Long Non-Coding RNAs Responsive to Witches’ Broom Disease in Paulownia tomentosa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Library Construction and Sequencing
2.3. Genome Mapping and Transcript Assembly
2.4. LncRNAs Identification
2.5. Genomic Characterization and Function Prediction of lncRNAs
2.6. Identification of PaWB-Related Genes and lncRNAs
2.7. Nested-PCR and Quantitative RT-PCR
3. Results
3.1. Changes in Morphology of the Four Samples
3.2. High-Throughput Sequencing and Differentially Expressed Genes Analysis
3.3. Genome-Wide Identification and Characterization of lncRNAs in P. tomentosa
3.4. Differential Expression of lncRNAs
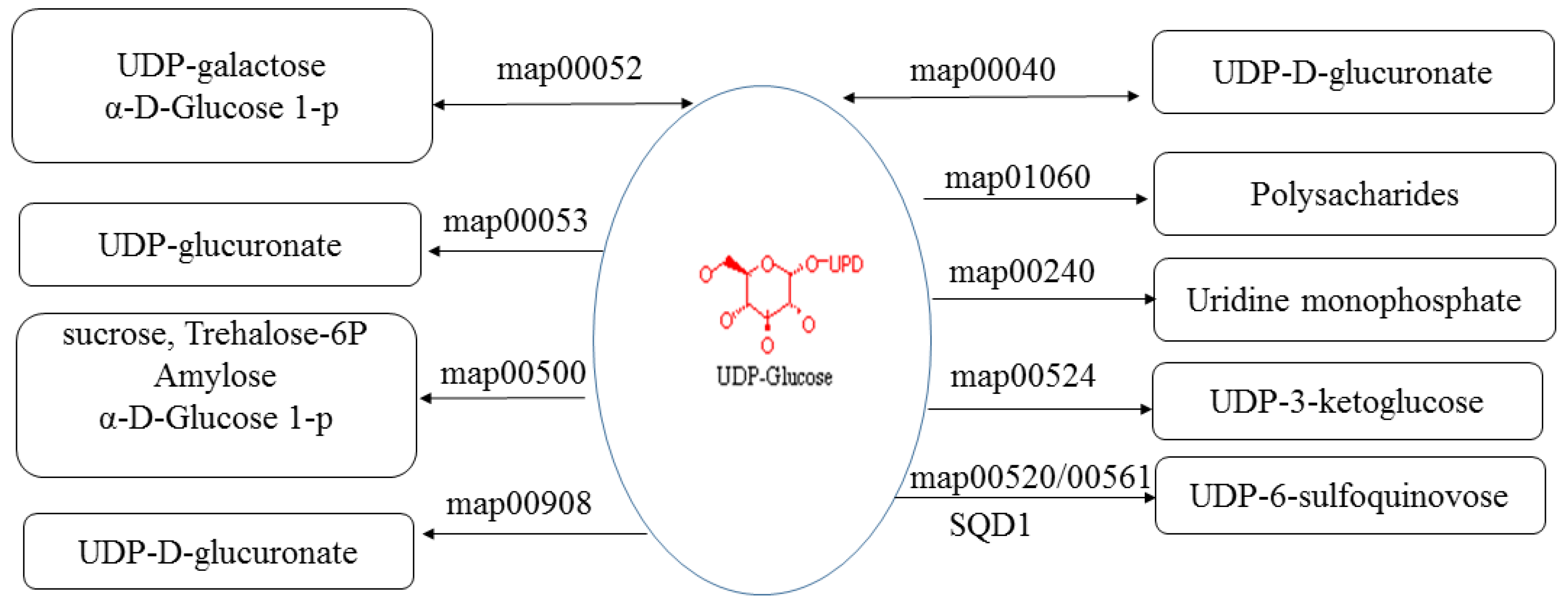
3.5. Prediction of lncRNA Candidate Target Genes
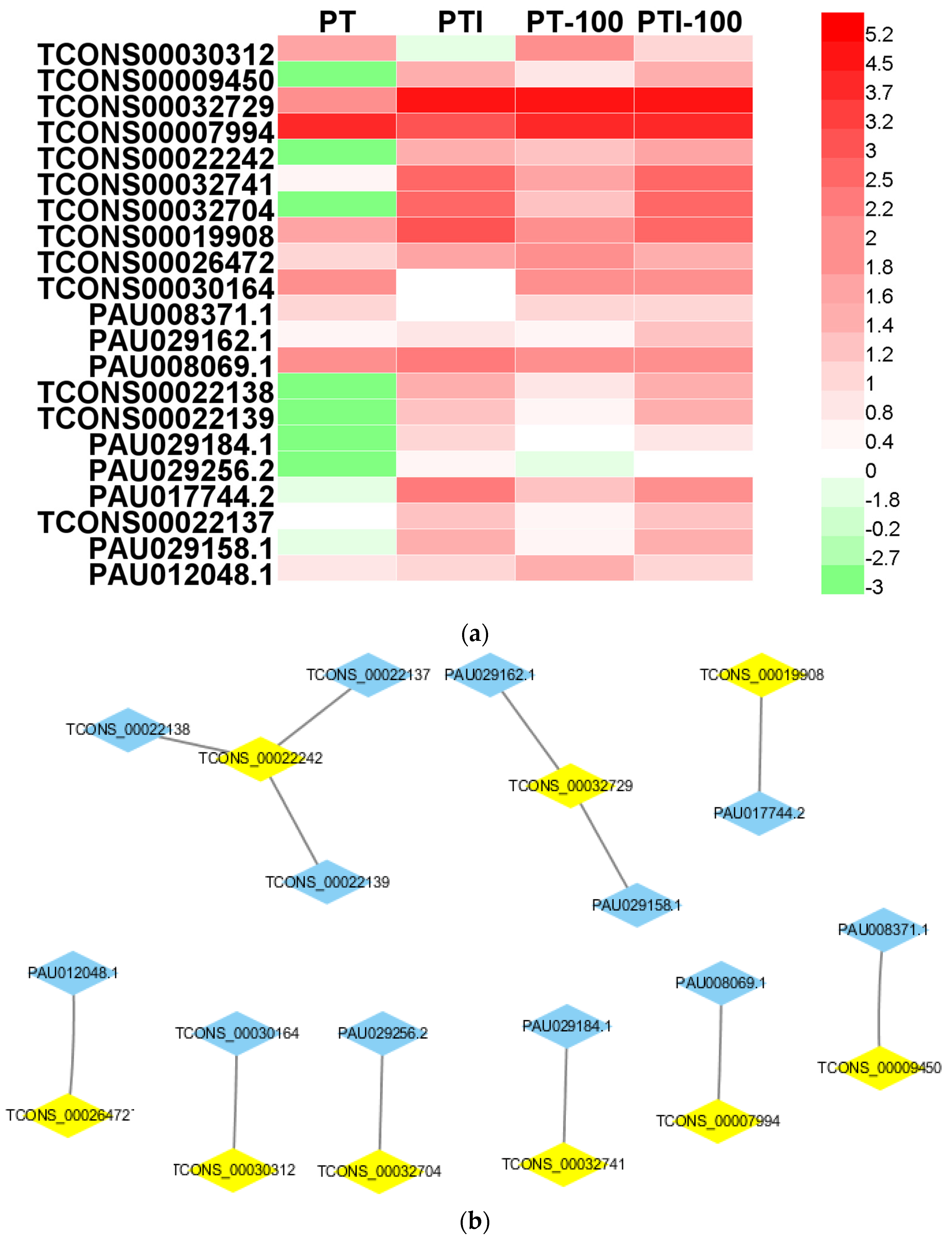
3.6. Analysis of PaWB-Related lncRNAs
3.7. Quantitative RT-PCR Analysis of DEGs and DELs
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fan, G.; Zhai, X.; Qin, H.; Yang, X.; Dong, Z.; Jiang, J. Study on In Vitro plantlet regeneration of paulownia plant with witches’ broom. J. Henan Agric. Univ. 2005, 39, 254–258. [Google Scholar]
- Hogenhout, S.A.; Oshima, K.; Ammar el, D.; Kakizawa, S.; Kingdom, H.N.; Namba, S. Phytoplasmas: Bacteria that manipulate plants and insects. Mol. Plant Pathol. 2008, 9, 403–423. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Dong, Y.; Deng, M.; Zhao, Z.; Niu, S.; Xu, E. Plant-pathogen interaction, circadian rhythm, and hormone-related gene expression provide indicators of phytoplasma infection in Paulownia fortunei. Int. J. Mol. Sci. 2014, 15, 23141–23162. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, W.; Fan, G.; Zhai, X.; Zhao, Z.; Dong, Y.; Deng, M.; Cao, Y. Quantitative proteome-level analysis of paulownia witches’ broom disease with methyl methane sulfonate assistance reveals diverse metabolic changes during the infection and recovery processes. PeerJ 2017, 5, e3495. [Google Scholar] [CrossRef] [PubMed]
- Ipekci, Z.; Gozukirmizi, N. Direct somatic embryogenesis and synthetic seed production from paulownia elongata. Plant Cell Rep. 2003, 22, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Fan, G.; Dong, Y.; Zhai, X.; Deng, M.; Zhao, Z.; Liu, W.; Cao, Y. Implications of polyploidy events on the phenotype, microstructure, and proteome of paulownia australis. PLoS ONE 2017, 12, e0172633. [Google Scholar] [CrossRef] [PubMed]
- Ju, G.; Wang, R.; Zhou, Y.; Dajin, R. Study on the relationship between the content of vitamin C and resistance of paulownia spp. To witches’ broom. For. Res. 1996, 9, 421–424. [Google Scholar]
- Wang, R.; Wang, S.; Sun, X. Effects of hormones on the morphogenesis of witches broom of Paulownia spp. Sci. Silvae Sin. 1981, 17, 281–286. [Google Scholar]
- Yue, H.; Wu, K.; Wu, Y.; Zhang, J.; Sun, R. Cloning and Characterization of Three Subunits of the Phytoplasma Sec Protein Translocation System Associated with the Paulownia Witches’-Broom. Plant Prot. 2009, 35, 25–31. [Google Scholar]
- Wang, J.; Zhu, X.; Gao, R.; Lin, C.; Li, Y.; Xu, Q.; Piao, C.; Li, X.; Li, H.; Tian, G. Genetic and serological analyses of elongation factor EF-TU of paulownia witches’-broom phytoplasma (16sri-d). Plant Pathol. 2010, 59, 972–981. [Google Scholar] [CrossRef]
- Lin, C.; Zhou, T.; Li, H.; Fan, Z.; Li, Y.; Piao, C.; Tian, G. Molecular characterisation of two plasmids from paulownia witches’-broom phytoplasma and detection of a plasmid-encoded protein in infected plants. Eur. J. Plant Pathol. 2008, 123, 321–330. [Google Scholar] [CrossRef]
- Du, T.; Wang, Y.; Hu, Q.; Chen, J.; Liu, S.; Huang, W.; Lin, M. Transgenic paulownia expressing shiva-1 gene has increased resistance to paulownia witches’ broom disease. J. Integr. Plant Biol. 2005, 47, 1500–1506. [Google Scholar] [CrossRef]
- Fan, G.; Zhang, B.; Zhai, X.; Liu, F.; Ma, Y.B.; Kan, S. Effects of rifampin on the changes of morphology and plant endogenous hormones of paulownia seedlings with witches’ broom. J. Henan Agric. Univ. 2007, 41, 387–391. [Google Scholar]
- Fan, G.; Cao, X.; Niu, S.; Deng, M.; Zhao, Z.; Dong, Y. Transcriptome, microrna, and degradome analyses of the gene expression of paulownia with phytoplamsa. BMC Genom. 2015, 16, 896. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Cao, X.; Zhao, Z.; Deng, M. Transcriptome analysis of the genes related to the morphological changes of paulownia tomentosa plantlets infected with phytoplasma. Acta Physiol. Plant. 2015, 37, 1–12. [Google Scholar] [CrossRef]
- Fan, G.; Xu, E.; Deng, M.; Zhao, Z.; Niu, S. Phenylpropanoid metabolism, hormone biosynthesis and signal transduction-related genes play crucial roles in the resistance of paulownia fortunei to paulownia witches’ broom phytoplasma infection. Genes Genom. 2015, 37, 913–929. [Google Scholar] [CrossRef]
- Liu, R.; Dong, Y.; Fan, G.; Zhao, Z.; Deng, M.; Cao, X.; Niu, S. Discovery of genes related to witches broom disease in paulownia tomentosa x paulownia fortunei by a de novo assembled transcriptome. PLoS ONE 2013, 8, e80238. [Google Scholar] [CrossRef] [PubMed]
- Mou, H.Q.; Lu, J.; Zhu, S.F.; Lin, C.L.; Tian, G.Z.; Xu, X.; Zhao, W.J. Transcriptomic analysis of paulownia infected by paulownia witches’-broom phytoplasma. PLoS ONE 2013, 8, e77217. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Cao, Y.; Deng, M.; Zhai, X.; Zhao, Z.; Niu, S.; Ren, Y. Identification and dynamic expression profiling of micrornas and target genes of paulownia tomentosa in response to paulownia witches’ broom disease. Acta Physiol. Plant. 2016, 39, 28. [Google Scholar] [CrossRef]
- Fan, G.; Niu, S.; Xu, T.; Deng, M.; Zhao, Z.; Wang, Y.; Cao, L.; Wang, Z. Plant-pathogen interaction-related micrornas and their targets provide indicators of phytoplasma infection in paulownia tomentosa x paulownia fortunei. PLoS ONE 2015, 10, e0140590. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Niu, S.; Zhao, Z.; Deng, M.; Xu, E.; Wang, Y.; Yang, L. Identification of micrornas and their targets in paulownia fortunei plants free from phytoplasma pathogen after methyl methane sulfonate treatment. Biochimie 2016, 127, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.; Fan, G.; Deng, M.; Zhao, Z.; Xu, E.; Cao, L. Discovery of micrornas and transcript targets related to witches’ broom disease in paulownia fortunei by high-throughput sequencing and degradome approach. Mol. Genet. Genom. 2016, 291, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Fan, G.; Dong, Y.; Zhao, Z.; Deng, M.; Wang, Z.; Liu, W. Proteome profiling of paulownia seedlings infected with phytoplasma. Front. Plant Sci. 2017, 8, 342. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [PubMed]
- Ben Amor, B.; Wirth, S.; Merchan, F.; Laporte, P.; d’Aubenton-Carafa, Y.; Hirsch, J.; Maizel, A.; Mallory, A.; Lucas, A.; Deragon, J.M.; et al. Novel long non-protein coding RNAs involved in arabidopsis differentiation and stress responses. Genome Res. 2009, 19, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jung, C.; Xu, J.; Wang, H.; Deng, S.; Bernad, L.; Arenas-Huertero, C.; Chua, N.H. Genome-wide analysis uncovers regulation of long intergenic noncoding RNAs in arabidopsis. Plant Cell 2012, 24, 4333–4345. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chung, P.J.; Liu, J.; Jang, I.C.; Kean, M.J.; Xu, J.; Chua, N.H. Genome-wide identification of long noncoding natural antisense transcripts and their responses to light in arabidopsis. Genome Res. 2014, 24, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Lu, Q.; Ouyang, Y.; Mao, H.; Zhang, P.; Yao, J.; Xu, C.; Li, X.; Xiao, J.; Zhang, Q. A long noncoding rna regulates photoperiod-sensitive male sterility, an essential component of hybrid rice. Proc. Natl. Acad. Sci. USA 2012, 109, 2654–2659. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.C.; Liao, J.Y.; Li, Z.Y.; Yu, Y.; Zhang, J.P.; Li, Q.F.; Qu, L.H.; Shu, W.S.; Chen, Y.Q. Genome-wide screening and functional analysis identify a large number of long noncoding RNAs involved in the sexual reproduction of rice. Genome Biol. 2014, 15, 512. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.; Wang, Y.; Yao, Y.; Song, N.; Hu, Z.; Qin, D.; Xie, C.; Peng, H.; Ni, Z.; Sun, Q. Identification and characterization of wheat long non-protein coding RNAs responsive to powdery mildew infection and heat stress by using microarray analysis and SBS sequencing. BMC Plant Biol. 2011, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, X.; Wang, C.; Xu, Z.; Wang, Y.; Liu, X.; Kang, Z.; Ji, W. Long non-coding genes implicated in response to stripe rust pathogen stress in wheat (Triticum aestivum L.). Mol. Biol. Rep. 2013, 40, 6245–6253. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Hao, Z.; Yan, J.; Li, G. Genome-wide identification and functional analysis of lincrnas acting as mirna targets or decoys in maize. BMC Genom. 2015, 16, 793. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Chen, X.; Mu, M.; Wang, J.; Wang, X.; Wang, D.; Yin, Z.; Fan, W.; Wang, S.; Guo, L.; et al. Genome-wide analysis of long noncoding RNAs and their responses to drought stress in cotton (Gossypium hirsutum L.). PLoS ONE 2016, 11, e0156723. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yuan, D.; Tu, L.; Gao, W.; He, Y.; Hu, H.; Wang, P.; Liu, N.; Lindsey, K.; Zhang, X. Long noncoding RNAs and their proposed functions in fibre development of cotton (Gossypium spp.). New Phytol. 2015, 207, 1181–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, R.K.; Megha, S.; Basu, U.; Rahman, M.H.; Kav, N.N. Genome wide identification and functional prediction of long non-coding RNAs responsive to sclerotinia sclerotiorum infection in Brassica napus. PLoS ONE 2016, 11, e0158784. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Song, Y.; Du, Q.; Yang, X.; Ci, D.; Chen, J.; Xie, J.; Li, B.; Zhang, D. Population genomic analysis of gibberellin-responsive long non-coding RNAs in populus. J. Exp. Bot. 2016, 67, 2467–2482. [Google Scholar] [CrossRef] [PubMed]
- Shuai, P.; Liang, D.; Tang, S.; Zhang, Z.; Ye, C.Y.; Su, Y.; Xia, X.; Yin, W. Genome-wide identification and functional prediction of novel and drought-responsive lincrnas in populus trichocarpa. J. Exp. Bot. 2014, 65, 4975–4983. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.H.; Stephen, S.; Taylor, J.; Helliwell, C.A.; Wang, M.B. Long noncoding RNAs responsive to fusarium oxysporum infection in Arabidopsis thaliana. New Phytol. 2014, 201, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, W.; Yang, Y.; Li, X.; Chen, T.; Liu, T.; Ma, N.; Yang, X.; Liu, R.; Zhang, B. Genome-wide analysis of tomato long non-coding RNAs and identification as endogenous target mimic for microrna in response to tylcv infection. Sci. Rep. 2015, 5, 16946. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Luan, Y.; Jiang, N.; Bao, H.; Meng, J. Comparative transcriptome analysis between resistant and susceptible tomato allows the identification of lncrna16397 conferring resistance to phytophthora infestans by co-expressing glutaredoxin. Plant J. 2017, 89, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. Tophat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with tophat and cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhang, Z.; Bailey, T.L.; Perkins, A.C.; Tallack, M.R.; Xu, Z.; Liu, H. Prediction of novel long non-coding RNAs based on RNA-Seq data of mouse klf1 knockout study. BMC Bioinform. 2012, 13, 331. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, Y.; Chen, M.; Xiao, P.; Hu, C.; Zeng, Z.; Wang, C.; Wang, J.; Hu, Z. Genome-wide long non-coding RNA screening, identification and characterization in a model microorganism chlamydomonas reinhardtii. Sci. Rep. 2016, 6, 34109. [Google Scholar] [CrossRef] [PubMed]
- Nawrocki, E.P.; Kolbe, D.L.; Eddy, S.R. Infernal 1.0: Inference of rna alignments. Bioinformatics 2009, 25, 1335–1337. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Osak, M.; Bogu, G.K.; Stanton, L.W.; Johnson, R.; Lipovich, L. Genome-wide computational identification and manual annotation of human long noncoding RNA genes. RNA 2010, 16, 1478–1487. [Google Scholar] [CrossRef] [PubMed]
- Tafer, H.; Hofacker, I.L. Rnaplex: A fast tool for RNA-RNA interaction search. Bioinformatics 2008, 24, 2657–2663. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta c(t)) method. Methods (San Diego, Calif.) 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Van Sandt, V.S.; Guisez, Y.; Verbelen, J.P.; Vissenberg, K. Analysis of a xyloglucan endotransglycosylase/hydrolase (xth) from the lycopodiophyte selaginella kraussiana suggests that xth sequence characteristics and function are highly conserved during the evolution of vascular plants. J. Exp. Bot. 2006, 57, 2909–2922. [Google Scholar] [CrossRef] [PubMed]
- Osato, Y.; Yokoyama, R.; Nishitani, K. A principal role for atxth18 in Arabidopsis thaliana root growth: A functional analysis using rnai plants. J. Plant Res. 2006, 119, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.E.; Fry, S.C. Restructuring of wall-bound xyloglucan by transglycosylation in living plant cells. Plant J. 2001, 26, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Shimojima, M.; Hoffmann-Benning, S.; Garavito, R.M.; Benning, C. Ferredoxin-dependent glutamate synthase moonlights in plant sulfolipid biosynthesis by forming a complex with SQD1. Arch. Biochem. Biophys. 2005, 436, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Kliebenstein, D.J. Secondary metabolites and plant/environment interactions: A view through Arabidopsis thaliana tinged glasses. Plant Cell Environ. 2004, 27, 675–684. [Google Scholar] [CrossRef]
- Litvak, M.E.; Monson, R.K. Patterns of induced and constitutive monoterpene production in conifer needles in relation to insect herbivory. Oecologia 1998, 114, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Ringer, K.L.; McConkey, M.E.; Davis, E.M.; Rushing, G.W.; Croteau, R. Monoterpene double-bond reductases of the (−)-menthol biosynthetic pathway: Isolation and characterization of cdnas encoding (−)-isopiperitenone reductase and (+)-pulegone reductase of peppermint. Arch. Biochem. Biophys. 2003, 418, 80–92. [Google Scholar] [CrossRef]
- Choi, H.W.; Lee, B.G.; Kim, N.H.; Park, Y.; Lim, C.W.; Song, H.K.; Hwang, B.K. A role for a menthone reductase in resistance against microbial pathogens in plants. Plant Physiol. 2008, 148, 383–401. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, E.J.; Peacock, W.J.; Dennis, E.S. DNA methylation, a key regulator of plant development and other processes. Curr. Opin. Genet. Dev. 2000, 10, 217–223. [Google Scholar] [CrossRef]
- Bestor, T.H.; Verdine, G.L. DNA methyltransferases. Curr. Opin. Cell Biol. 1994, 6, 380–389. [Google Scholar] [CrossRef]
- Cao, X.; Fan, G.; Deng, M.; Zhao, Z.; Dong, Y. Identification of genes related to paulownia witches’ broom by AFLP and MSAP. Int. J. Mol. Sci. 2014, 15, 14669–14683. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Fan, G.; Zhao, Z.; Deng, M.; Dong, Y. Morphological changes of paulownia seedlings infected phytoplasmas reveal the genes associated with witches’ broom through aflp and msap. PLoS ONE 2014, 9, e112533. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PT | PTI | PT-100 | PTI-100 | |
|---|---|---|---|---|
| Raw reads | 53,370,668 | 53,369,943 | 41,161,819 | 47,621,768 |
| Clean reads | 52,474,513 | 52,726,949 | 40,258,456 | 46,657,407 |
| mRNAs | 26,155 | 27,017 | 26,334 | 26,826 |
| lncRNAs | 3434 | 3504 | 3520 | 3479 |
| mRNA | Description | lncRNA |
|---|---|---|
| PAU008069.1 | indol-3-yl-methylglucosinolate hydroxylase | TCONS_00007994 |
| PAU008371.1 | trans-2,3-dihydro-3-hydroxyanthranilate isomerase | TCONS_00009450 |
| PAU017744.2 | hypothetical protein | TCONS_00019908 |
| TCONS_00022138 | nucleoporin-like protein 2 | TCONS_00022242 |
| TCONS_00022139 | xyloglucan:xyloglucosyl transferase | |
| TCONS_00022137 | nucleoporin-like protein 2 | |
| PAU012048.1 | neomenthol dehydrogenase | TCONS_00026472 |
| TCONS_00030164 | uncharacterized protein | TCONS_00030312 |
| PAU029256.2 | DNA (cytosine-5)-methyltransferase | TCONS_00032704 |
| PAU029162.1 | hypothetical protein | TCONS_00032729 |
| PAU029158.1 | UDP-sulfoquinovose synthase | |
| PAU029184.1 | hypothetical protein | TCONS_00032741 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Zhai, X.; Cao, Y.; Dong, Y.; Fan, G. Long Non-Coding RNAs Responsive to Witches’ Broom Disease in Paulownia tomentosa. Forests 2017, 8, 348. https://doi.org/10.3390/f8090348
Wang Z, Zhai X, Cao Y, Dong Y, Fan G. Long Non-Coding RNAs Responsive to Witches’ Broom Disease in Paulownia tomentosa. Forests. 2017; 8(9):348. https://doi.org/10.3390/f8090348
Chicago/Turabian StyleWang, Zhe, Xiaoqiao Zhai, Yabing Cao, Yanpeng Dong, and Guoqiang Fan. 2017. "Long Non-Coding RNAs Responsive to Witches’ Broom Disease in Paulownia tomentosa" Forests 8, no. 9: 348. https://doi.org/10.3390/f8090348
APA StyleWang, Z., Zhai, X., Cao, Y., Dong, Y., & Fan, G. (2017). Long Non-Coding RNAs Responsive to Witches’ Broom Disease in Paulownia tomentosa. Forests, 8(9), 348. https://doi.org/10.3390/f8090348