Is Resistance to Mountain Pine Beetle Associated with Genetic Resistance to White Pine Blister Rust in Limber Pine?


Abstract
:1. Introduction
2. Materials and Methods
2.1. Resin Duct Measurements
2.2. Data Analysis
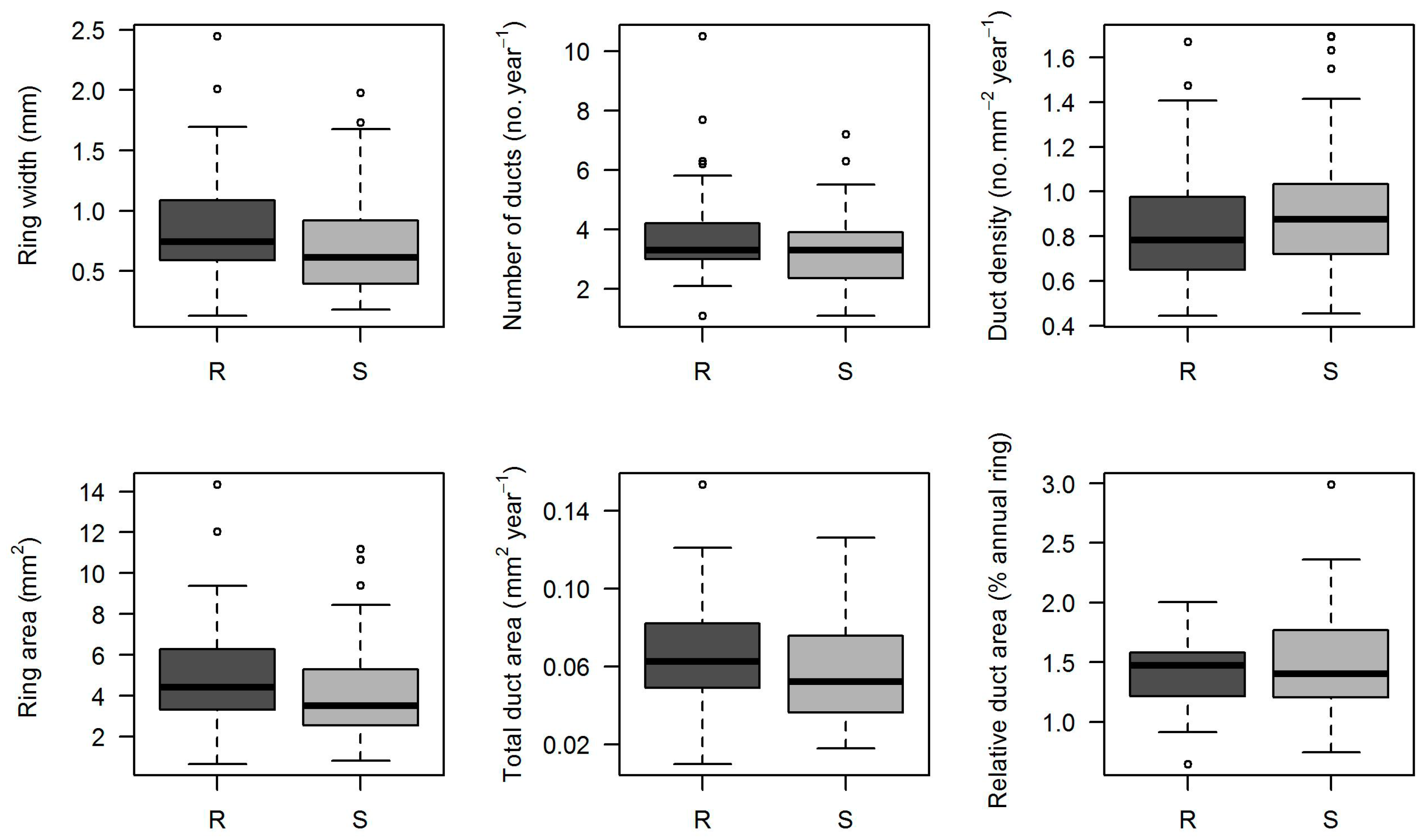
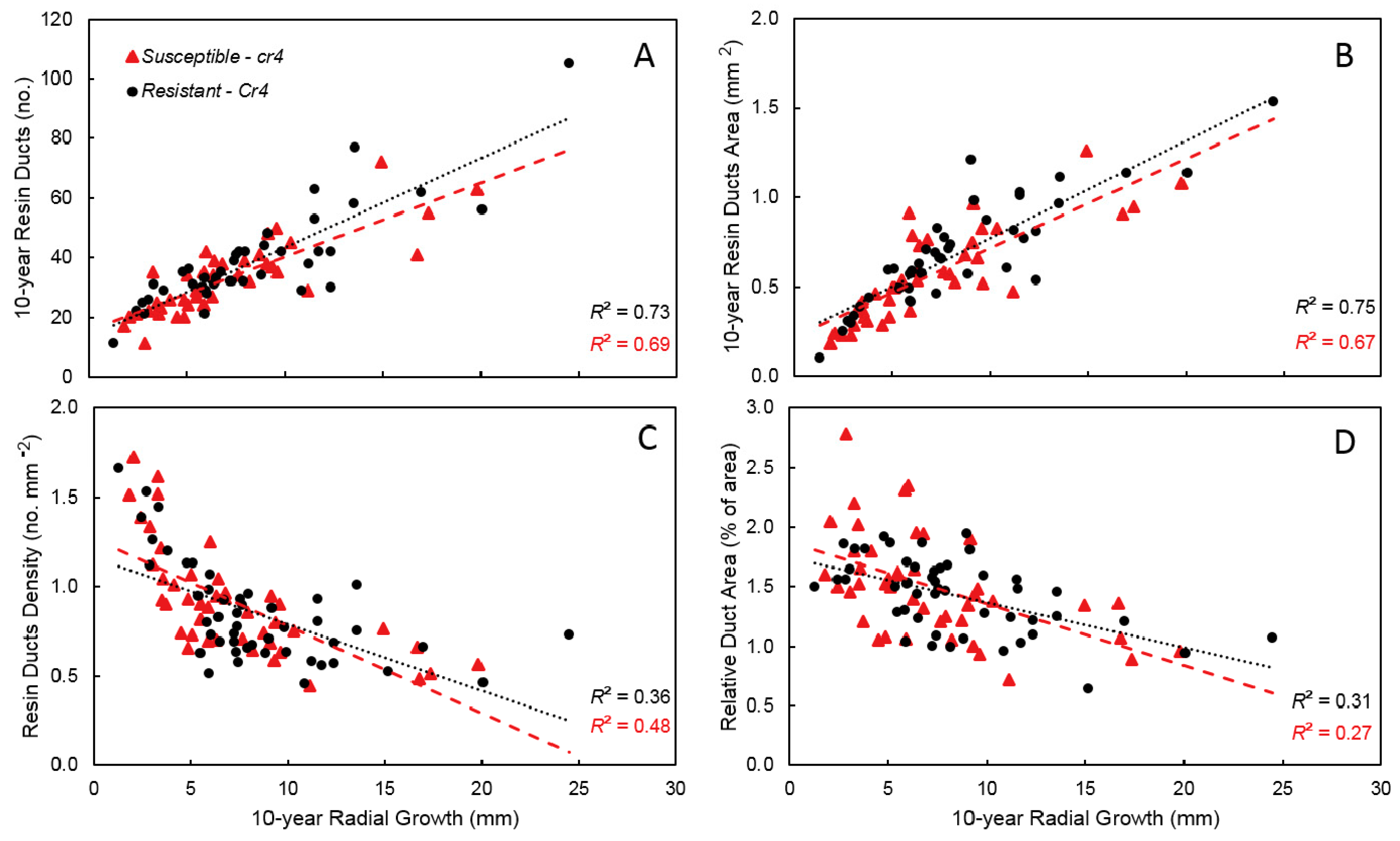
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Krist, F.J.; Ellenwood, J.R.; Woods, M.E.; McMahan, A.J.; Cowardin, J.P.; Ryerson, D.E.; Sapio, F.J.; Zweifler, M.O.; Romero, S.A. 2013–2027 National Insect and Disease Forest Risk Assessment; United States Forest Service: Missoula, MT, USA, 2014; pp. 1–199. [Google Scholar]
- Wood, D.L. The role of pheromones kairomones, and allomones in the host selection and colonization behavior of bark beetles. Annu. Rev. Entomol. 1982, 27, 411–446. [Google Scholar] [CrossRef]
- Six, D.L.; Bracewell, R. Dendroctonus ponderosae Hopkins (mountain pine beetle). In Bark Beetles: Biology and Ecology of Native and Invasive Insect; Fernando, E.V., Richard, W.H., Eds.; Elsevier: London, UK, 2015; pp. 305–350. [Google Scholar]
- Hicke, J.A.; Logan, J.A.; Powell, J.; Ojima, D.S. Changing temperatures influence suitability for modeled mountain pine beetle (Dendroctonus ponderosae) outbreaks in the western United States. J. Geophys. Res. 2006, 111, 1–12. [Google Scholar] [CrossRef]
- Bentz, B.; Campbell, E.; Gibson, K.; Kegley, S.; Logan, J.; Six, D. Mountain pine beetle in high-elevation five-needle white pine ecosystems. In The Future of High-Elevation, Five-Needle White Pines in Western North America, Proceedings of the High Five Symposium, Missoula, MT, USA, 28–30 June 2010; Keane, R.E., Tomback, D.F., Murray, M.P., Smith, C.M., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2011; pp. 78–84. [Google Scholar]
- Weed, A.S.; Bentz, B.J.; Ayres, M.P.; Holmes, T.P. Geographically variable response of Dendroctonus ponderosae to winter warming in the western United States. Landsc. Ecol. 2015, 30, 1075–1093. [Google Scholar] [CrossRef]
- Gibson, K.; Slov, K.; Kegley, S.; Jorgensen, C.; Smith, R.; Witcosky, J. Mountain Pine Beetle Impacts in High-Elevation Five-Needle Pines: Current Trends and Challenges; USDA Forest Service, Forest Health Protection, Northern Region: Missoula, MT, USA, 2008; pp. 1–32.
- Logan, J.A.; Macfarlane, W.W.; Willcox, L. Whitebark pine vulnerability to climate-driven mountain pine beetle disturbance in the Greater Yellowstone Ecosystem. Ecol. Appl. 2010, 20, 895–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fettig, C.J.; Gibson, K.E.; Munson, A.S.; Negron, J.F. Cultural practices for prevention and mitigation of mountain pine beetle infestations. For. Sci. 2014, 60, 450–463. [Google Scholar] [CrossRef]
- Buotte, P.C.; Hicke, J.A.; Preisler, H.K.; Abatzoglou, J.T.; Raffa, K.F.; Logan, J.A. Climate influences on whitebark pine mortality from mountain pine beetle in the Greater Yellowstone Ecosystem. Ecol. Appl. 2016, 26, 2507–2524. [Google Scholar] [CrossRef] [PubMed]
- Van de Gevel, S.L.; Larson, E.R.; Grissino-Mayer, H.D. Separating trends in whitebark pine radial growth related to climate and mountain pine beetle outbreaks in the northern Rocky Mountains, USA. Forests 2017, 8, 195. [Google Scholar] [CrossRef]
- Shepherd, B.; Jones, B.; Sissons, R.; Cochrane, J.; Park, J.; Smith, C.M.; Stafl, N. Ten years of monitoring illustrates a cascade of effects of white pine blister rust and focuses whitebark pine restoration in the Canadian Rocky and Columbia Mountains. Forests 2018, 9, 138. [Google Scholar] [CrossRef]
- Langor, D.W. Host effect on the phenology, development, and mortality of field populations of the mountain pine beetle. Can. Entomol. 1989, 121, 149–157. [Google Scholar] [CrossRef]
- Amman, G.D.; Pace, V.E. Optimum Egg Gallery Densities for The Mountain Pine Beetle in Relation to Lodgepole Pine Phloem Thickness; USDA Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1976; pp. 1–8, Research Note INT–209.
- Langor, D.W.; Spence, J.R.; Pohl, G.R. Host effects on fertility and reproductive success of Dendroctonus ponderosae Hopkins (Coleoptera:Scolytidae). Evolution 1990, 44, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Cerezke, H.F. Egg gallery, brood production, and adult characteristics of mountain pine beetle, Dendroctonus ponderosae Hopkins (Coleoptera:Scolytidae), in three pine hosts. Can. Entomol. 1995, 127, 955–965. [Google Scholar] [CrossRef]
- Negron, J.F.; Fettig, C.J. Mountain pine beetle, a major disturbance agent in U.S. western coniferous forests: A synthesis of the state knowledge. For. Sci. 2014, 60, 409–413. [Google Scholar] [CrossRef]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. Bioscience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- O’Neill, G.A.; Aitken, J.N.; Alfaro, R.I. Geographic variation in resin canal defenses in seedlings from the Sitka spruce x white spruce introgression zone. Can. J. For. Res. 2002, 32, 390–400. [Google Scholar] [CrossRef]
- Kane, J.M.; Kolb, T.E. Importance of resin ducts in reducing ponderosa pine mortality from bark beetle attack. Oecologia 2010, 164, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Ferrenberg, S.; Kane, J.M.; Mitton, J.B. Resin duct characteristics associated with tree resistance to bark beetles across lodgepole and limber pines. Oecologia 2014, 174, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Seybold, S.J.; Huber, D.P.W.; Lee, J.C.; Graves, A.D.; Bohlmann, J. Pine monoterpenes and pine bark beetles: A marriage of convenience for defense and chemical communication. Phytochem. Rev. 2006, 5, 143–178. [Google Scholar] [CrossRef]
- Reid, R.W.; Whitney, H.S.; Watson, J.A. Reactions of lodgepole pine to attack by Dendroctonus ponderosae Hopkins and blue stain fungi. Can. J. Bot. 1967, 45, 1115–1126. [Google Scholar] [CrossRef]
- Hood, S.; Sala, A. Ponderosa pine resin defenses and growth: Metrics matter. Tree Physiol. 2015, 35, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- King, J.N.; Noshad, D.D.; Smith, J.A. review of genetic approaches to the management of blister rust in white pines. For. Pathol. 2010, 40, 292–313. [Google Scholar] [CrossRef]
- Looney, C.E.; Waring, K.M.; Fairweather, M.L. Monitoring the health of Pinus strobiformis: Early impacts of white pine blister rust invasion. In Forest Health Monitoring: National Status, Trends, and Analysis 2013; Kevin, M.P., Barbara, L.C., Eds.; U.S. Department of Agriculture Forest Service: Missoula, MT, USA, 2015; pp. 167–174, General Technical Report SRS-207. [Google Scholar]
- Johnson, D.W.; Jacobi, W.R. First report of white pine blister rust in Colorado. Plant Dis. 2000, 84, 595. [Google Scholar] [CrossRef]
- Blodgett, J.T.; Sullivan, K.F. First Report of white pine blister rust on rocky mountain bristlecone pine. Plant Dis. 2004, 88, 311. [Google Scholar] [CrossRef]
- Kearns, H.S.; Jacobi, W.R. The distribution and incidence of white pine blister rust in Central and Southeastern Wyoming and Northern Colorado. Can. J. For. Res. 2007, 37, 462–472. [Google Scholar] [CrossRef]
- Government of Alberta. COSEWIC Assessment and Status Report on the Limber Pine Pinus flexilis in Canada; Species at Risk Public Registry: Ottawa, ON, Canada, 2014.
- Burns, K.S.; Schoettle, A.W.; Jacobi, W.R.; Mahalovich, M.F. Options for the Management of White Pine Blister Rust in The Rocky Mountain Region; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2008; pp. 1–26, RMRS-GTR-206.
- Schoettle, A.W.; Burns, K.S.; Cleaver, C.M.; Connor, J.J. Proactive Limber Pine Conservation Strategy for the Greater Rocky Mountain National Park Area; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2018; pp. 1–81, RMRS-GTR-379.
- Tomback, D.F.; Achuff, P. Blister rust and western forest biodiversity: Ecology, values and outlook for white pines. For. Pathol. 2010, 40, 186–225. [Google Scholar] [CrossRef]
- Smith, C.M.; Langor, D.W.; Myrholm, C.; Weber, J.; Gillies, C.; Stuart-Smith, J. Changes in white pine blister rust infection and mortality in limber pine over time. Can. J. For. Res. 2013, 43, 919–928. [Google Scholar] [CrossRef]
- Cleaver, C.M.; Jacobi, W.R.; Burns, K.S.; Means, R.E. Limber pine in the central and southern Rocky Mountains: Stand conditions and interactions with blister rust, mistletoe, and bark beetles. For. Ecol. Manag. 2015, 358, 139–153. [Google Scholar] [CrossRef]
- Hebard, F.V.; Griffin, G.J.; Elkins, J.R. Summary of research on biology of hypovirulent and virulent Endothia parasitica on blight-resistant and blight-susceptible Chestnut trees at Virginia Polytechnic Institute and State University. In American Chestnut Proceedings; USDA Forest Service American Chestnut Cooperators’ Meeting: Morgantown, WV, USA, 1982. [Google Scholar]
- Griffin, G.J.; Hebard, F.V.; Wendt, R.W.; Elkins, J.R. Survival of American chestnut trees: Evaluation of blight resistance and virulence in Endothia parasitica. Phytopathology 1983, 73, 1084–1092. [Google Scholar] [CrossRef]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliot, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Schoettle, A.W.; Sniezko, R.A.; Kegley, A.; Burns, K.S. White pine blister rust resistance in limber pine: Evidence for a major gene. Phytopathology 2014, 104, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Kinloch, B.B., Jr.; Dupper, G.E. Genetic specificity in the white pine-blister rust pathosystem. Phytopath. 2002, 92, 278–280. [Google Scholar] [CrossRef] [PubMed]
- Kinloch, B.B., Jr; Littlefield, J.L. White pine blister rust: Hypersensitive resistance in sugar pine. Can. J. Bot. 1977, 55, 1148–1155. [Google Scholar] [CrossRef]
- Kinloch, B.B., Jr.; Parks, G.K.; Fowler, C.W. White pine blister rust: Simply inherited resistance in sugar pine. Science 1970, 167, 193–195. [Google Scholar] [CrossRef] [PubMed]
- Kinloch, B.B., Jr.; Sniezko, R.A.; Barnes, G.D.; Greathouse, T.E. A major gene for resistance to white pine blister rust in western white pine from the Cascade Range. Phytopathology 1999, 89, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-J.; Schoettle, A.W.; Sniezko, R.A.; Sturrock, R.N.; Zamany, A.; Williams, H.; Ha, A.; Chan, D.; Danchok, B.; Savin, D.P.; et al. Genetic mapping of Pinus flexilis major gene (Cr4) for resistance to white pine blister rust using transcriptome-based SNP genotyping. BMC Genomics 2016, 17, 753. [Google Scholar] [CrossRef] [PubMed]
- Kinloch, B.B., Jr.; Sniezko, R.A.; Dupper, G.E. Origin and distribution of Cr2, a gene for resistance to white pine blister rust in natural populations of western white pine. Phytopathology 2003, 93, 691–694. [Google Scholar] [CrossRef] [PubMed]
- Kinloch, B.B., Jr. Distribution and frequency of a gene for resistance to white pine blister rust in natural populations of sugar pine. Can. J. Bot. 1992, 70, 1319–1323. [Google Scholar] [CrossRef]
- Vogan, P.J.; Schoettle, A.W. Selection of resistance to white pine blister rust affects the abiotic stress tolerances of limber pine. For. Ecol. Manag. 2015, 344, 110–119. [Google Scholar] [CrossRef]
- Vogan, P.J.; Schoettle, A.W. Carbon costs of constitutive and expressed resistance to a non-native pathogen in limber pine. PLoS ONE 2016, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bockino, N.K.; Tinker, D.B. Interactions of white pine blister rust and mountain pine beetle in whitebark pine ecosystems in the southern Greater Yellowstone Area. Nat. Area J. 2012, 32, 31–40. [Google Scholar] [CrossRef]
- Six, D.L.; Adams, J. White pine blister rust severity and selection of individual whitebark pine by the mountain pine beetle (Coleoptera: Curculionidae, Scolytinae). J. Entomol. Sci. 2007, 42, 345–353. [Google Scholar] [CrossRef]
- Boone, C.K.; Aukema, B.H.; Bohlmann, J.; Carroll, A.L.; Raffa, K.F. Efficacy of tree defense physiology varies with bark beetle population density: A basis for positive feedback in eruptive species. Can. J. For. Res. 2011, 41, 1174–1188. [Google Scholar] [CrossRef]
- Dooley, E.M.; Six, D.L. Severe white pine blister rust infection in whitebark pine alters mountain pine beetle (Coleoptera: Curculionidae) attack density, emergence rate, and body size. Environ. Entomol. 2015, 44, 1384–1394. [Google Scholar] [CrossRef] [PubMed]
- Schwandt, J.; Kegley, S. Mountain Pine Beetle, Blister Rust, and Their Interaction on Whitebark Pine at Trout Lake and Fisher Peak in Northern Idaho; United States Department of Agriculture Forest Service: Missoula, MT, USA, 2004; pp. 1–6, Region 1 Report No. 04-9.
- Schoettle, A.W.; Sniezko, R.A.; Kegley, A.; Burns, K.S. Preliminary overview of the first extensive rust resistance screening tests of Pinus flexilis and Pinus aristata. In The Future of High-Elevation, Five-Needle White Pines in Western North America, Proceedings of High Five Symposium, Missoula, MT, USA, 28–30 June 2010; Keane, R.E., Tomback, D.F., Murray, M.P., Smith, C.M., Eds.; Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2011; pp. 265–269, RMRS-P-63. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Wimmer, R.; Grabner, M. Effects of climate on vertical resin duct density and radial growth of Norway spruce (Picea abies (L.) Karst.). Trees 1997, 11, 271–276. [Google Scholar] [CrossRef]
- Schoettle, A.W.; Rochelle, S.G. Morphological variation of Pinus flexilis (Pinaceae), a bird-dispersed pine, across a range of elevation. Am. J. Bot. 2000, 87, 1797–1806. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, A.; Wyka, T.P.; Zytkowiak, R.; Nihlgard, B.; Reich, P.B.; Oleksyn, J. Cold adaptation drives variability in needle structure and anatomy in Pinus sylvestris L. along a 1900 km temperate-boreal transect. Funct. Ecol. 2017, 31, 2212–2223. [Google Scholar] [CrossRef]
- Sheue, C.-R.; Yang, Y.-P.; Kuo-Huang, L.-L. Altitudinal variation of resin ducts in Pinus taiwanensis Hayata (Pinaceae) needles. Bot. Bull. Acad. Sin. 2003, 44, 305–313. [Google Scholar]
- Ferrenberg, S.; Langenhan, J.M.; Loskot, S.A.; Rozal, L.M.; Mitton, J.B. Resin monoterpene defenses decline within three widespread species of pine (Pinus) along a 1530-m elevational gradient. Ecosphere 2017, 8, 1–18. [Google Scholar] [CrossRef]
- Coley, P.D.; Bryant, J.P.; Chapin III, F.S. Resource availability and plant antiherbivore defense. Science 1985, 230, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Werker, E.; Fahn, A.F.L.S. Resin ducts of Pinus halepensis Mill.—Their structure, development and pattern of arrangement. Bot. J. Linn. Soc. 1969, 62, 379–411. [Google Scholar] [CrossRef]
- Six, D.L.; Vergobbi, C.; Cutter, M. Are survivors different? Genetic-based selection of trees by mountain pine beetle during a climate change-driven outbreak in a high-elevation pine forest. Front. Plant. Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Larson, E.R. Influences of the biophysical environment on blister rust and mountain pine beetle, and their interactions, in whitebark pine forests. J. Biogeogr. 2011, 38, 453–470. [Google Scholar] [CrossRef]
- Sniezko, R.A.; Mahalovich, M.F.; Schoettle, A.W.; Vogler, D.R. Past and Current investigations of the genetic resistance to Cronartium ribicola in high-elevation five-needle pines. In The Future of High-Elevation, Five-Needle White Pines in Western North America, Proceedings of High Five Symposium, Missoula, MT, USA, 28–30 June 2010; Keane, R.E., Tomback, D.F., Murray, M.P., Smith, C.M., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2011; pp. 246–264, RMRS-P-63. [Google Scholar]
- Keane, R.E.; Schoettle, A.W. Strategies, tools, and challenges for sustaining and restoring high elevation five-needle white pine forests in western North America. In The Future of High-Elevation, Five-Needle White Pines in Western North America, Proceedings of High Five Symposium, Missoula, MT, USA, 28–30 June 2010; Keane, R.E., Tomback, D.F., Murray, M.P., Smith, C.M., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2011; pp. 276–294, RMRS-P-63. [Google Scholar]
- Keane, R.E.; Tomback, D.F.; Aubry, C.A.; Bower, A.D.; Campbell, E.M.; Cripps, C.L.; Jenkins, M.B.; Mahalovich, M.F.; Manning, M.; McKinney, S.T.; et al. A Range-Wide Restoration Strategy for Whitebark Pine (Pinus albicaulis); U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2012; pp. 1–108, RMRS-GTR-279.
- McCaughey, W.; Scott, G.L.; Izlar, K.L. Whitebark pine planting guidelines. West. J. Appl. Forest. 2009, 24, 163–166. [Google Scholar]
- Larson, E.R.; Kipfmueller, K.F. Patterns in whitebark pine regeneration and their relationships to biophysical site characteristics in southwest Montana, central Idaho, and Oregon, USA. Can. J. For. Res. 2010, 40, 476–487. [Google Scholar] [CrossRef]
- Keane, R.E.; Parsons, R.A. Restoring whitebark pine forests of the northern Rocky Mountains, USA. Ecol. Restor. 2010, 28, 56–70. [Google Scholar] [CrossRef]
- McKinney, S.T.; Fiedler, C.E.; Tomback, D.F. Invasive pathogen threatens bird-pine mutualism: Implications for sustaining a high-elevation ecosystem. Ecol. Appl. 2009, 19, 597–607. [Google Scholar] [CrossRef] [PubMed]
- McDonald, G.I.; Hoff, R.J. Blister rust: An introduced plague. In Whitebark Pine Communities: Ecology and Restoration; Tomback, D.F., Arno, S.F., Keane, R.E., Eds.; Island Press: Washington, DC, USA, 2001; pp. 193–220. [Google Scholar]
- Schoettle, A.W.; Sniezko, R.A. Proactive intervention to sustain high-elevation pine ecosystems threatened by white pine blister rust. J. For. Res. 2007, 12, 237–336. [Google Scholar] [CrossRef]
- Schoettle, A.W.; Jacobi, W.R.; Waring, K.M.; Burns, K.S. Regeneration for resilience framework to support regeneration decisions for species with populations at risk of extirpation by white pine blister rust. New For. 2018, in press. [Google Scholar] [CrossRef]
- Tomback, D.F.; Kendall, K.C. Rocky road in the Rockies: Challenges to biodiversity. In Rocky Mountain Futures: An Ecological Perspective; Baron, J.S., Ed.; Island Press: Washington, DC, USA, 2002; pp. 153–180. [Google Scholar]
- Coop, J.D.; Schoettle, A.W. Regeneration of Rocky Mountain bristlecone pine (Pinus aristata) and limber pine (Pinus flexilis) three decades after stand-replacing fires. For. Ecol. Manag. 2009, 257, 893–903. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| State, Study Site, Land Ownership a | Site Elevation (m) | Mean Stem Diameter b (cm) | Number of Trees Sampled | |
|---|---|---|---|---|
| R c | S c | |||
| Colorado | ||||
| Brown′s Lake (Roosevelt NF) | 3230 | 26.1 | 2 | 2 |
| Cow Creek (Rocky Mountain NP) | 2763 | 17.9 | 1 | 1 |
| Crown Point (Roosevelt NF) | 3123 | 35.4 | 2 | 2 |
| Deer Mountain (Rocky Mountain NP) | 3035 | 14.6 | 1 | 1 |
| Estes Cone (Rocky Mountain NP) | 3250 | 30.5 | 3 | 2 |
| Gem Lake (Rocky Mountain NP) | 2679 | 19.2 | 4 | 4 |
| Geneva (Pike NF) | 3152 | 26.6 | 2 | 2 |
| Hunter (Rocky Mountain NP) | 3225 | 18.9 | 1 | 1 |
| Jenny Lake (Roosevelt NF) | 3325 | 24.3 | 3 | 3 |
| Lake John (Medicine Bow-Routt NF) | 2640 | 18.0 | 0 | 3 |
| Lily Lake (Rocky Mountain NP) | 2761 | 24.1 | 3 | 3 |
| MacGregor Falls (Rocky Mountain NP) | 2758 | 26.4 | 2 | 2 |
| Mosca Pass (Great Sand Dunes NP) | 2897 | 34.4 | 0 | 1 |
| Pine Ridge (Rocky Mountain NP) | 3227 | 20.7 | 2 | 2 |
| Rainbow Curve (Rocky Mountain NP) | 3312 | 20.4 | 3 | 3 |
| Sourdough (Roosevelt NF) | 2819 | 12.1 | 1 | 1 |
| Steep Mountain (Rocky Mountain NP) | 2630 | 19.0 | 2 | 2 |
| Switzerland Trail (Roosevelt NF) | 3086 | 24.6 | 3 | 1 |
| Twin Sisters (Rocky Mountain NP) | 3228 | 25.8 | 3 | 3 |
| Upper Ute (Rocky Mountain NP) | 3273 | 21.0 | 3 | 3 |
| Wyoming | ||||
| Pilot Hill (Medicine Bow NF) | 2654 | 43.6 | 5 | 2 |
| Middle Crow Creek (Medicine Bow NF) | 2376 | 28.4 | 2 | 2 |
| Variable | n | Mean (SE) | ||
|---|---|---|---|---|
| R | S | R | S | |
| Radial ring width (mm) | 49 | 44 | 0.840 (0.064) | 0.723 (0.065) |
| No. Vertical Resin Ducts (no. yr−1) | 43 | 40 | 3.847 (0.248) | 3.303 (0.200) |
| Total Duct Area (mm2 yr−1) | 43 | 40 | 0.068 (0.004) | 0.057 (0.004) |
| Duct Density (no. mm−2 yr−1) | 49 | 44 | 0.847 (0.040) | 0.925 (0.048) |
| Relative Duct Area (% annual ring) | 49 | 44 | 1.422 (0.044) | 1.509 (0.069) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holtz, C.T.; Schoettle, A.W. Is Resistance to Mountain Pine Beetle Associated with Genetic Resistance to White Pine Blister Rust in Limber Pine? Forests 2018, 9, 595. https://doi.org/10.3390/f9100595
Holtz CT, Schoettle AW. Is Resistance to Mountain Pine Beetle Associated with Genetic Resistance to White Pine Blister Rust in Limber Pine? Forests. 2018; 9(10):595. https://doi.org/10.3390/f9100595
Chicago/Turabian StyleHoltz, Christine T., and Anna W. Schoettle. 2018. "Is Resistance to Mountain Pine Beetle Associated with Genetic Resistance to White Pine Blister Rust in Limber Pine?" Forests 9, no. 10: 595. https://doi.org/10.3390/f9100595
APA StyleHoltz, C. T., & Schoettle, A. W. (2018). Is Resistance to Mountain Pine Beetle Associated with Genetic Resistance to White Pine Blister Rust in Limber Pine? Forests, 9(10), 595. https://doi.org/10.3390/f9100595