1. Introduction
Net primary production (NPP) not only reflects biological properties and stand structures for a forest ecosystem, but also describes the influence of environmental factors on forest growth [
1]. Growth is substantially affected by CO
2 fertilization, climate variability, nitrogen (N) deposition [
2,
3], and disturbance factors, such as wildfires, harvesting, and insects [
4,
5,
6]. Disturbances are major determinants of forest carbon stocks and uptake. They generally reduce land carbon stocks but also initiate a regrowth legacy that contributes substantially to the contemporary rate of carbon stock increase in forestlands [
7].
The interannual variability of NPP is not only affected by climatic variability, tree species composition, and forest succession [
8], but is also closely related to forest age. Many previous studies have demonstrated that stand age impacts on NPP can be substantial at both local and national scales [
9,
10,
11]. For example, NPP rapidly increases in young forest stands, reaches a maximum when mature, and gradually declines in older stands [
12,
13,
14]. However, several recent studies have provided evidence that not all forest types exhibit this decline [
15,
16]. Additionally, NPP is not only affected by forest age, but also by factors related to environmental site conditions. Chen et al. [
17] showed that NPP increased faster and the value of NPP was higher at more productive sites. Hence, both quantitative analyses of the successional change of NPP along with age structure and an understanding of the relationships between NPP, age, and site conditions, are critically important for improving forest carbon cycle estimation.
Traditional forest inventories have provided a large amount of ground data on tree growth, including diameter at breast height (DBH) and tree height (H). Fang et al. [
18] used forest inventory data from 1949 to 1998 to estimate forest biomass and carbon (C) storage in China. Similarly, stand yield tables were developed and designed based on forest inventories, and contain the most readily available information regarding the effects of stand age on tree growth. Tree growth can be combined with stand yield tables to estimate the relationships between NPP and age for various site conditions. Due to averaging and smoothing procedures which are generally applied in yield table development, NPP-age relationships are averaged over long-term mean environmental conditions. Therefore, this method can produce mean NPP-age relationships [
17].
Biogeochemical process models are effective tools used to help understand the response of forest ecosystems to various environmental drivers. The Integrated Terrestrial Ecosystem Carbon (InTEC) model is a process-based biogeochemical model produced by Chen et al. [
19] which integrates the effects of both disturbance and non-disturbance factors in long-term C budget simulations [
20,
21]. The model has been validated and used to simulate the historical carbon budget of forest environments. For instance, it was used to simulate the historical change of C dynamics and to analyze spatiotemporal carbon sink distributions in Canada [
22]. Then, the model was calibrated further for China [
3,
23]. Recently, the InTEC model has been used to estimate the C balance and distribution in North American forest ecosystems [
5,
6].
Studies have used a variety of methods to develop the NPP-age relationships for different forest types and regions. Chen et al. [
17] used yield tables to quantify mean age-NPP relationships in black spruce stands in Ontario, Canada. Wang et al. [
24] developed functions describing the relationship between national mean NPP and stand age for five typical forest ecosystems in China in 2001, using stand age information derived from forest inventory data and NPP simulated by BEPS (Boreal Ecosystem Productivity Simulator). Zaehle et al. [
25] analyzed the first-order effects of age on aboveground productivity of forests in Europe using a global terrestrial biosphere model (the Lund-Potsdam-Jena model, LPJ). He et al. [
26] developed NPP-age relationships for major forest biomes in the U.S. by combining data from the Forest Inventory and Analysis (FIA), remote sensing, and species-specific traits.
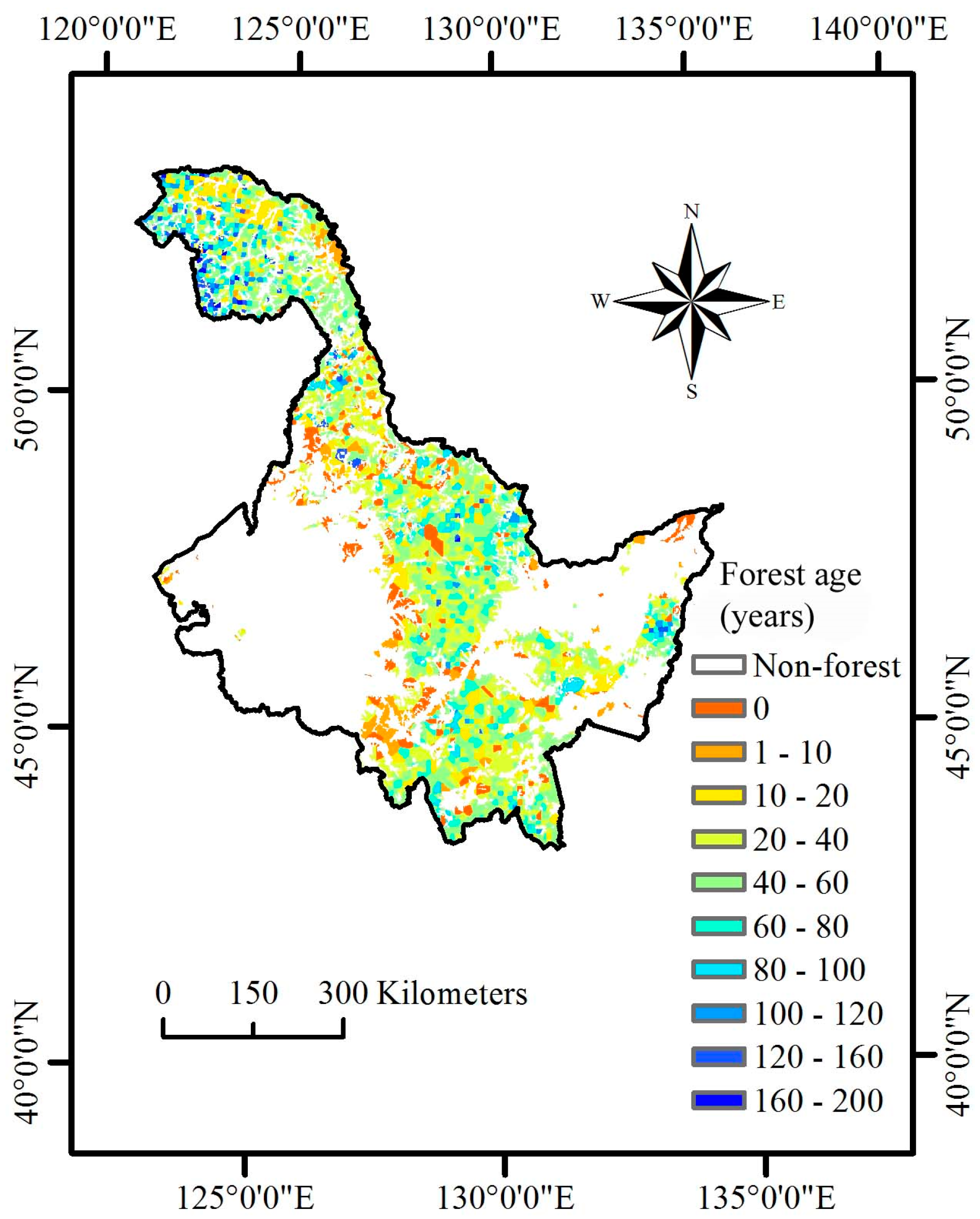
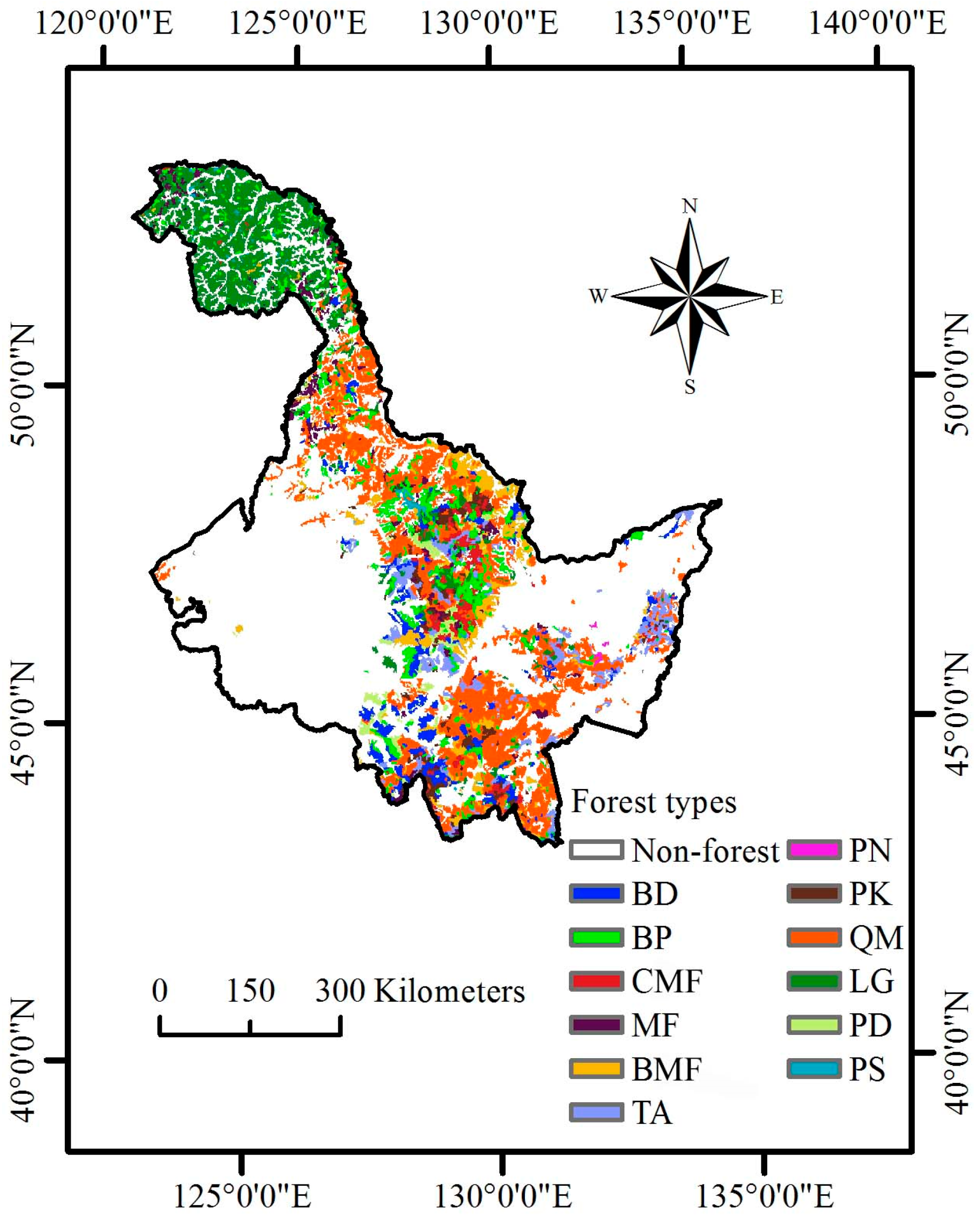
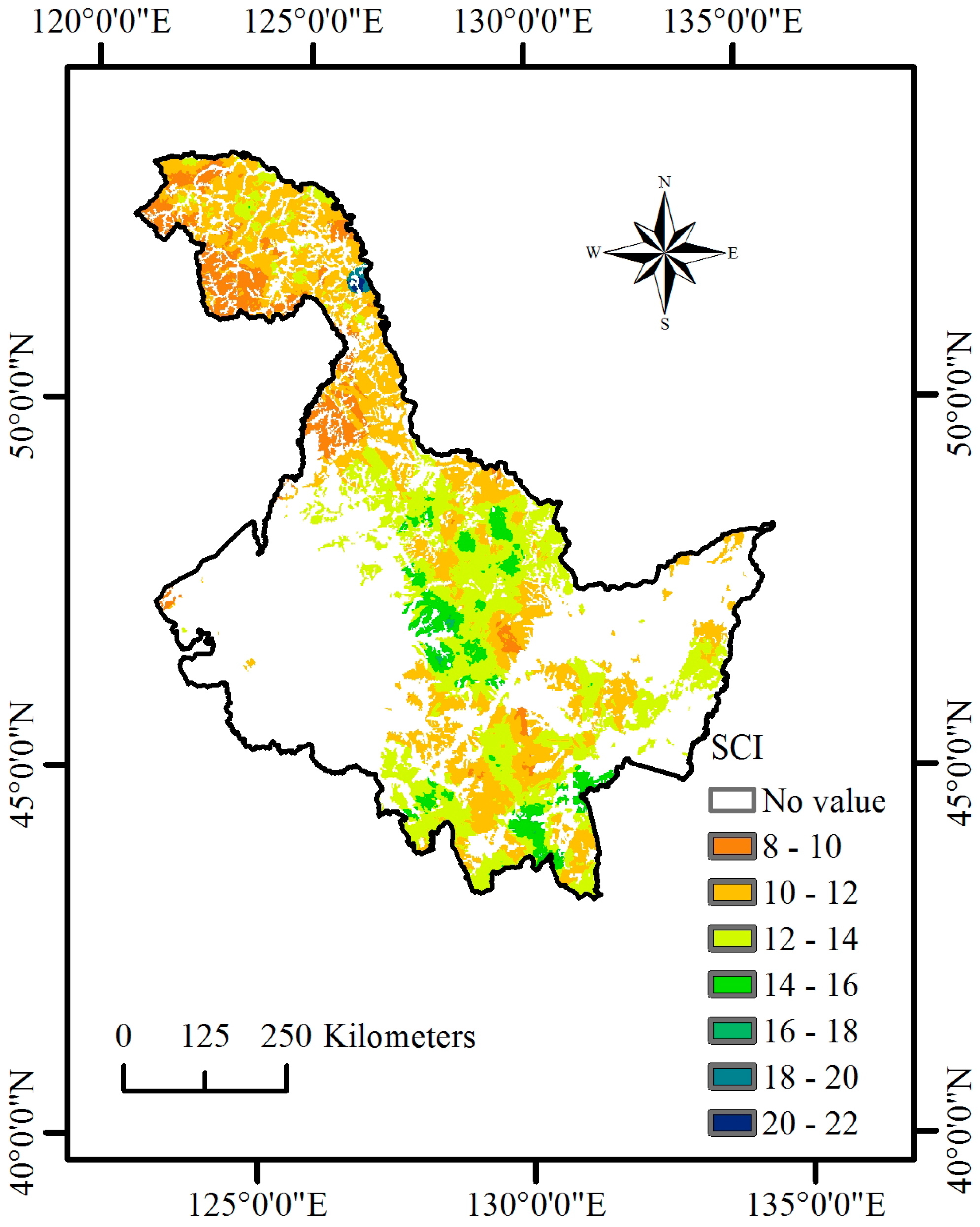
Until now, none of the existing regional C cycle modeling has incorporated systematic NPP-age relationships at various site class index (SCI) values in Heilongjiang Province, China. Heilongjiang Province (121°11′ E to 135°05′ E and 43°25′ N to 53°33′ N) is located in Northeast China (
Figure 1). The province has a total forest area of over 20 million ha, which is about 45.8% of the total land area of the province, and the forest coverage rate is 45.7%. The total volume of living trees is 1.76 billion m
3 and the average volume per hectare is 78.6 m
3. In addition, the forest fire hazard in Heilongjiang Province is the highest of any area in China [
27]. Therefore, the objectives of this study were to develop systematic NPP-age relationships at various SCI values for major forest types and groups in Heilongjiang Province, China, by combining data from forest inventories and stand yield tables, and to examine whether the new NPP-age relationships at various SCI values could estimate NPP through C cycle modeling.
5. Discussion
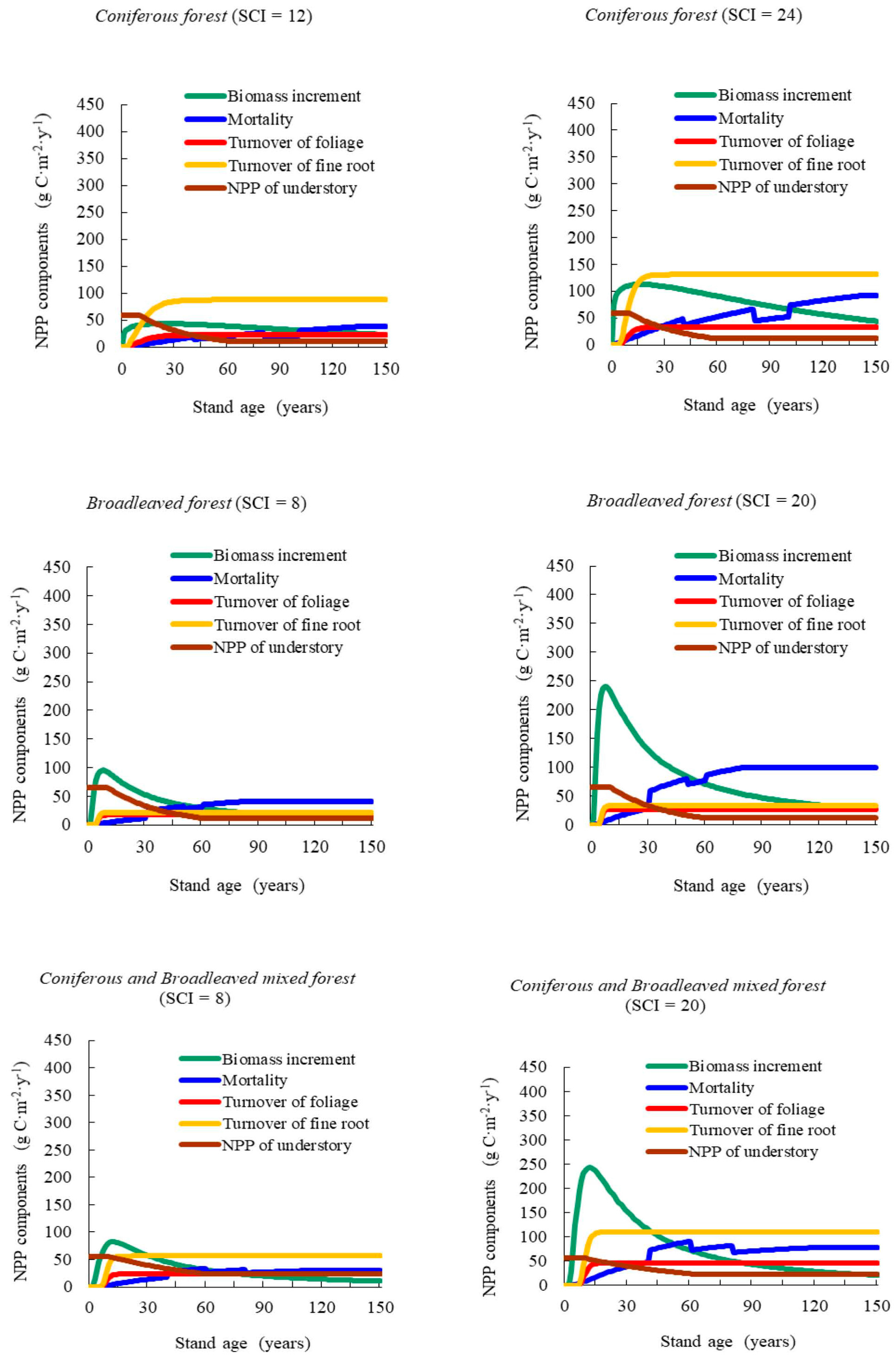
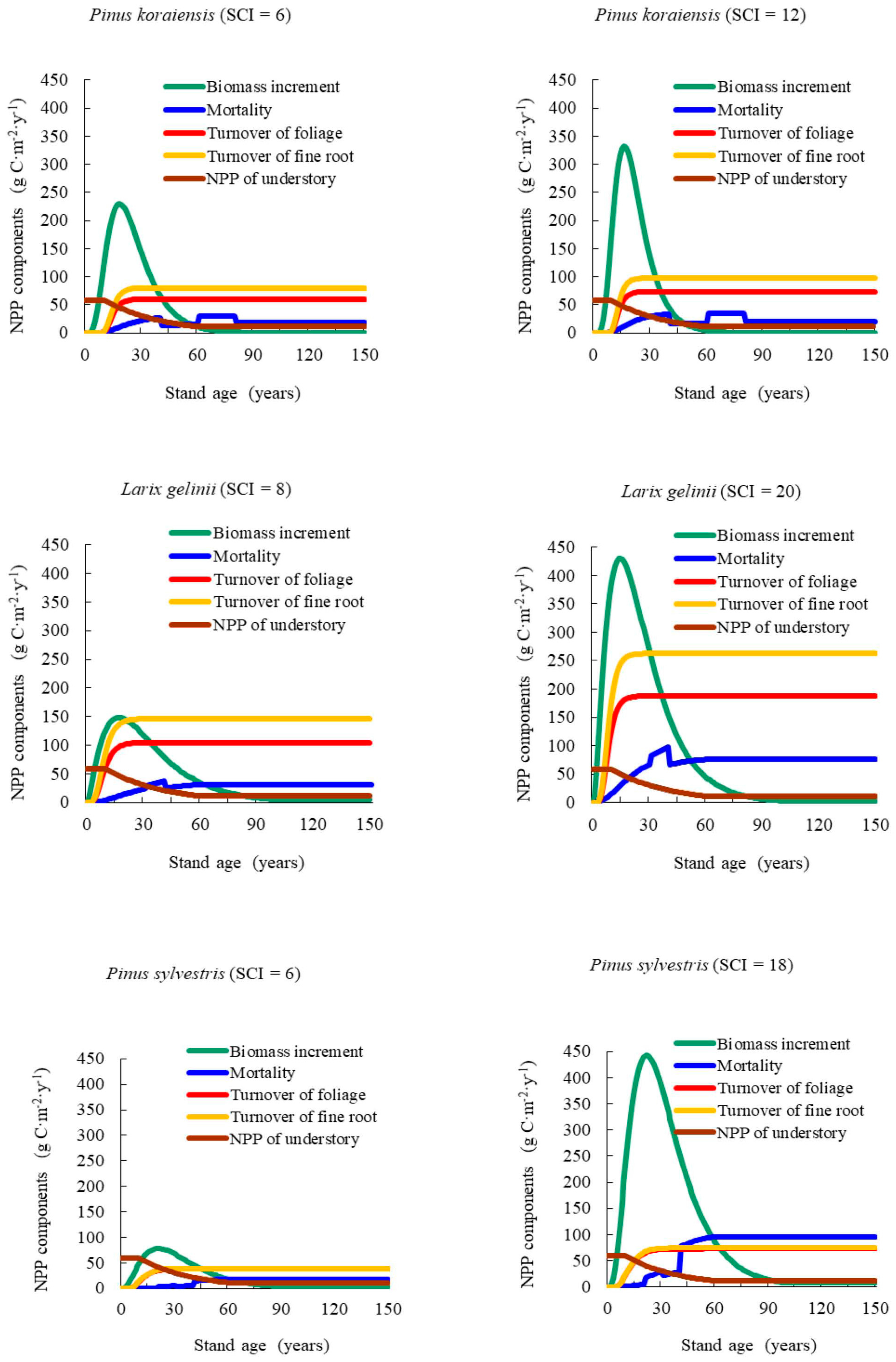
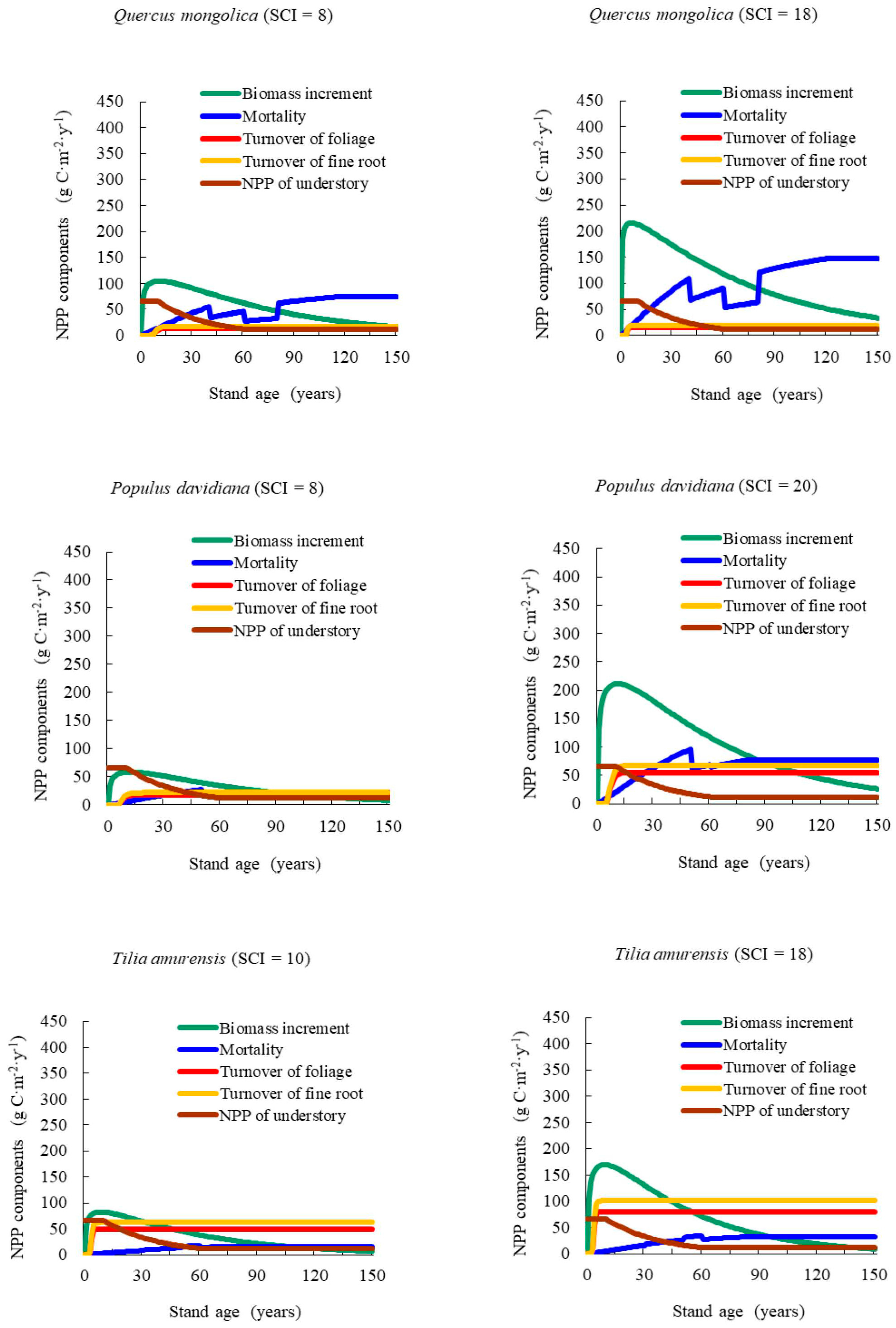
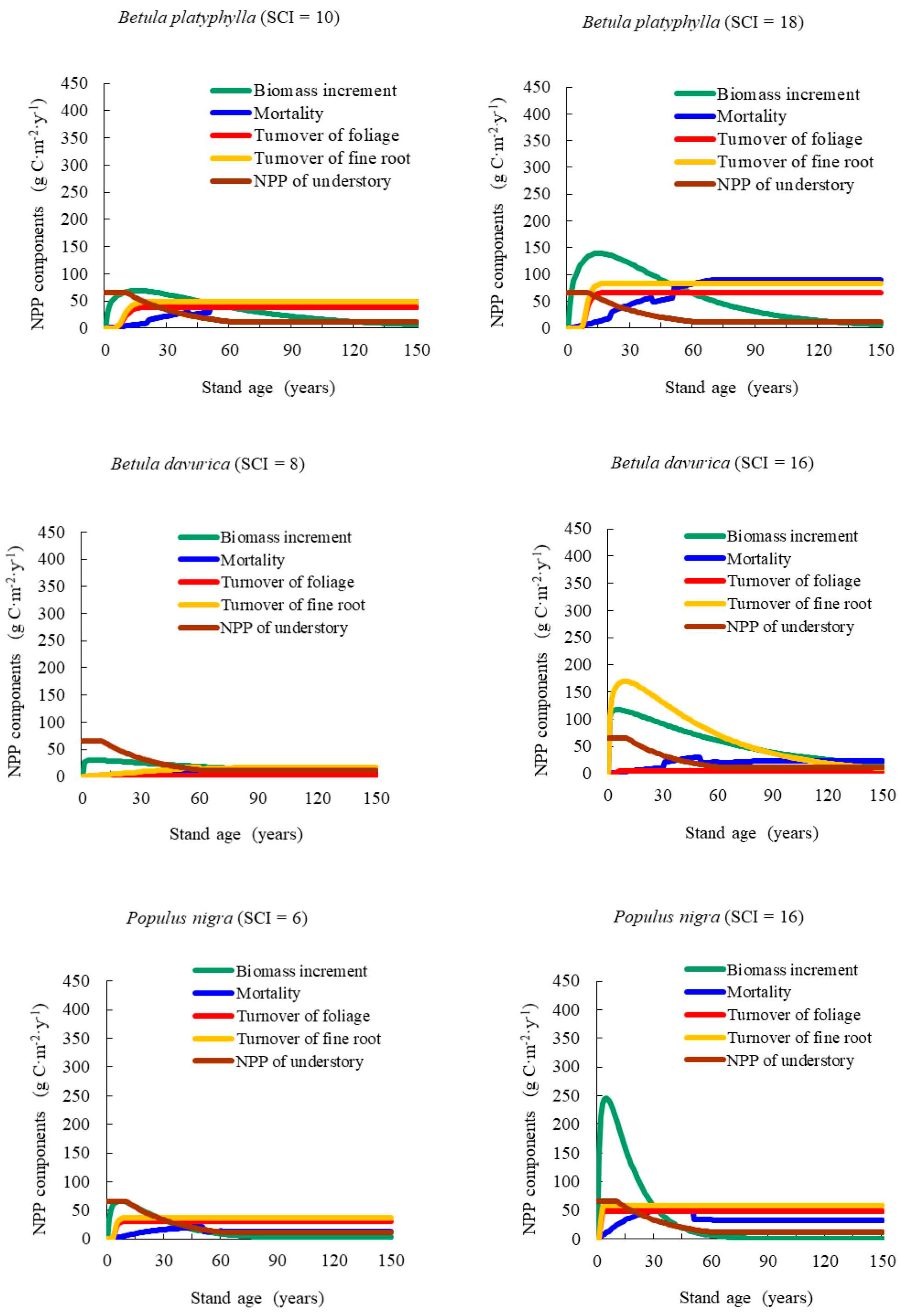
Our results showed that NPP varied with stand age and SCI, presenting a generally consistent trend of NPP-age relationships at various SCI values for different forest types and groups across the Heilongjiang Province, China. The NPP-age relationship curves at various SCI values were determined by components including the increase in total biomass, mortality, the fine root and foliage turnover rates, and the NPP of understory vegetation and moss. This method was similar in principle to that of Gower et al. [
51] and Bernier et al. [
52], who estimated stand-level NPP in a specific year using measured annual radial increased, biomass allometric equations, and measured foliage and root turnover rates. In the growth of young forests, NPP increasing rapidly with age was caused mainly by increases in biomass growth. Zha et al. [
14] reported that aboveground NPP (ANPP) increased with age, stabilizing after 25 years. These results aligned well with those of our study. For old stand forests, where the products of photosynthesis were mainly consumed by foliage and fine root turnover, NPP began to decrease. Ryan et al. [
13] reported that the range of NPP decreased by one half to one third from the peak growth period. Our results showed that it remained steady at 30% to 60% of the maximum NPP.
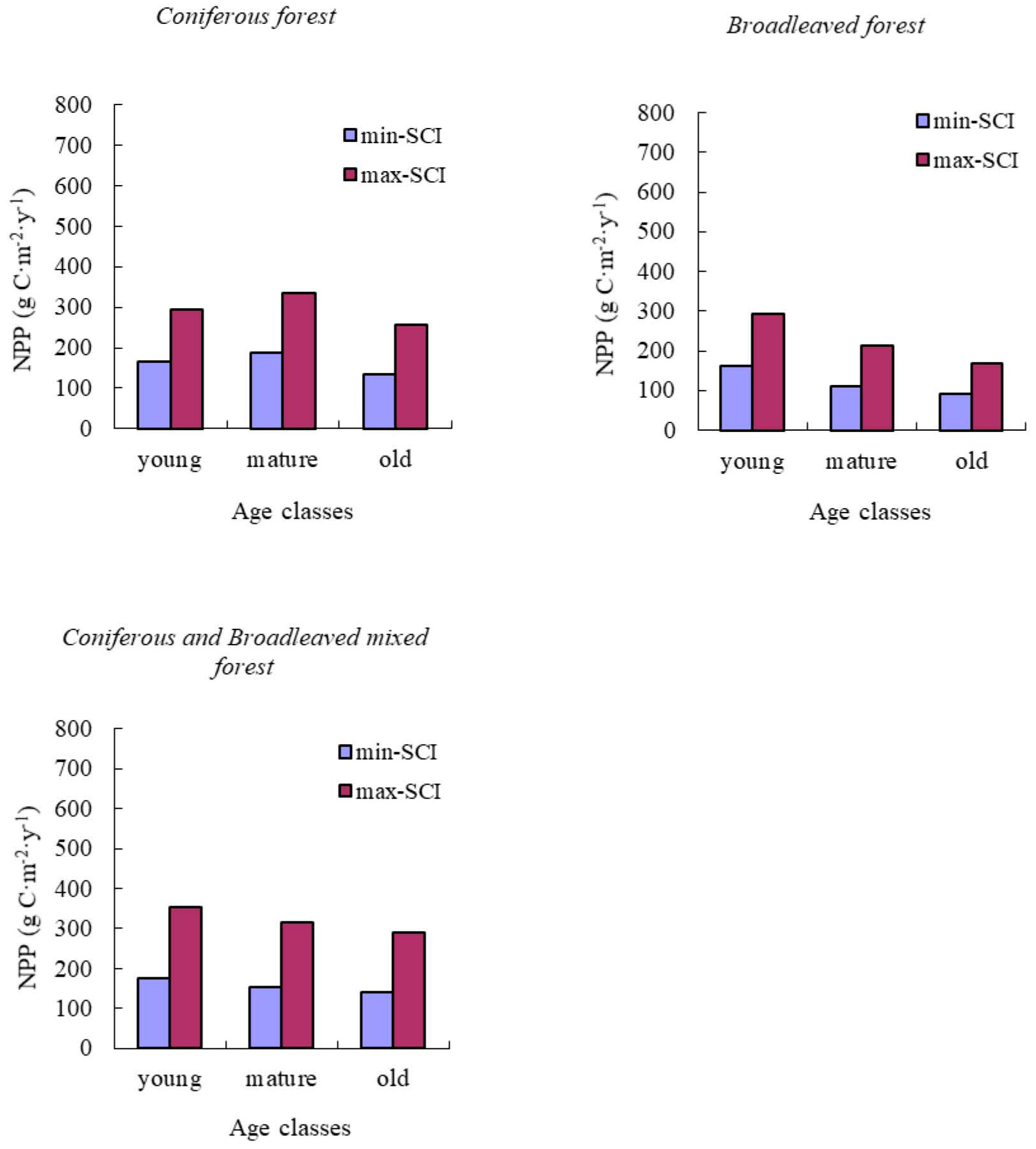
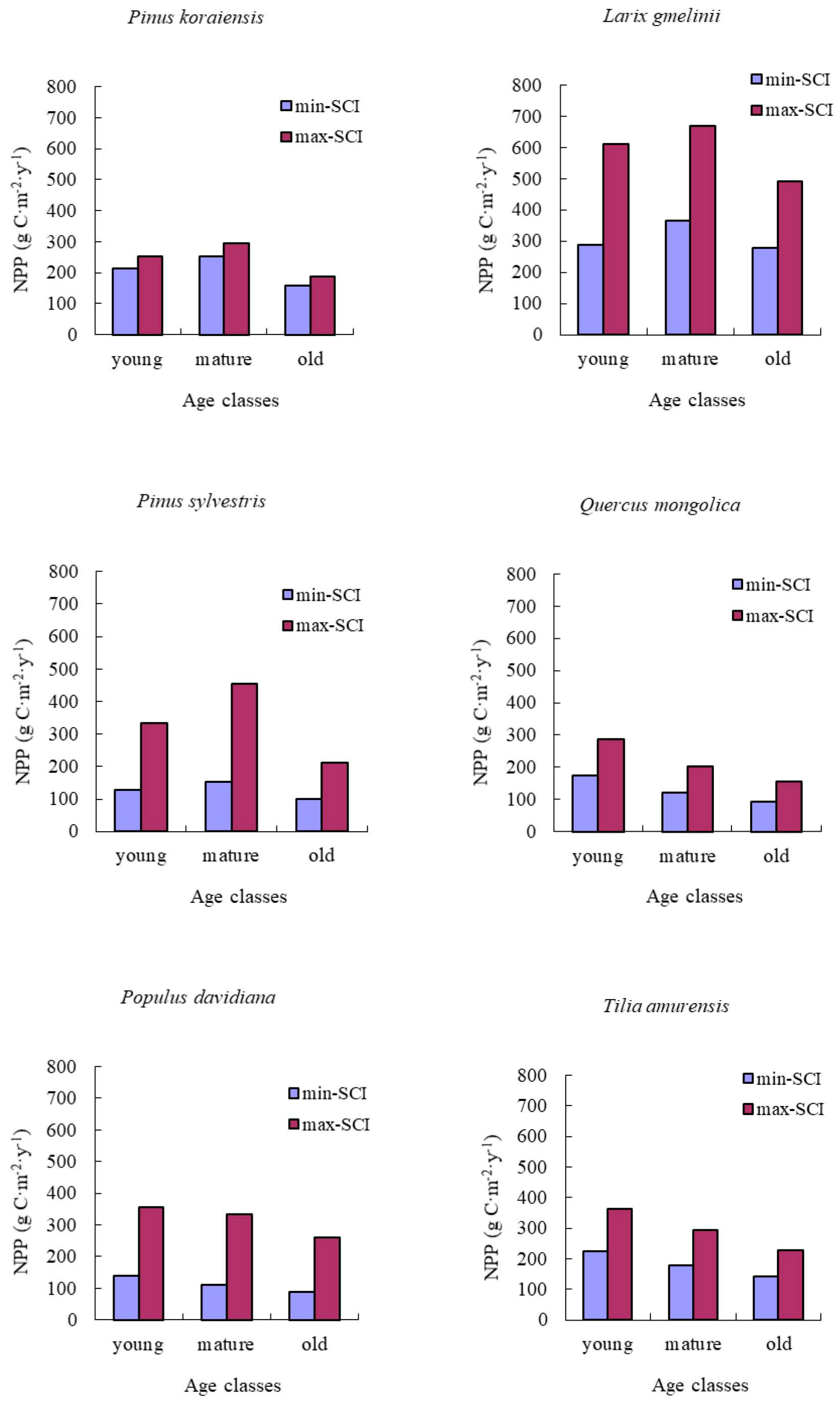
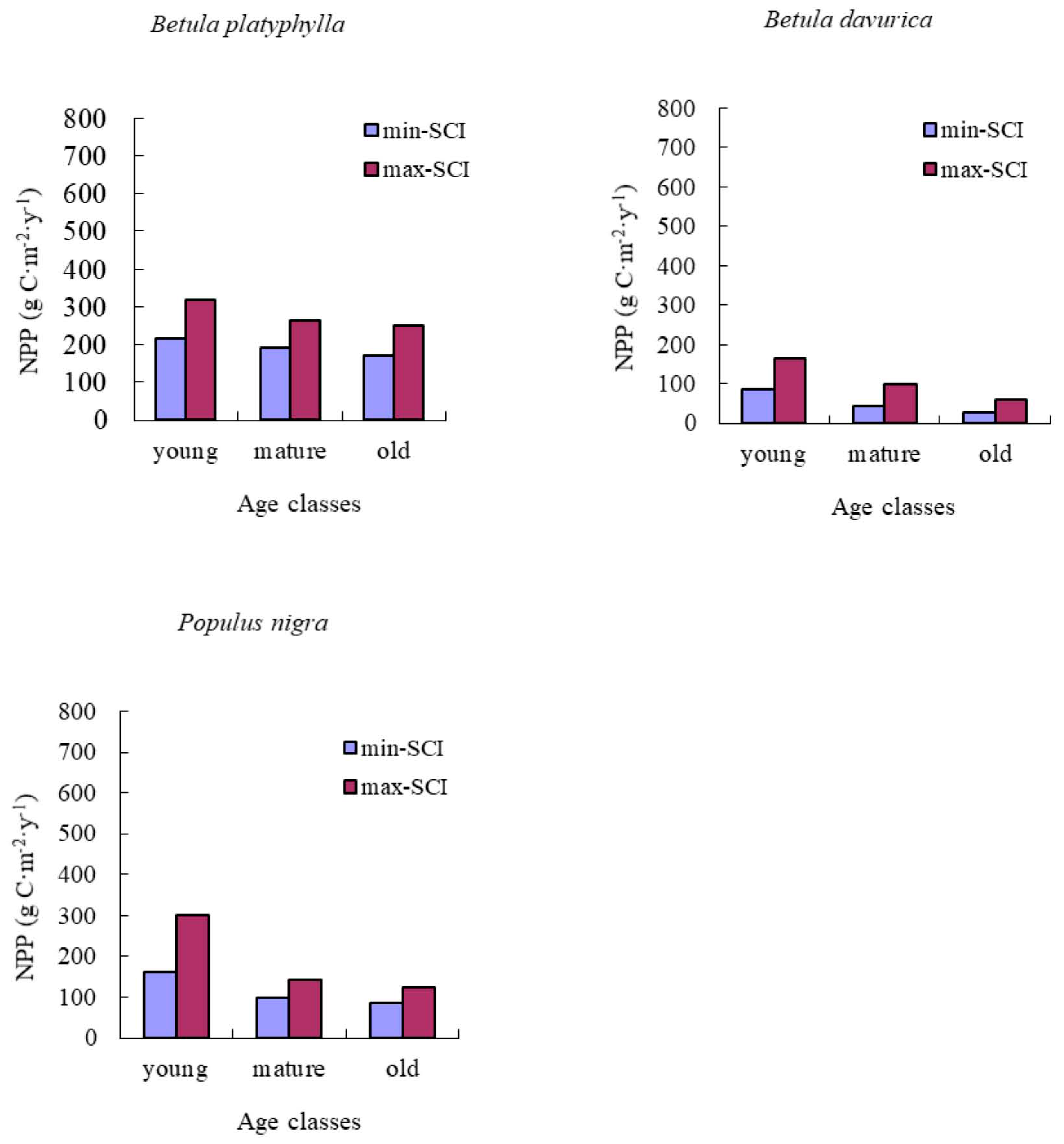
NPP-age relationships varied with site quality; higher productivity stands reached maximum NPP earlier than less productive stands. In addition, the decrease in NPP with age was also more dramatic for more productive sites. Our results were similar to those reported by Chen et al. [
17].
We compared our NPP-age relationships with similar studies for China and the U.S. [
24,
26]. Our NPP-age curves were similar to the curves derived for China and U.S. forests except for DBF. Wang et al. [
24] reported that the age at which the NPP reached peak value was always older, especially for DBF, which did not show a decline until the age of 120. However, our NPP-age relationships for DBF showed the decline of NPP at approximately 20 years of stand age. Three possible reasons included: first, there were only 37 samples from Heilongjiang Province in Wang’s study; second, Heilongjiang Province has fertile soil and earlier forest maturity than other regions; and lastly, the NPP-age relationships derived from BEPS modeling used in Wang et al. [
24] may cause more error than that from yield tables.
Two studies had found old forests to be as productive as young forest stands [
15,
16]. In our study, the predicted NPP of old forests declined by an average value of 33% from its peak for all of forest types and groups studied. This may be caused by many other site factors, including climate, soil, and drainage. These factors caused the decline of biomass increase in old stands.
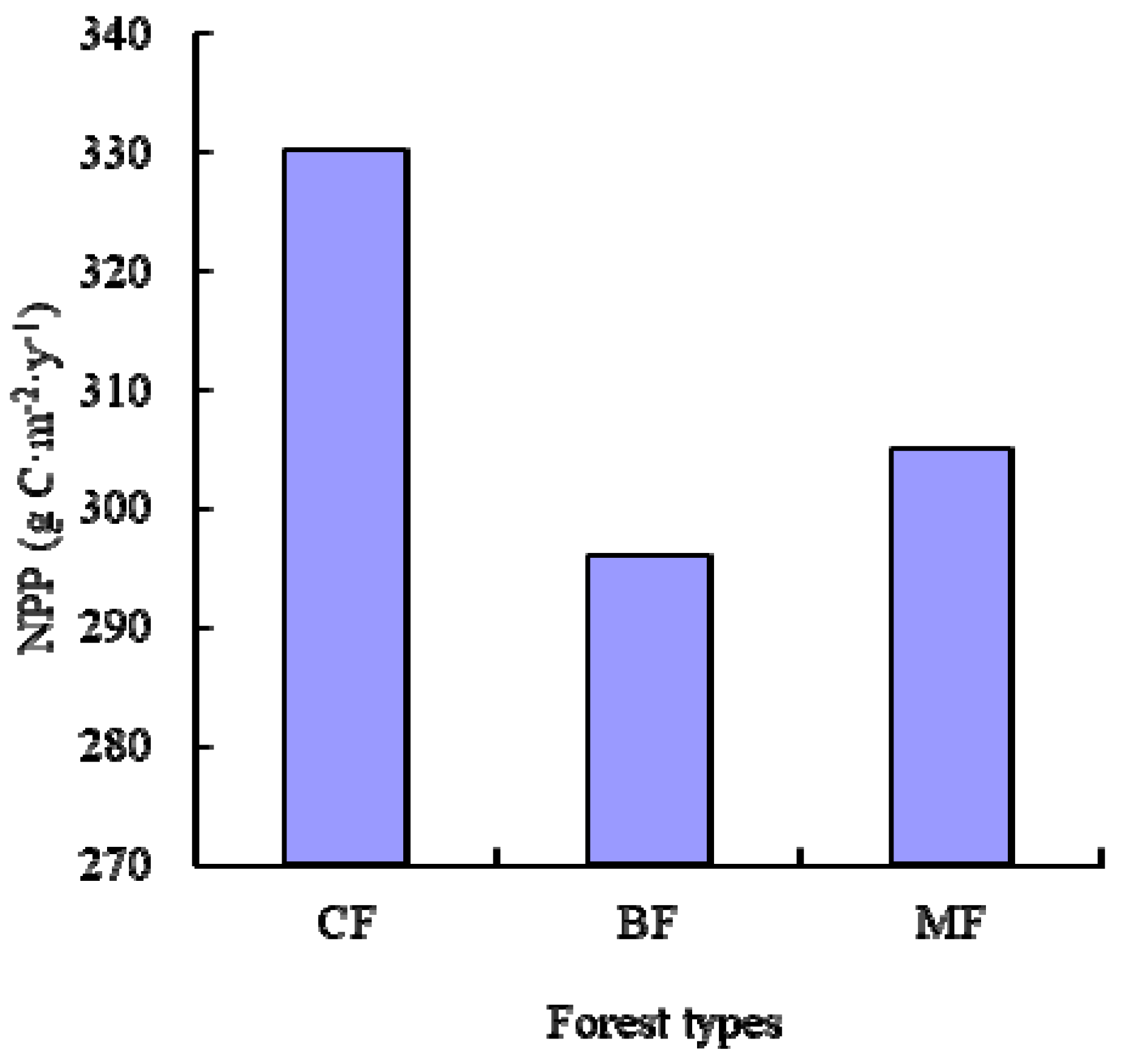
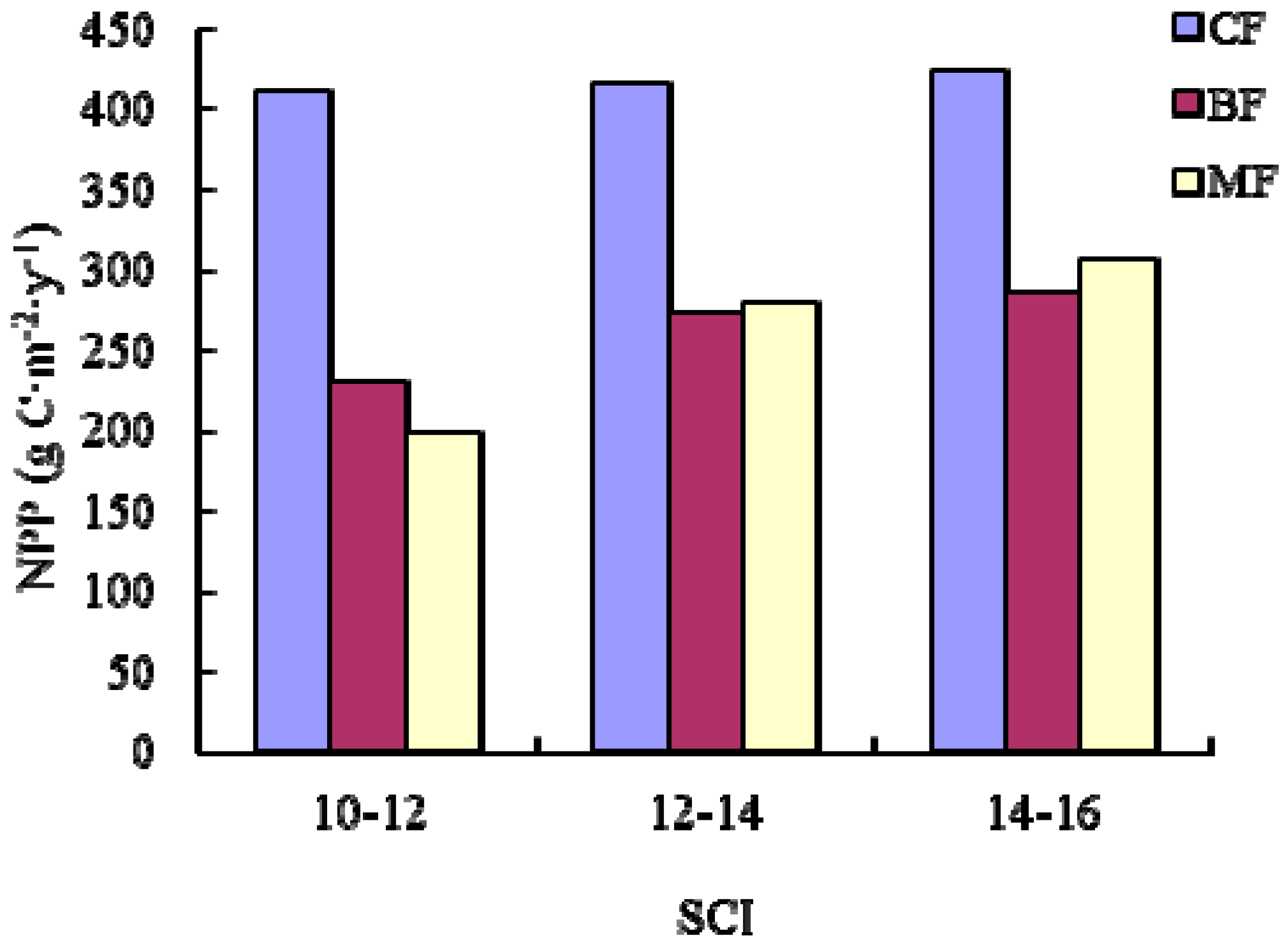
We implemented these NPP-age relationships into the InTEC model to estimate the NPP in Heilongjiang from 2000 to 2010. Our results showed that the average NPP of coniferous forest type groups was higher than that for the other two forest type groups. This aligned with the research of Mao et al. [
53], which simulated NPP based on MODIS and AVHRR NDVI data sets. Our results also indicated the younger stands and higher SCI values may lead to a higher NPP, which agrees with the findings of Chen et al. [
17]. However, the average NPP of the three main forest type groups was slightly lower than found in the research of Mao et al. [
53]. MODIS NPP had been proven to oversimulate because the product was strongly driven by climate data [
54].
6. Conclusions
In this study, we developed a specific method for estimating the relationships between NPP and age, at different SCI values for 12 forest types and groups, based on data from yield tables, biomass equations, and forest inventories. We derived the stand-level NPP through quantifying the four components: total biomass annual increment, tree mortality, turnover of foliage and fine roots, and the NPP of understory vegetation and moss.
Two similarities in the temporal patterns of the NPP-age relationships at various SCI values occurred between various forest types and groups: first, NPP increased rapidly during early stages of stand growth, reached a peak when young or mature, and then declined after maturity; secondly, high productivity sites had higher maximum NPP than poor productivity sites.
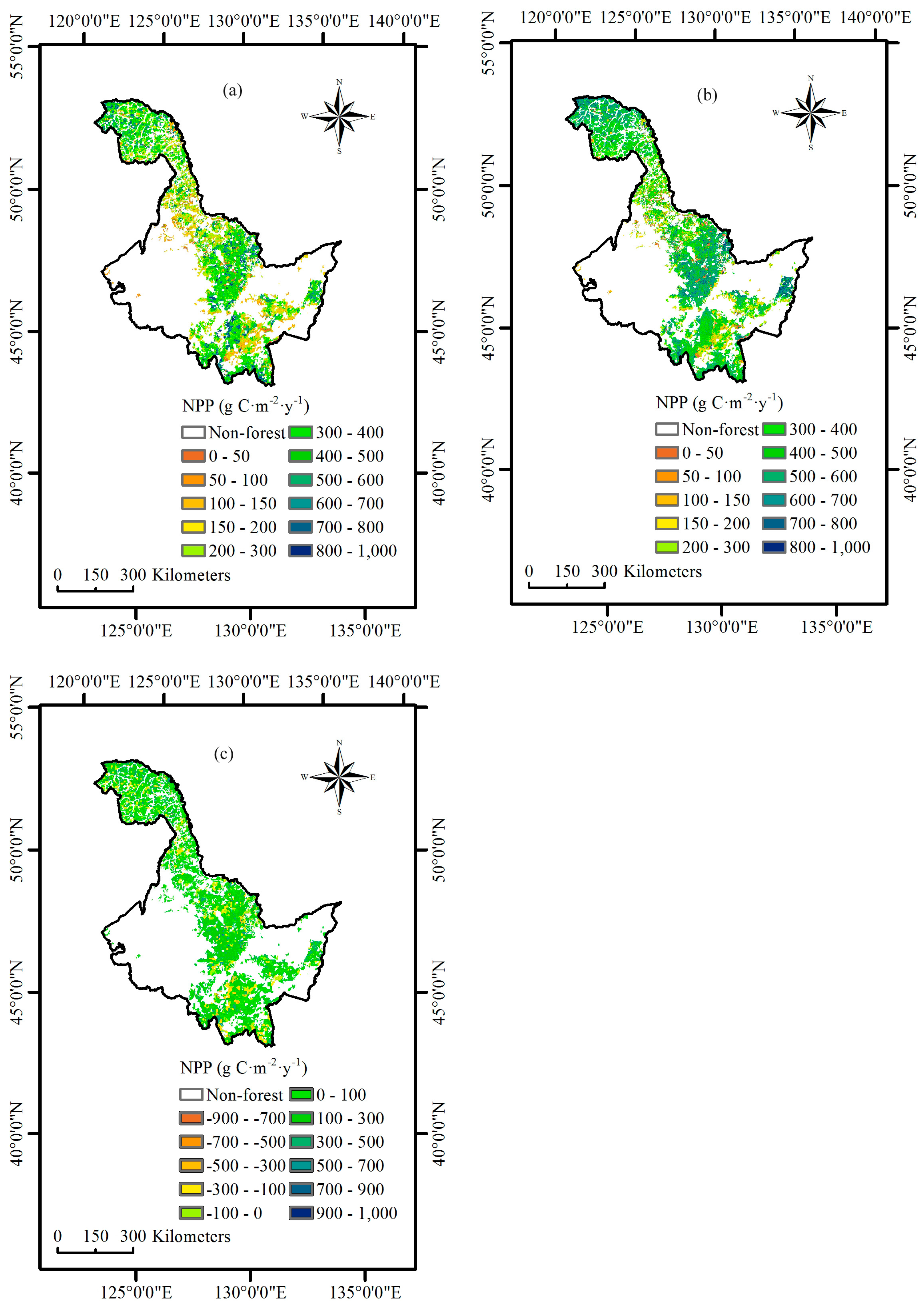
We implemented these NPP-age relationships at various SCI values into the InTEC model to estimate the NPP for three major forest biomes: coniferous forest, broadleaved forest, and mixed forest in Heilongjiang Province, China, from 2001 to 2010. During this period, the average NPP of coniferous forest was larger than broadleaved forest and mixed forest due to a greater carbon fixation capability and being productive at a mature age.
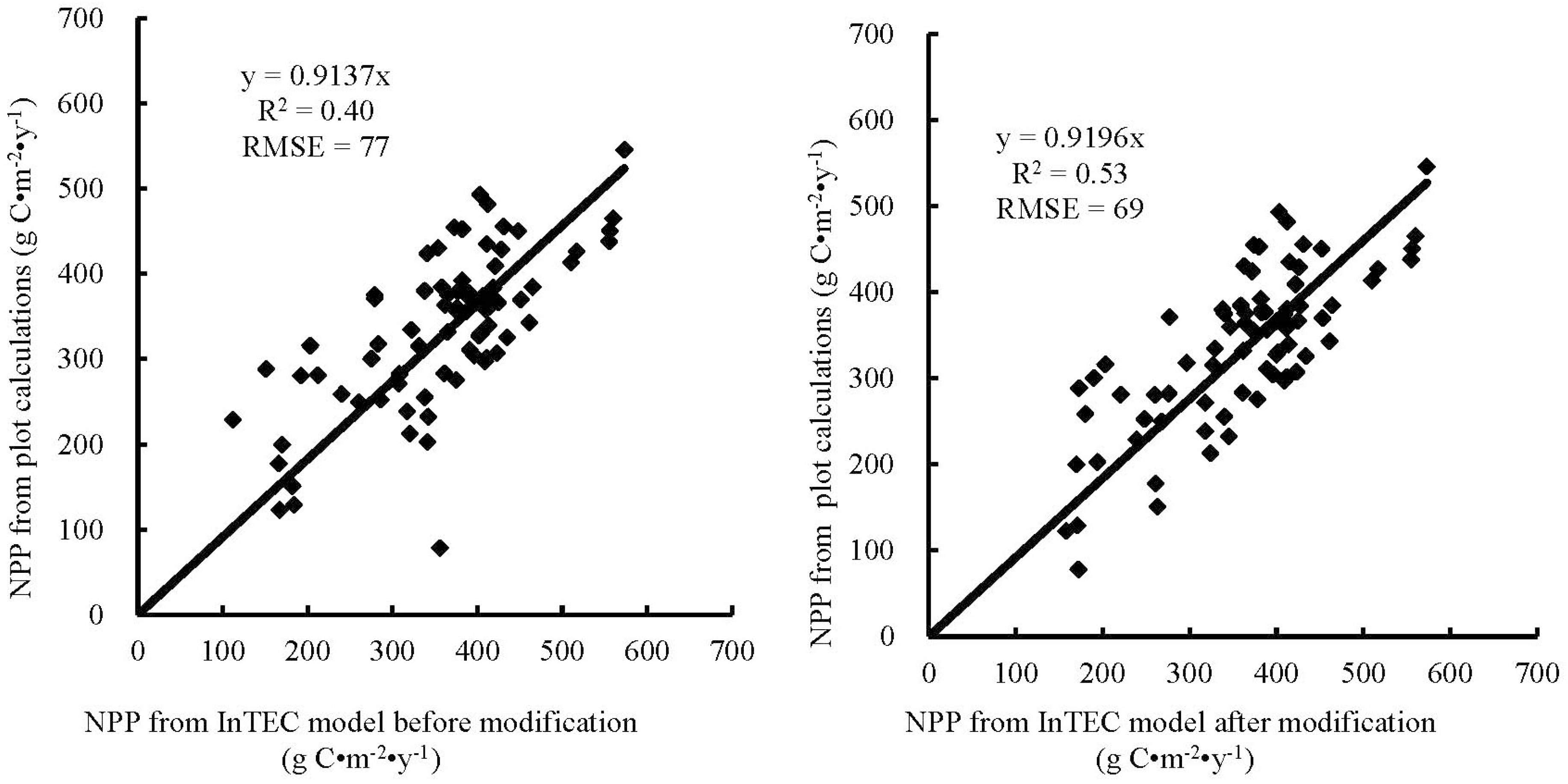
In this study, we also compared the results from InTEC modeling after modification with the new NPP-age relationships at various SCI values derived from this study. The model results showed that the new NPP-age relationships could better fit the NPP changes with age and SCI variation than the old ones used in the model.
However, there were still some weaknesses in our study: First, because mortality information was not included in the yield tables, we used the average mortality rate of biomass as the first order approximation; and second, the parameters of foliage and fine root turnover rates were obtained from White et al. [
49] for North American forests, because of the lack of local measurements for these parameters. Our future studies could be directed toward acquiring this information in our region.
Although large uncertainties still exist in the development of these NPP-age relationships, this was nevertheless the first study in China to develop localized NPP-age relationships at various SCI values. These relationships have many potential uses for the analysis of forest management because they provide a new, independent, and comprehensive source of information on forest growth. This study should have implications for other forest regions of the world where SCI varies considerably.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










