; Simon, J.; Papen, H. Nitrogen balance in forest soils: Nutritional limitation of plants under climate change stresses. Plant Biol. 2009, 11, 4–23. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.; Dannenmann, M.; Pena, R.; Gessler, A.; Rennenberg, H. Nitrogen nutrition of beech forests in a changing climate: Importance of plant-soil-microbe water, carbon, and nitrogen interactions. Plant Soil Marschner Rev. 2017. submitted. [Google Scholar]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K.A. How strongly can forest management influence soil carbon sequestration? Geoderma 2007, 137, 253–268. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Saiz, G.; Wandera, F.M.; Pelster, D.E.; Ngetich, W.; Okalebo, J.R.; Rufino, M.C.; Butterbach-Bahl, K. Long-term assessment of soil and water conservation measures (Fanya-juu terraces) on soil organic matter in South Eastern Kenya. Geoderma 2016, 274, 1–9. [Google Scholar] [CrossRef]
- Johnson, M.G.; Levine, E.R.; Kern, J.S. Soil organic matter: Distribution, genesis, and management to reduce greenhouse gas emissions. Water Air Soil Pollut. 1995, 82, 593–615. [Google Scholar] [CrossRef]
- Dannenmann, M.; Butterbach-Bahl, K.; Gasche, R.; Willibald, G.; Papen, H. Dinitrogen emissions and the N2:N2O emission ratio of a Rendzic Leptosol as influenced by pH and forest thinning. Soil Biol. Biochem. 2008, 40, 2317–2323. [Google Scholar] [CrossRef]
- Dannenmann, M.; Gasche, R.; Ledebuhr, A.; Holst, T.; Mayer, H.; Papen, H. The effect of forest management on trace gas exchange at the pedosphere-atmosphere interface in beech (Fagus sylvatica L.) forests stocking on calcareous soils. Eur. J. For. Res. 2007, 126, 331–346. [Google Scholar] [CrossRef]
- Medinets, S.; Skiba, U.; Rennenberg, H.; Butterbach-Bahl, K. A review of soil NO transformation: Associated processes and possible physiological significance on organisms. Soil Biol. Biochem. 2015, 80, 92–117. [Google Scholar] [CrossRef]
- Powers, M.D.; Kolka, R.K.; Bradford, J.B.; Palik, B.J.; Fraver, S.; Jurgensen, M.F. Carbon stocks across a chronosequence of thinned and unmanaged red pine (Pinus resinosa) stands. Ecol. Appl. 2012, 22, 1297–1307. [Google Scholar] [CrossRef] [PubMed]
- Nave, L.E.; Vance, E.D.; Swanston, C.W.; Curtis, P.S. Harvest impacts on soil carbon storage in temperate forests. For. Ecol. Manag. 2010, 259, 857–866. [Google Scholar] [CrossRef]
- Herold, N.; Schöning, I.; Michalzik, B.; Trumbore, S.; Schrumpf, M. Controls on soil carbon storage and turnover in German landscapes. Biogeochemistry 2014, 119, 435–451. [Google Scholar] [CrossRef]
- Hedde, M.; Aubert, M.; Decaëns, T.; Bureau, F. Dynamics of soil carbon in a beechwood chronosequence forest. For. Ecol. Manag. 2008, 255, 193–202. [Google Scholar] [CrossRef]
- Johnson, D.W.; Curtis, P.S. Effects of forest management on soil C and N storage: Meta analysis. For. Ecol. Manag. 2001, 140, 227–238. [Google Scholar] [CrossRef]
- Wiesmeier, M.; Schad, P.; von Lützow, M.; Poeplau, C.; Spörlein, P.; Geuß, U.; Hangen, E.; Reischl, A.; Schilling, B.; Kögel-Knabner, I. Quantification of functional soil organic carbon pools for major soil units and land uses in southeast Germany (Bavaria). Agric. Ecosyst. Environ. 2014, 185, 208–220. [Google Scholar] [CrossRef]
- Johnson, D.; Knoepp, J.; Swank, W.; Shan, J.; Morris, L.; Van Lear, D.; Kapeluck, P. Effects of forest management on soil carbon: Results of some long-term resampling studies. Environ. Pollut. 2002, 116, S201–S208. [Google Scholar] [CrossRef]
- Wall, A. Effect of removal of logging residue on nutrient leaching and nutrient pools in the soil after clearcutting in a Norway spruce stand. For. Ecol. Manag. 2008, 256, 1372–1383. [Google Scholar] [CrossRef]
- Dannenmann, M.; Gasche, R.; Ledebuhr, A.; Papen, H. Effects of forest management on soil N cycling in beech forests stocking on calcareous soils. Plant Soil 2006, 287, 279–300. [Google Scholar] [CrossRef]
- Boncina, A.; Kadunc, A.; Robic, D. Effects of selective thinning on growth and development of beech (Fagus sylvatica L.) forest stands in south-eastern Slovenia. Ann. For. Sci. 2007, 64, 44–57. [Google Scholar] [CrossRef]
- Van der Maaten, E. Thinning prolongs growth duration of European beech (Fagus sylvatica L.) across a valley in southwestern Germany. For. Ecol. Manag. 2013, 306, 135–141. [Google Scholar] [CrossRef]
- Fotelli, M.N.; Rienks, M.; Rennenberg, H.; Geßler, A. Climate and forest management affect 15N-uptake, N balance and biomass of European beech seedlings. Trees Struct. Funct. 2004, 18, 157–166. [Google Scholar] [CrossRef]
- Le Goff, N.; Ottorini, J.-M. Thinning and climate effects on growth of beech (Fagus sylvatica L.) in experimental stands. For. Ecol. Manag. 1993, 62, 1–14. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.-J.; Nabuurs, G.-J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2012, 3, 203–207. [Google Scholar] [CrossRef]
- Dannenmann, M.; Bimüller, C.; Gschwendtner, S.; Leberecht, M.; Tejedor, J.; Bilela, S.; Gasche, R.; Hanewinkel, M.; Baltensweiler, A.; Kögel-Knabner, I.; et al. Climate change impairs nitrogen cycling in European beech forests. PLoS ONE 2016, 11, e0158823. [Google Scholar] [CrossRef] [PubMed]
- Cermak, J.; Matyssek, R.; Kucera, J. Rapid response of large, drought-stressed beech trees to irrigation. Tree Physiol. 1993, 12, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Misson, L.; Vincke, C.; Devillez, F. Frequency responses of radial growth series after different thinning intensities in Norway spruce (Picea abies (L.) Karst.) stands. For. Ecol. Manag. 2003, 177, 51–63. [Google Scholar] [CrossRef]
- Ma, S.; Concilio, A.; Oakley, B.; North, M.; Chen, J. Spatial variability in microclimate in a mixed-conifer forest before and after thinning and burning treatments. For. Ecol. Manag. 2010, 259, 904–915. [Google Scholar] [CrossRef]
- Dannenmann, M.; Gasche, R.; Papen, H. Nitrogen turnover and N2O production in the forest floor of beech stands as influenced by forest management. J. Plant Nutr. Soil Sci. 2007, 170, 134–144. [Google Scholar] [CrossRef]
- Poage, M.A.; Feng, X. A theoretical analysis of steady state δ13C profiles of soil organic matter. Glob. Biogeochem. Cycles 2004, 18. [Google Scholar] [CrossRef]
- Bedard-Haughn, A.; van Groenigen, J.W.; van Kessel, C. Tracing 15N through landscapes: Potential uses and precautions. J. Hydrol. 2003, 272, 175–190. [Google Scholar] [CrossRef]
- Geßler, A.; Jung, K.; Gasche, R.; Papen, H.; Heidenfelder, A.; Börner, E.; Metzler, B.; Augustin, S.; Hildebrand, E.; Rennenberg, H. Climate and forest management influence nitrogen balance of European beech forests: Microbial N transformations and inorganic N net uptake capacity of mycorrhizal roots. Eur. J. For. Res. 2005, 124, 95–111. [Google Scholar] [CrossRef]
- Bimüller, C.; Dannenmann, M.; Tejedor, J.; von Lützow, M.; Buegger, F.; Meier, R.; Haug, S.; Schroll, R.; Kögel-Knabner, I. Prolonged summer droughts retard soil N processing and stabilization in organo-mineral fractions. Soil Biol. Biochem. 2014, 68, 241–251. [Google Scholar] [CrossRef]
- Gessler, A.; Schrempp, S.; Matzarakis, A.; Mayer, H.; Rennenberg, H.; Adams, M.A. Radiation modifies the effect of water availability on the carbon isotope composition of beech (Fagus sylvatica). New Phytol. 2001, 150, 653–664. [Google Scholar] [CrossRef]
- Fotelli, M.N.; Nahm, M.; Heidenfelder, A.; Papen, H.; Rennenberg, H.; Geßler, A. Soluble nonprotein nitrogen compounds indicate changes in the nitrogen status of beech seedlings due to climate and thinning. New Phytol. 2002, 154, 85–97. [Google Scholar] [CrossRef]
- Keitel, C.; Adams, M.A.; Holst, T.; Matzarakis, A.; Mayer, H.; Rennenberg, H.; Gessler, A. Carbon and oxygen isotope composition of organic compounds in the phloem sap provides a short-term measure for stomatal conductance of European beech (Fagus sylvatica L.). Plant Cell Environ. 2003, 26, 1157–1168. [Google Scholar] [CrossRef]
- Nahm, M.; Holst, T.; Matzarakis, A.; Mayer, H.; Rennenberg, H.; Geßler, A. Soluble N compound profiles and concentrations in European beech (Fagus sylvatica L.) are influenced by local climate and thinning. Eur. J. For. Res. 2005, 125, 1–14. [Google Scholar] [CrossRef]
- Holst, T.; Mayer, H.; Schindler, D. Microclimate within beech stands–Part II: Thermal conditions. Eur. J. For. Res. 2004, 123, 13–28. [Google Scholar] [CrossRef]
- Fotelli, M.N.; Rennenberg, H.; Holst, T.; Mayer, H.; Gessler, A. Carbon isotope composition of various tissues of beech (Fagus sylvatica) regeneration is indicative of recent environmental conditions within the forest understorey. New Phytol. 2003, 159, 229–244. [Google Scholar] [CrossRef]
- Saiz, G.; Bird, M.I.; Domingues, T.; Schrodt, F.; Schwarz, M.; Feldpausch, T.R.; Veenendaal, E.; Djagbletey, G.; Hien, F.; Compaore, H.; et al. Variation in soil carbon stocks and their determinants across a precipitation gradient in West Africa. Glob. Chang. Biol. 2012, 18, 1670–1683. [Google Scholar] [CrossRef]
- Saiz, G.; Albrecht, A. Methods for smallholder quantification of soil carbon stocks and stock changes. In Measurement Methods Standard Assessment of Agricultural Mitigation Potential And Livelihoods; Rosenstock, T.S., Rufino, M.C., Butterbach-Bahl, K., Wollenberg, E., Richards, M., Eds.; Springer: New York, NY, USA, 2016. [Google Scholar]
- Walthert, L.; Graf, U.; Kammer, A.; Luster, J.; Pezzotta, D.; Zimmermann, S.; Hagedorn, F. Determination of organic and inorganic carbon, δ13C, and nitrogen in soils containing carbonates after acid fumigation with HCl. J. Plant Nutr. Soil Sci. 2010, 173, 207–216. [Google Scholar] [CrossRef]
- Harris, D.; Horwáth, W.R.; van Kessel, C. Acid fumigation of soils to remove carbonates prior to total organic carbon or carbon-13 isotopic analysis. Soil Sci. Soc. Am. J. 2001, 65, 1853. [Google Scholar] [CrossRef]
- Liu, M.; Dannenmann, M.; Lin, S.; Saiz, G.; Yan, G.; Yao, Z.; Pelster, D.E.; Tao, H.; Sippel, S.; Tao, Y.; et al. Ground cover rice production systems increase soil carbon and nitrogen stocks at regional scale. Biogeosciences 2015, 12, 4831–4840. [Google Scholar] [CrossRef]
- Jobbagy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Meier, I.C.; Leuschner, C. Variation of soil and biomass carbon pools in beech forests across a precipitation gradient. Glob. Chang. Biol. 2010, 16, 1035–1045. [Google Scholar] [CrossRef]
- Guckland, A.; Jacob, M.; Flessa, H.; Thomas, F.M.; Leuschner, C. Acidity, nutrient stocks, and organic-matter content in soils of a temperate deciduous forest with different abundance of European beech (Fagus sylvatica L.). J. Plant Nutr. Soil Sci. 2009, 172, 500–511. [Google Scholar] [CrossRef]
- De Clercq, T.; Heiling, M.; Dercon, G.; Resch, C.; Aigner, M.; Mayer, L.; Mao, Y.; Elsen, A.; Steier, P.; Leifeld, J.; et al. Predicting soil organic matter stability in agricultural fields through carbon and nitrogen stable isotopes. Soil Biol. Biochem. 2015, 88, 29–38. [Google Scholar] [CrossRef]
- Diochon, A.; Kellman, L. Natural abundance measurements of 13C indicate increased deep soil carbon mineralization after forest disturbance. Geophys. Res. Lett. 2008, 35, 1–5. [Google Scholar] [CrossRef]
- Van der Merwe, N.J.; Medina, E. Photosynthesis and 13C:12C ratios in Amazonian rain forests. Geochim. Cosmochim. Acta 1989, 53, 1091–1094. [Google Scholar] [CrossRef]
- Medina, E.; Minchin, P. Stratification of δ13C values of leaves in Amazonian rain forests. Oecologia 1980, 45, 377–378. [Google Scholar] [CrossRef] [PubMed]
- Ometto, J.P.H.B.; Ehleringer, J.R.; Domingues, T.F.; Berry, J.A.; Ishida, F.Y.; Mazzi, E.; Higuchi, N.; Flanagan, L.B.; Nardoto, G.B.; Martinelli, L.A. The stable carbon and nitrogen isotopic composition of vegetation in tropical forests of the Amazon Basin, Brazil. Biogeochemistry 2006, 79, 251–274. [Google Scholar] [CrossRef]
- Rennenberg, H.; Dannenmann, M. Nitrogen nutrition of trees in temperate forests-the significance of nitrogen availability in the pedosphere and atmosphere. Forests 2015, 6, 2820–2835. [Google Scholar] [CrossRef]
- Cremer, M.; Kern, N.V.; Prietzel, J. Soil organic carbon and nitrogen stocks under pure and mixed stands of European beech, Douglas fir and Norway spruce. For. Ecol. Manag. 2016, 367, 30–40. [Google Scholar] [CrossRef]
- Bauhus, J.; Vor, T.; Bartsch, N.; Cowling, A. The effects of gaps and liming on forest floor decomposition and soil C and N dynamics in a Fagus sylvatica forest. Can. J. For. Res. 2004, 34, 509–518. [Google Scholar] [CrossRef]
- Stevenson, B.A.; Parfitt, R.L.; Schipper, L.A.; Baisden, W.T.; Mudge, P. Relationship between soil δ15N, C/N and N losses across land uses in New Zealand. Agric. Ecosyst. Environ. 2010, 139, 736–741. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil | Leaf Litter | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Depth (cm) | Bulk Density (gr cm−3) | Soil Mass (t dry soil ha−1) | SOC Stock (t SOC ha−1) | SOC Concentration (gr C Kg dry soil−1) | TN Stocks (t N ha−1) | TN Concentration (gr N Kg dry soil−1) | Litter Mass (t ha−1) | SOC Stocks (t C ha−1) | SOC Concentration (gr C Kg dry mass−1) | TN Stocks (t N ha−1) | TN Concentration (gr N Kg dry soil−1) |
| NE 0–5 | 0.82 ± 0.03 | 311.3 ±1 8.7 | 25.4 ± 2.3 | 81.83 ± 7.50 | 1.90 ± 0.20 | 6.03 ± 0.60 | 15.5 ± 0.8 | 6.6 ± 0.4 | 423.7 ± 2.3 | 0.30 ± 0.02 | 17.69 ± 0.40 |
| NE 5–10 | 0.96 ± 0.04 | 264.5 ± 29.1 | 16.5 ± 2.4 | 62.3 ± 9.00 | 1.30 ± 0.20 | 4.97 ± 0.70 | |||||
| NW 0–5 | 0.82 ± 0.03 | 328.2 ± 13.1 | 26.8 ± 0.5 | 81.37 ± 1.60 | 1.90 ± 0.05 | 5.91 ± 0.20 | 16.0 ± 0.8 | 6.8 ± 0.4 | 423.4 ± 4.9 | 0.30 ± 0.02 | 17.05 ± 0.60 |
| NW 5–10 | 0.89 ± 0.04 | 279.0 ± 16.7 | 18.5 ± 0.9 | 66.36 ± 3.10 | 1.60 ± 0.20 | 5.80 ± 0.90 | |||||
| SW 0–5 | 0.68 ± 0.03 | 265.1 ± 13.2 | 32.9 ± 3 | 124.13 ± 11.00 | 1.90 ± 0.10 | 7.34 ± 0.40 | 11.6 ± 1.2 | 5.0 ± 0.5 | 433.6 ± 2.7 | 0.20 ± 0.02 | 13.68 ± 0.60 |
| SW 5–10 | 0.90 ± 0.05 | 185.6 ± 14.8 | 15.6 ± 0.4 | 84.00 ± 1.90 | 1.20 ± 0.08 | 6.67 ± 0.40 | |||||
| Depth (cm) | SOC Stocks | TN Stocks | C/N Ratio | δ15N | δ13C | |
|---|---|---|---|---|---|---|
| Soil | 0–5 | 0.093 | 0.541 | 0.041↑ | 0.875 | 0.603 |
| 5–10 | 0.878 | 0.959 | 0.388 | 0.456 | 0.209 | |
| Leaf litter | 0.028↓ | 0.028↓ | 0.028↑ | 0.072 | 0.027↑ |
| SOC Stocks | TN Stocks | C/N Ratio | δ15N | δ13C | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Thinning | Depth | Thinning | Depth | Thinning | Depth | Thinning | Depth | Thinning | Depth | |||
| NE | Soil | 0–5 | 0.068 | 0.005↓ | 0.144 | 0.005↓ | 0.753 | 0.005 | 0.917 | 0.002↑ | 0.028↓ | 0.002↑ |
| 5–10 | 0.144 | 0.068 | 0.345 | 0.753 | 0.345 | |||||||
| Litter | 0.593 | 1.000 | 1.000 | 0.285 | 0.414 | |||||||
| NW | Soil | 0–5 | 0.345 | 0.002↓ | 0.753 | 0.034↓ | 0.833 | 0.084 | 0.293 | 0.034↑ | 0.173 | 0.002↑ |
| 5–10 | 0.116 | 0.6 | 0.028↓ | 0.753 | 0.116 | |||||||
| Litter | 1.000 | 1.000 | 0.109 | 0.109 | 0.180 | |||||||
| SW | Soil | 0–5 | 0.463 | 0.003↓ | 0.173 | 0.002↓ | 0.917 | 0.182 | 0.345 | 0.005↑ | 0.345 | 0.023↑ |
| 5–10 | 0.249 | 0.345 | 0.173 | 0.249 | 0.753 | |||||||
| Litter | 0.285 | 0.593 | 1.000 | 0.180 | 1.000 | |||||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tejedor, J.; Saiz, G.; Rennenberg, H.; Dannenmann, M. Thinning of Beech Forests Stocking on Shallow Calcareous Soil Maintains Soil C and N Stocks in the Long Run. Forests 2017, 8, 167. https://doi.org/10.3390/f8050167
Tejedor J, Saiz G, Rennenberg H, Dannenmann M. Thinning of Beech Forests Stocking on Shallow Calcareous Soil Maintains Soil C and N Stocks in the Long Run. Forests. 2017; 8(5):167. https://doi.org/10.3390/f8050167
Chicago/Turabian StyleTejedor, Javier, Gustavo Saiz, Heinz Rennenberg, and Michael Dannenmann. 2017. "Thinning of Beech Forests Stocking on Shallow Calcareous Soil Maintains Soil C and N Stocks in the Long Run" Forests 8, no. 5: 167. https://doi.org/10.3390/f8050167
APA StyleTejedor, J., Saiz, G., Rennenberg, H., & Dannenmann, M. (2017). Thinning of Beech Forests Stocking on Shallow Calcareous Soil Maintains Soil C and N Stocks in the Long Run. Forests, 8(5), 167. https://doi.org/10.3390/f8050167