1. Introduction
In the course of recent decades, it has become a prominent goal of forest policy worldwide that future forests provide a broad spectrum of ecosystem services. Coincidentally, private forest stakeholder groups call for targeted usage of subsidies to pay forest owners for the provision of ecosystem services other than the traditional one of wood production [
1].
Mixed species stands are widely accepted to provide a broad range of benefits. They can minimize the risk of calamities [
2]. They can be more stable and more economical than monospecific stands [
3]. Thus, in many cases, they are better suited for the multifunctional provision of ecosystem services than monocultures [
4]. However, this depends on the identity of species and on the ecological context in which the mixed species stands are embedded [
5].
Pretzsch and Forrester [
6] underpin that the ecosystem service productivity of mixed stands depends on the shares of the contributing species. Mixture regulation, thus, is an obvious instrument for controlling the provision of ecosystem services. Within that scope, the tradeoff between groundwater recharge, tree species and structural diversity, and wood production is crucial. Because drought incidents will likely become more frequent in the future [
7,
8], groundwater recharge might then become a limiting process for public water supply. Diversity is a criterion of risk mitigation towards climate change as well as a forest management goal itself. Wood production provides renewable raw materials and preserves jobs. However, the specific effect of mixing proportions on these prominent ecosystem services has, to our knowledge, not been studied so far.
Due to the rapid paradigm shift towards mixed stands within Central Europe, there is a strong requirement for novel methods that control the development of mixed species forest stands [
9]. Under unmanaged conditions, mixture obviously converges towards a natural state that depends on the species-specific site suitability [
10]. According to Gadow and Füldner [
11], the crucial requirements for achieving sustainability can exclusively be met if criteria of objective control are created and subjected to a more detailed description and quantification. Given the increasing relevance of mixed species stands, the step from qualitative descriptions towards species-specific quantitative regulations is overdue [
12,
13]. Coll et al. [
14] reveal key questions about mixed forest management through a survey conducted among forest managers. They point out a knowledge gap concerning the quantitative regulation of mixed species stands.
Quantitative silvicultural guidelines are largely limited to even-aged, homogeneous monospecific stand types [
15,
16], except for very few existing approaches for density management, for example [
17,
18]. However, most existing concepts, if any, typically emphasize the qualitative steering of the species composition [
9,
19,
20]. Management planning, in contrast, requires regulation, i.e., the maintenance of quantitative target values of mixing proportions [
9]. Most current guidelines for the management of mixing proportions apply steering principles [
21,
22,
23,
24]. Abetz and Ohnemus [
18] define the number of trees as dependent on the production target, time and risk. Thus, the resulting growing space per tree does not consider the species-specific growing requirements like a typical development of the crown projection area. Rather, it considers only the requirements of the silvicultural actor and assumes a dynamic adjustment of the species’ growth requirements. A crucial aspect for mixing regulation is the ongoing availability of growing space per tree, considering species-specific typical stand and individual tree growth [
16]. Typically, thinning actions, however, do not take into account varying conditions of individual tree growth at onward time scales [
24]. Recent scientific advances increased the understanding of mixed species forests, but the extent to which this information is already suitable for consideration in practice is questionable [
14]. Pretzsch and Zenner [
9] stated that mixed species forest management guidelines should consider five aspects. When establishing mixed species stands, the temporal or spatial association of the species has to be designed: (i) a species combination with appropriate complementarity in mineral nutrients and water exploitation, as well as in space filling and light use, has to be chosen; (ii) according to mixing proportions; (iii) according to stand density; (iv) these have to be regulated during stand life; and (v) the final aspect in a quantitative management guideline for mixed species forests is the goal-oriented initiation of regeneration by volume reduction in the overstory.
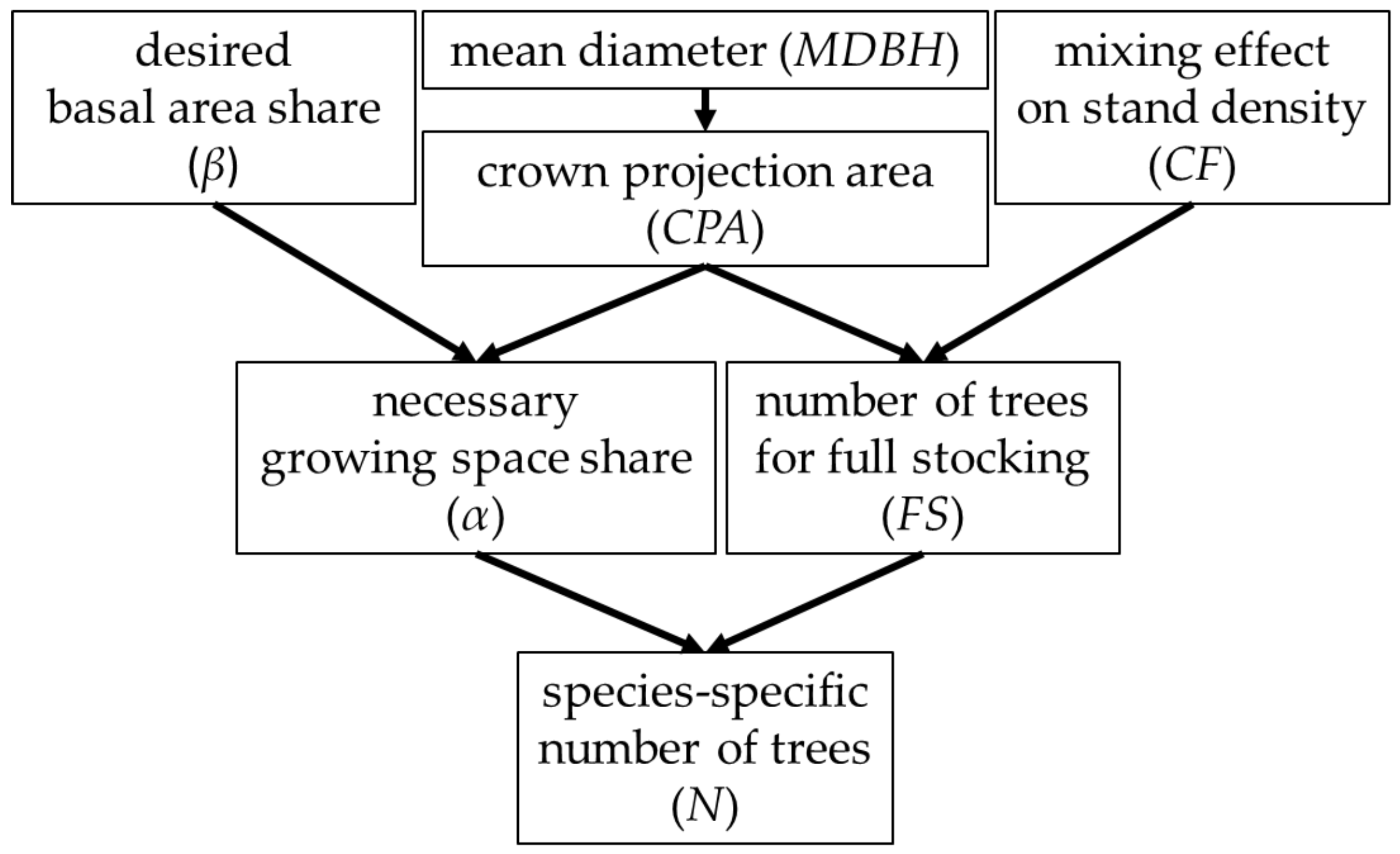
Therefore, first and foremost, in the study at hand we present an approach for the quantitative regulation of mixture proportions. In order to evaluate our approach, we implemented it in the forest management model SILVA [
25]. This enabled us to exemplarily simulate the development of mixed-species stands with different desired species shares. To this end, we chose the species Norway spruce (
Picea abies (L.) H. Karst.) and European beech (
Fagus sylvatica L.) on a fertile site which represented a very typical Central European setting so we could investigate the differential effect of the basal area shares on groundwater recharge, diversity, and wood production.
The study’s key objectives were:
To propose a quantitative, growing space-based approach for regulation of mixture proportions in mixed stands;
To demonstrate the efficiency of the approach by means of scenario simulations for a highly prominent tree species mixture in Central Europe;
To assess the effect of mixture regulation on the provision of the wood production, diversity, and groundwater recharge ecosystem services.
3. Results
3.1. Exemplary Guide Curve Calibration
3.1.1. Assumed Diameter and Top Height over Stand Age
Assuming or knowing the relation of
MDBH and
h100 to stand age is a basis for calibration of the presented regulation approach. We exemplarily assume values for beech and spruce (calculation in
Section 2.2) to calibrate the approach for the simulation study.
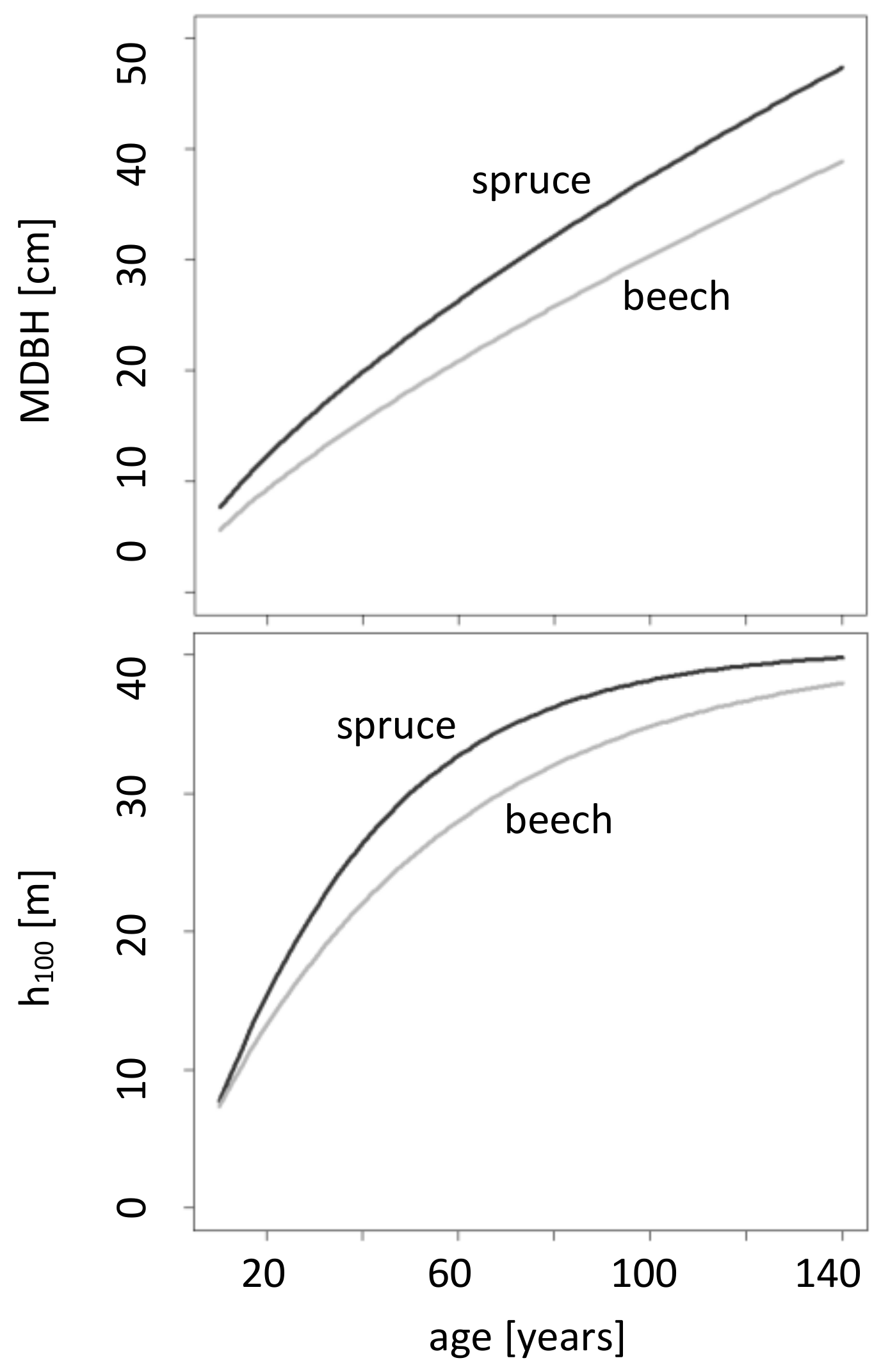
Growth of top height and diameter at breast height is higher in the case of spruce compared to beech (
Figure 2). At age 100, the prevalent top height for beech is 35 m and 38 m for spruce. Moreover, in a 100-year-old stand, a mean beech diameter at breast height of 30 cm and a mean spruce diameter at breast height of 37 cm can be assumed.
The fitted values (
Table 2 and
Table 3) describe the species-specific curves of
MDBH and
h100 in
Figure 2 and thus reveal the exact difference between the assumed growth mean diameter at breast height and top height of spruce and beech. To sum up, we can say that the assumed growth potential of spruce regarding
h100 and
MDBH is higher compared to beech.
3.1.2. Diameter Related Crown Projection Area
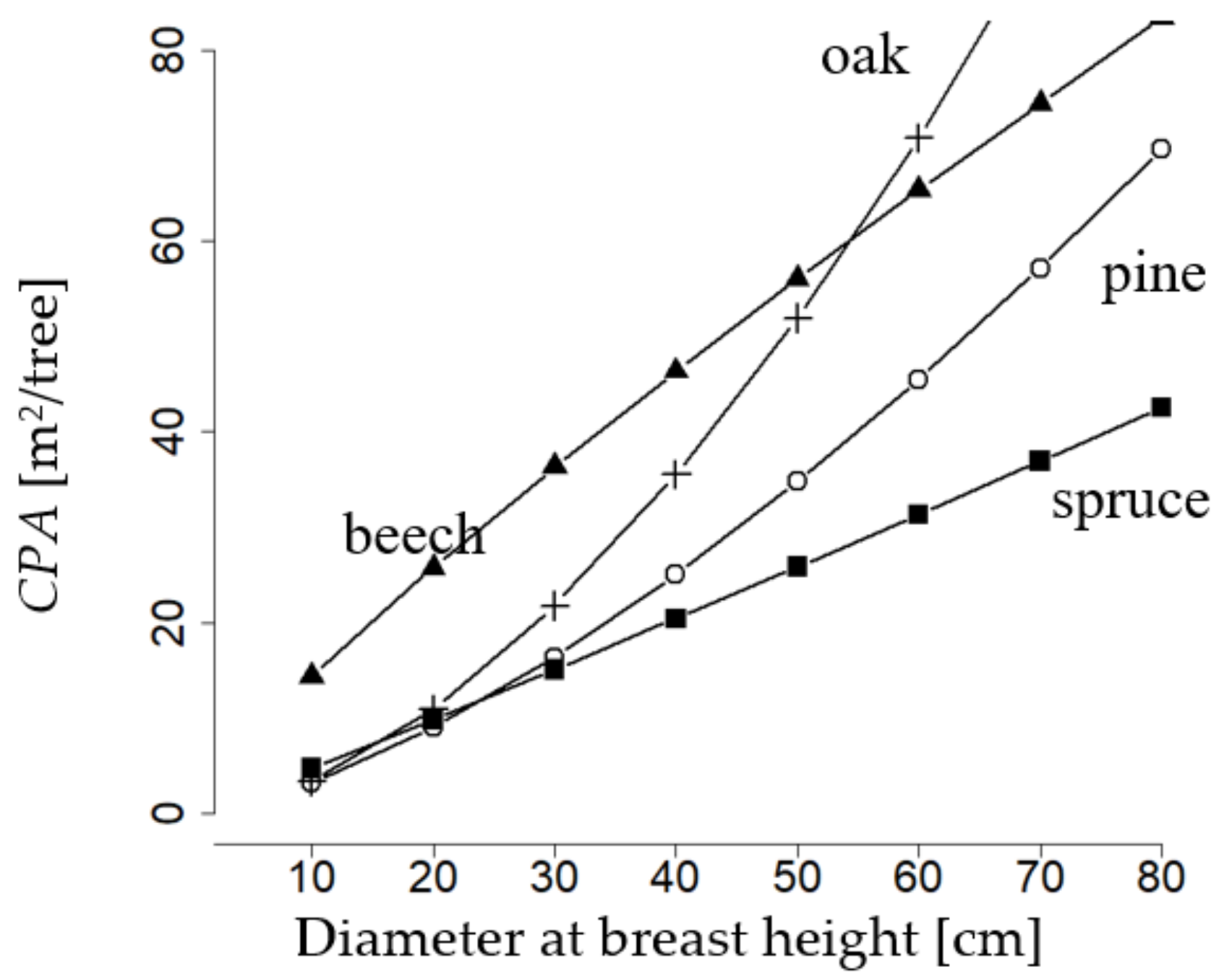
The diameter related crown projection area (
Figure 3) is a basis for the presented regulation approach. Those values are suitable for the parametrization of the mixing regulation concerning the four most important tree species in Germany. Thus, they generalize our approach beside the exemplary simulation.
Norway spruce, European beech, sessile oak, and Scots pine, notably differ in their parameters (
Table 4) obtained through fitting of the crown projection area function (
Section 2.1.1, Equation (1)). That of European beech starts with a large value of about 15 m
2 at diameter at breast height of 10 cm, while the one of the remainder species is at only 5 m
2. The crown projection area of European beech, starting from low values of diameter at breast height and throughout the whole diameter at breast height range, is markedly larger than that of Scots pine and Norway spruce. Up to a diameter at breast height of 50 cm, it also surpasses that of sessile oak. However, the slope of the crown projection area over diameter at breast height of beech decreases with diameter at breast height. Conversely, that of oak strongly increases. Thus, at a diameter at breast height of more than 50 cm, oak outruns the crown projection area of all other species. Pine, which like oak, is a light-demanding species, has a similar course of crown projection area over diameter at breast height as oak and approximates the values of beech at a diameter at breast height of 80 cm. Spruce has the lowest crown projection area over the whole range of diameter at breast height and one that constantly increases with diameter at breast height. To sum up, we can say that the species-specific relations between diameter and crown projection area are very different, even intersections are visible. Consequently, this relationship is of fundamental importance for the mixture regulation approach of this study.
3.1.3. Exemplary Guide Curve Calculation
The difference between the number of trees for full stocking in monoculture and mixture is one essential intermediate result at the calculation of the guide curve. The exemplary calibrated curves we calculated for a mixture of spruce and beech consider the following difference. Full stocking at crown closure without overlap in monoculture (
Section 2.1.2, Equation (2)) has just slightly lower values as the one under assumption of overlapping crowns resp. with mixture adjustment (
Figure 4). This is true for both tree species we focus on. However, beech stands have a stem density that is about 50% of that of spruce stands, a direct consequence of the
MDBH-
CPA relations shown in
Figure 3. That proportion continues over the whole stand age.
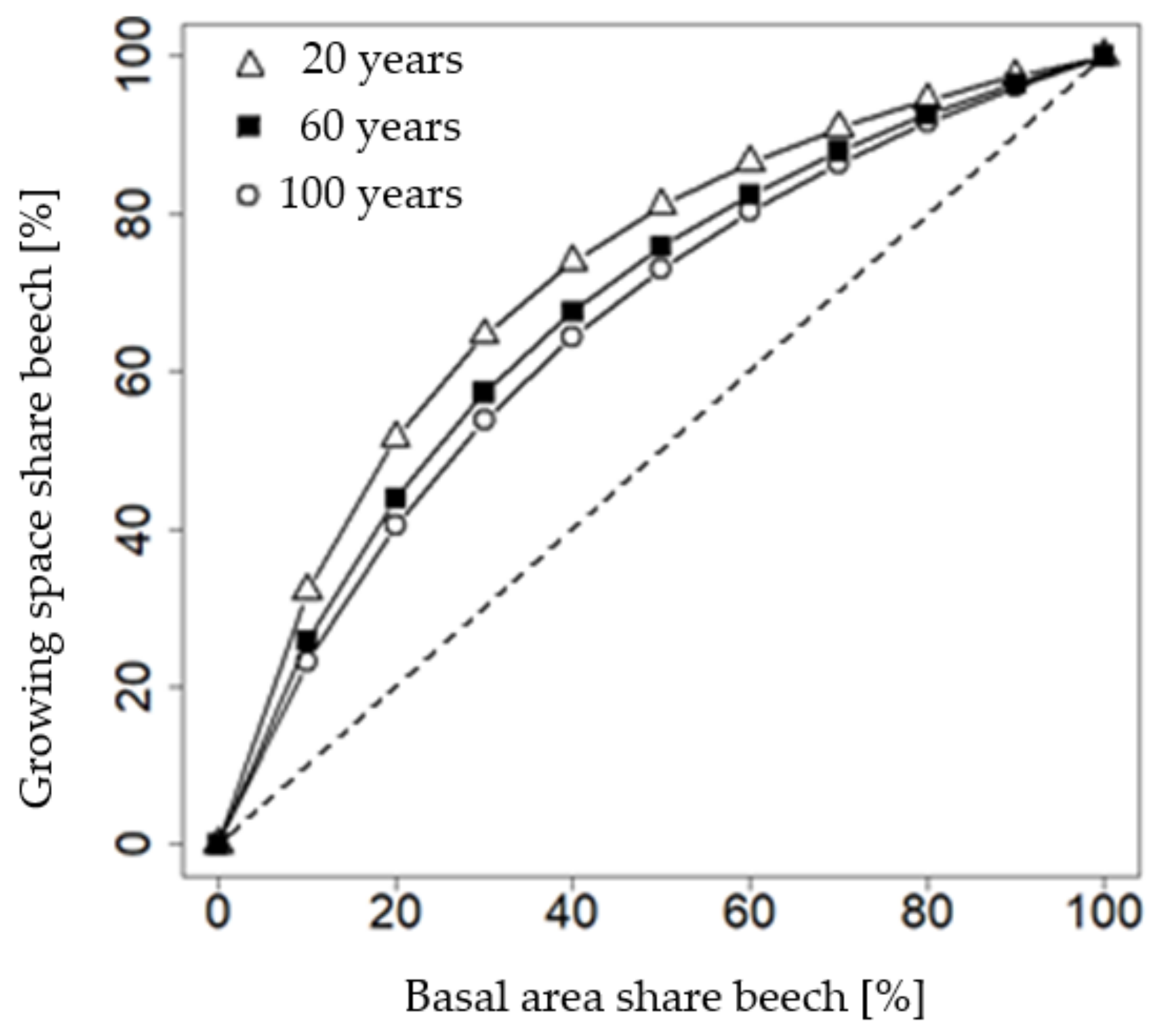
The growing space share of beech mixed with spruce is larger than its basal area share at any mixture proportion of both species being considered (
Figure 5). For example, 50% of basal area share requires a beech growing space share of 80% in a 20-year-old stand. This ratio depends on the age of the stand. In an approximately 120 year-old stand, a 50% basal area mixture requires only 70% of the growing space.
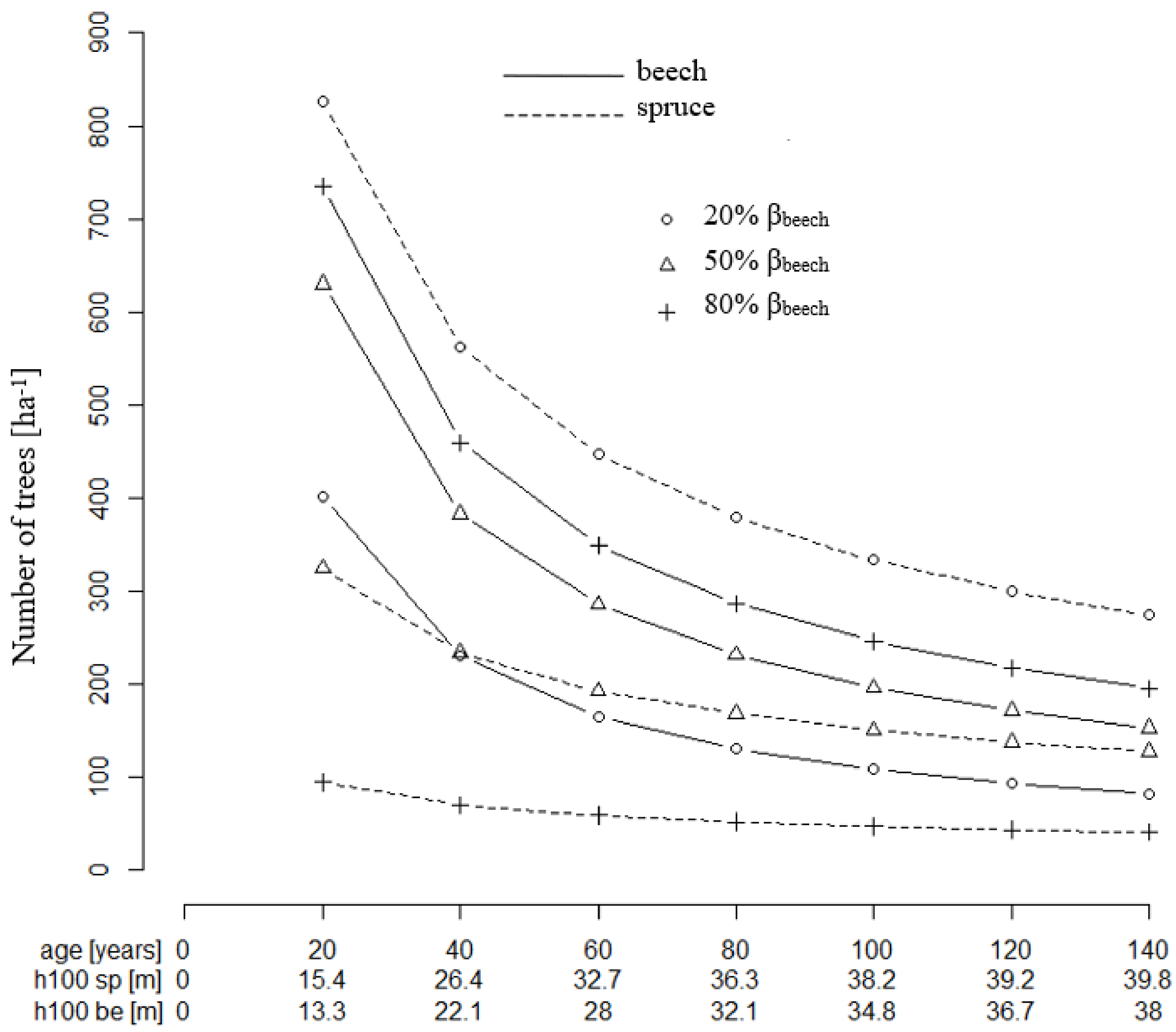
The resulting tree number guide curve (Equation (13)) for European beech and a 50% basal area mixture are marked by a strong decrease of the tree number per ha from age 20 to 140 (
Figure 6), which is the typical behavior of stem number curves in even-aged stands. While the beech growing space decreases overtime, the number of trees accordingly drops from about 630 to about 160 (beech) and 320 to about 150 (spruce), respectively. Thus, for equal basal area shares, twice as many beech trees as spruces are required in the beginning, while in an old stand with equal basal area shares, the tree numbers are about balanced. This results from the shifting relation between basal area and growing space shares, as shown above.
3.2. Simulation Study Quantifying Ecosystem Services Provision Depending on Species Shares
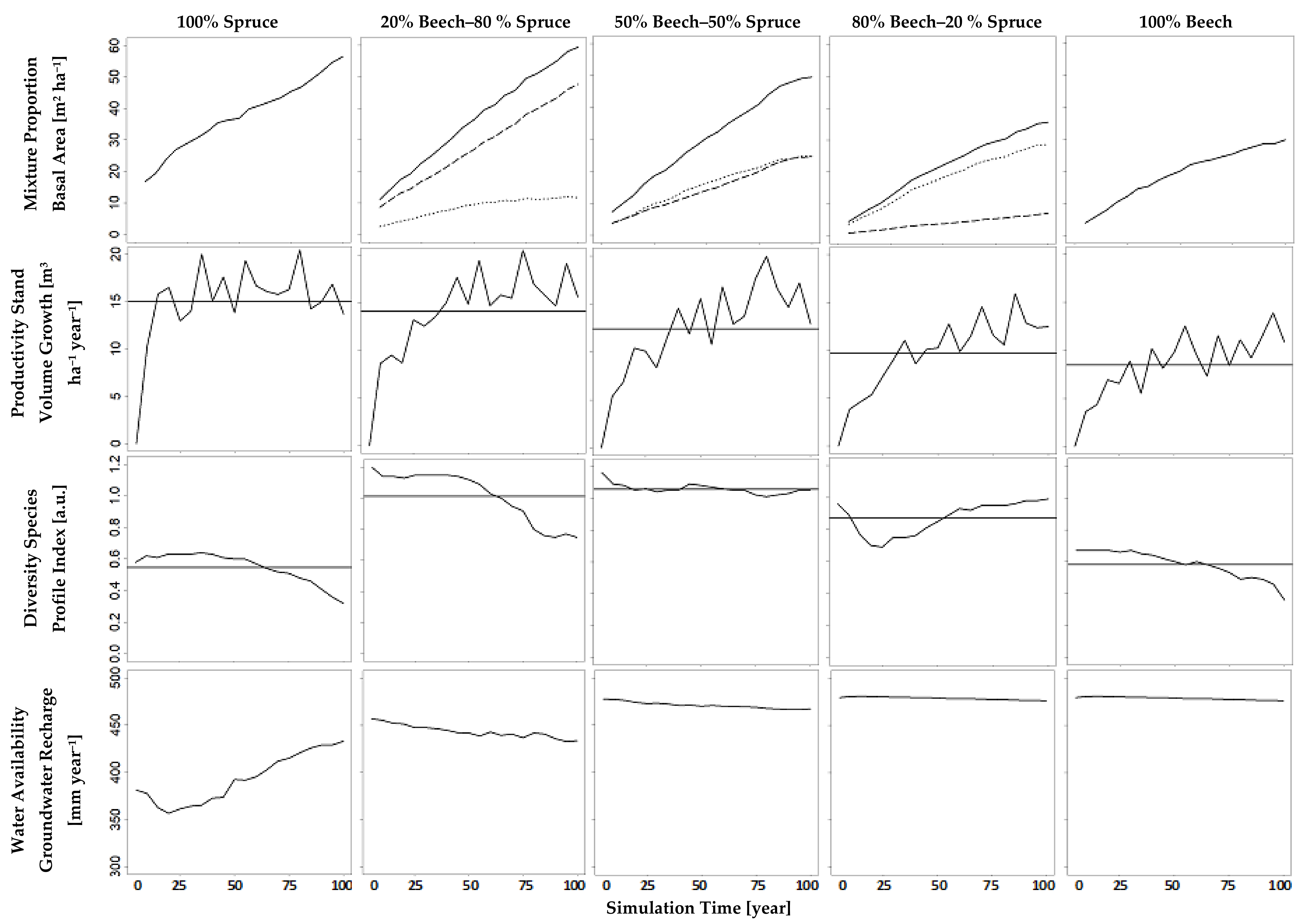
The results of our simulation runs show that the indicators for the ecosystem services we focus on, namely water availability (groundwater recharge), diversity (species profile index), and productivity (stand volume growth), notably depended on the mixture proportions (
Figure 7). The latter were defined as the desired basal area shares of beech and spruce, controlled in the simulations using the method developed above. Over the whole simulation period, the simulated beech basal area shares (
Figure 7, first row, dotted lines) had an average of 23%, 52%, and 82%, when the tree number guide curves had been adapted to a basal area share of 20%, 50% respectively 80% (
Table 5) which we deem reasonably close to the desired values. The resulting simulated growing space shares and basal area shares can be compared with the default desired basal area shares in
Table 5.
An increasing basal area share of beech reduced the stand’s total basal area (
Figure 7, first row). Accordingly, it caused a reduction of stand volume growth, i.e., productivity. Conversely, an increasing basal area share of that deciduous species markedly fostered groundwater recharge, i.e., water availability; but a beech share higher than 50% did not increase groundwater recharge anymore (
Figure 7, bottom row). The results show that groundwater recharge, within that study, is not very sensitive above a beech basal area share over 50%. This is due to the fact that even small basal area shares of beech result in large growing space shares (
Table 5). At a basal area share beech of 50%, the indicator of diversity, species profile index was highest.
The curves in
Figure 7 illustrate a trade-off between productivity and water availability with changing mixture proportions. Water availability may be aggravated in the future through the choice of a tree species that aims at increasing forest productivity; 100% spruce maximizes productivity and minimizes groundwater recharge; 100% beech maximizes groundwater recharge and minimizes productivity. The optimized provision of both ecosystem services can be reached with a beech share between 20% and 50% in a mixed stand with spruce.
Hence, the results reveal an advantage of monocultures regarding the maximization of single ecosystem services. However, they also point out disadvantages of a monocultural stand. In the simulation study of this paper, even small shares of a secondary tree species considerably increase two ecosystem services, whereas coincidentally only one ecosystem service slightly decreases. Small basal area shares of spruce in beech stands increase productivity and diversity. That increase is being paid for by only a minute decrease of groundwater recharge. In turn, small basal area shares of beech in spruce stands increase groundwater recharge and diversity. This is being paid for by an only slightly decreasing productivity.
Comparing diversity with productivity, we see a trade-off in spruce-dominated stands (100% to 50% spruce) and a synergy in beech-dominated (50% to 100% beech) stands. Furthermore, a change from a synergy to a trade-off is visible comparing diversity with groundwater recharge. In spruce-dominated stands, there is a synergy between diversity and groundwater recharge and in beech-dominated stands, there is a trade-off.
In a real decision making situation, such results based on controlled mixture proportions could be presented to stakeholders. Clearly, it would depend on the stakeholders’ value judgements which mixture proportions they prefer. Productivity-oriented stakeholders like large private forest owners e.g., would typically prefer the spruce monoculture or a 20% beech share at most, because the latter still maintains a high level of wood increment while already profiting from the benefits coming along with having a small share of beech. Multifunctional- and diversity-oriented stakeholders like state forest managers would prefer the 50% mixture because the provision of all ecosystem services considered is the most balanced one. A stakeholder who is responsible for guaranteeing water supply, e.g., a forest owning municipality with own wells, would possibly also prefer the composition with 50% beech shares due to a sparsely decreasing groundwater recharge with higher shares of beech. Coincidentally, the other ecosystem services would decrease with higher beech shares than 50% basal area.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






