Whitebark Pine Prevalence and Ecological Function in Treeline Communities of the Greater Yellowstone Ecosystem, U.S.A.: Potential Disruption by White Pine Blister Rust




Abstract
:
1. Introduction
2. Materials and Methods
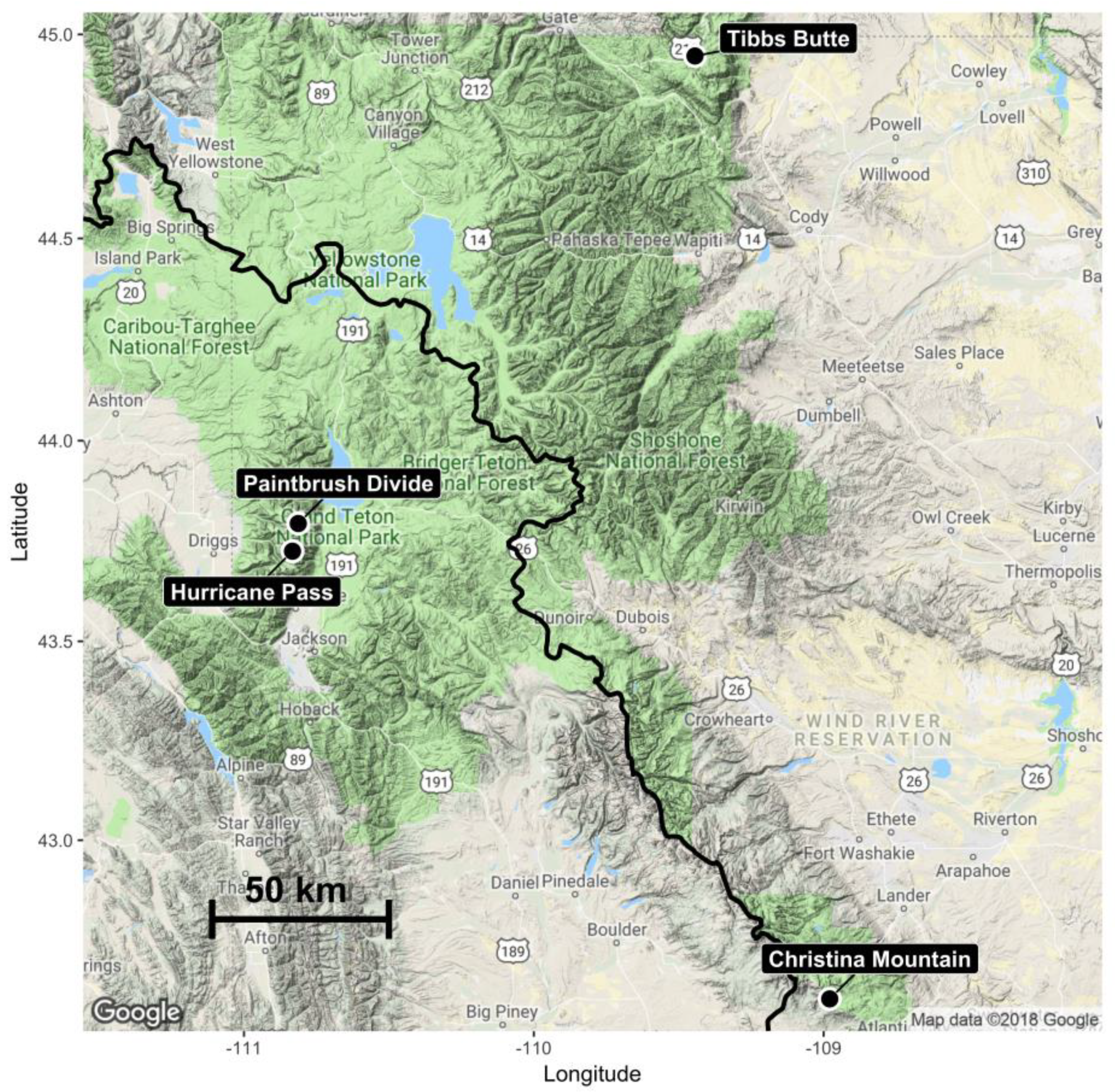
2.1. Study Areas
2.2. Community Assessments
2.3. Mesoclimate Assessments
2.4. Data Analysis
2.4.1. Community Analysis
2.4.2. Mesoclimate Analysis
3. Results
3.1. Community Structure, Composition, and Ecological Roles
3.2. Facilitation by Nurse Objects and Blister Rust Assessments
3.3. Mesoclimate Assessments
4. Discussion
4.1. Community Structure and Composition
4.2. Tree Island Initiation and the Relationship to Species Relative Abundance among Solitary Trees
4.3. Retrospective Evaluation of the Sampling Strategy
4.4. Differences in Nurse Object Facilitation
4.5. The Effects of Mesoclimate on Tree Island Composition
4.6. The Level and Impact of C. ribicola Infection: Management Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Malanson, G.P.; Butler, D.R.; Fagre, D.B.; Walsh, S.J.; Tomback, D.F.; Daniels, L.D.; Resler, L.M.; Smith, W.K.; Weiss, D.J.; Peterson, D.L.; et al. Alpine treeline of Western North America: Linking organism-to-landscape dynamics. Phys. Geogr. 2007, 28, 378–396. [Google Scholar] [CrossRef]
- Geddes, C.A.; Brown, D.G.; Fagre, D.B. Topography and vegetation as predictors of snow water equivalent across the alpine treeline ecotone at Lee Ridge, Glacier National Park, Montana, U.S.A. Arct. Antarct. Alp. Res. 2005, 37, 197–205. [Google Scholar] [CrossRef]
- Tomback, D.F.; Blakeslee, S.C.; Wagner, A.C.; Wunder, M.B.; Resler, L.M.; Pyatt, J.C.; Diaz, S. Whitebark pine facilitation at treeline: Potential interactions for disruption by an invasive pathogen. Ecol. Evol. 2016, 6, 5144–5157. [Google Scholar] [CrossRef] [PubMed]
- Mattson, D. Wilderness-dependent wildlife. Int. J. Wilderness 1997, 3, 34–38. [Google Scholar]
- Bansal, S.; Reinhardt, K.; Germino, M.J. Linking carbon balance to establishment patterns: Comparison of whitebark pine and Engelmann spruce seedlings along an herb cover exposure gradient at treeline. Plant Ecol. 2011, 212, 219–228. [Google Scholar] [CrossRef]
- Körner, C. Alpine Treelines; Springer: New York, NY, USA, 2012; ISBN 978-3-0348-0395-3. [Google Scholar]
- Marr, J.W. The development and movement of tree islands near the upper limit of tree growth in the Southern Rocky Mountains. Ecology 1977, 58, 1159–1164. [Google Scholar] [CrossRef]
- Körner, C.; Paulsen, J.A. World-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Holtmeier, F.K.; Broll, G.; Müterthies, A.; Anschlag, K. Regeneration of trees in the treeline ecotone: Northern Finnish Lapland. Fenn. Int. J. Geogr. 2003, 181, 103–128. [Google Scholar]
- Maher, E.L.; Germino, M.J.; Hasselquist, N.J. Interactive effects of tree and herb cover on survivorship, physiology, and microclimate of conifer seedlings at the alpine tree-line ecotone. Can. J. For. Res. 2005, 35, 567–574. [Google Scholar] [CrossRef]
- Baker, W.L.; Weisberg, P.J. Landscape Analysis of the forest-tundra ecotone in Rocky Mountain National Park, Colorado. Prof. Geogr. 1995, 47, 361–375. [Google Scholar] [CrossRef]
- Resler, L.M.; Shao, Y.; Tomback, D.F.; Malanson, G.P. Predicting functional role and occurrence of whitebark pine (Pinus albicaulis) at alpine treelines: Model accuracy and variable importance. Ann. Assoc. Am. Geogr. 2014, 104, 703–722. [Google Scholar] [CrossRef]
- Sakulich, J. Reconstruction and spatial analysis of alpine treeline in the Elk Mountains, Colorado, USA. Phys. Geogr. 2015, 36, 471–488. [Google Scholar] [CrossRef]
- Tomback, D.F.; Resler, L.M.; Keane, R.E.; Pansing, E.R.; Andrade, A.J.; Wagner, A.C. Community structure, biodiversity, and ecosystem services in treeline whitebark pine communities: Potential impacts from a non-native pathogen. Forests 2016, 7. [Google Scholar] [CrossRef]
- Resler, L.M.; Tomback, D.F. Blister rust prevalence in krummholz whitebark pine: Implications for treeline dynamics, Northern Rocky Mountains, Montana, U.S.A. Arct. Antarct. Alp. Res. 2008, 40, 161–170. [Google Scholar] [CrossRef]
- Callaway, R.M. The detection of neighbors by plants. Trends Ecol. Evol. 2002, 17, 104–105. [Google Scholar] [CrossRef]
- Bertness, M.D.; Callaway, R. Positive interactions in communities. Trends Ecol. Evol. 1994, 9, 191–193. [Google Scholar] [CrossRef]
- Lortie, C.J.; Brooker, R.W.; Choler, P.; Kikvidze, Z.; Michalet, R.; Pugnaire, F.I.; Callaway, R.M. Rethinking plant community theory. Oikos 2004, 107, 433–438. [Google Scholar] [CrossRef]
- Brooker, R.W.; Maestre, F.T.; Callaway, R.M.; Lortie, C.L.; Cavieres, L.A.; Kunstler, G.; Liancourt, P.; Tielbörger, K.; Travis, J.M.J.; Anthelme, F.; et al. Facilitation in plant communities: The past, the present, and the future. J. Ecol. 2008, 96, 18–34. [Google Scholar] [CrossRef]
- Pansing, E.R.; Tomback, D.F.; Wunder, M.B.; French, J.P.; Wagner, A.C. Microsite and elevation zone effects on seed pilferage, germination, and seedling survival during early whitebark pine recruitment. Ecol. Evol. 2017, 7, 9027–9040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resler, L.M. Geomorphic controls of spatial pattern and process at alpine treeline. Prof. Geogr. 2006, 58, 124–138. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Smith, W.K. Seedling occurrence in alpine treeline conifers: A case study from the central Rocky Mountains, USA. Acta Oecol. 1999, 20, 219–224. [Google Scholar] [CrossRef]
- Alftine, K.J.; Malanson, G.P. Directional positive feedback and pattern at an alpine tree line. J. Veg. Sci. 2004, 15. [Google Scholar] [CrossRef]
- Resler, L.; Butler, D.; Malanson, G. Topographic shelter and conifer establishment and mortality in an alpine environment, Glacier National Park, Montana. Phys. Geogr. 2005, 26, 112–125. [Google Scholar] [CrossRef]
- Germino, M.J.; Smith, W.K.; Resor, A.C. Conifer seedling distribution and survival in an alpine-treeline ecotone. Plant Ecol. 2002, 162, 157–168. [Google Scholar] [CrossRef]
- Pyatt, J.C.; Tomback, D.F.; Blakeslee, S.C.; Wunder, M.B.; Resler, L.M.; Boggs, L.A.; Bevency, H.D. The importance of conifers for facilitation at treeline: Comparing biophysical characteristics of leeward microsites in whitebark pine communities. Arct. Antarct. Alp. Res. 2016, 48, 427–444. [Google Scholar] [CrossRef]
- Callaway, R.M. Competition and facilitation on elevation gradients in subalpine forests of the Northern Rocky Mountains, USA. Oikos 1998, 82. [Google Scholar] [CrossRef]
- Renkin, R.A.; Despain, D.G. Fuel moisture, forest type, and lightning-caused fire in Yellowstone National Park. Can. J. For. Res. 1992, 22, 37–45. [Google Scholar] [CrossRef]
- Mellmann-Brown, S. Regeneration of whitebark pine in the timberline ecotone of the Beartooth Plateau, U.S.A.: Spatial distribution and responsible agents. In Mountain Ecosystems: Studies in Treeline Ecology; Broll, G., Keplin, B., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; Volume 86, pp. 97–115. [Google Scholar]
- Tomback, D.F.; Chipman, K.G.; Resler, L.M.; Smith-McKenna, E.K.; Smith, C.M. Relative abundance and functional role of whitebark pine at treeline in the Northern Rocky Mountains. Arct. Antarct. Alp. Res. 2014, 46, 407–418. [Google Scholar] [CrossRef]
- Arno, S.F.; Hoff, R.J. Silvics of Whitebark Pine (Pinus albicaulis); U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1989.
- Tomback, D.F. Dispersal of whitebark pine seeds by Clark’s Nutcracker: A mutualism hypothesis. J. Anim. Ecol. 1982, 51. [Google Scholar] [CrossRef]
- Hutchins, H.E.; Lanner, R.M. The central role of Clark’s nutcracker in the dispersal and establishment of whitebark pine. Oecologia 1982, 55, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Tomback, D.F. Post-fire regeneration of krummholz whitebark pine: A consequence of nutcracker seed caching. Madroño 1986, 33, 100–110. [Google Scholar]
- Lorenz, T.J.; Sullivan, K.A. Seasonal differences in space use by Clark’s Nutcrackers in the Cascade Range. Condor 2009, 111, 326–340. [Google Scholar] [CrossRef]
- Tomback, D.F. Foraging strategies of Clark’s nutcracker. Living Bird 1978, 16, 123–161. [Google Scholar]
- McDonald, G.I.; Hoff, R.J. Blister rust: An introduced plague. In Whitebark Pine Communities: Ecology and Restoration; Tomback, D.F., Arno, S.F., Keane, R.E., Eds.; Island Press: Washington, DC, USA, 2001; pp. 193–220. [Google Scholar]
- Tomback, D.F.; Achuff, P. Blister rust and western forest biodiversity: Ecology, values and outlook for white pines: Blister rust and western forest biodiversity. For. Pathol. 2010, 40, 186–225. [Google Scholar] [CrossRef]
- Schwandt, J.W.; Lockman, I.B.; Kliejunas, J.T.; Muir, J.A. Current health issues and management strategies for white pines in the western United States and Canada: Current health and management strategies. For. Pathol. 2010, 40, 226–250. [Google Scholar] [CrossRef]
- US Fish and Wildlife Service. Endangered and Threatened Wildlife and Plants; 12-Month Finding on a Petition to List Pinus Albicaulis as Endangered or Threatened with Critical Habitat. Available online: https://www.federalregister.gov/documents/2011/07/19/2011-17943/endangered-and-threatened-wildlife-and-plants-12-month-finding-on-a-petition-to-list-pinus (accessed on 19 July 2011).
- Government of Canada. Order Amending Schedule 1 to the Species at Risk Act. Available online: https://www.registrelep-sararegistry.gc.ca/document/default_e.cfm?documentID=2947 (accessed on 18 June 2016).
- Greater Yellowstone Whitebark Pine Monitoring Working Group. Monitoring Whitebark Pine in the Greater Yellowstone Ecosystem. Available online: https://www.fs.fed.us/foresthealth/fhm/posters/posters07/white_pine_bark_yellowstone.pdf (accessed on 10 October 2018).
- Smith-Mckenna, E.K.; Resler, L.M.; Tomback, D.F.; Zhang, H.; Malanson, G.P. Topographic influences on the distribution of white pine blister rust in Pinus albicaulis treeline communities. Écoscience 2013, 20, 215–229. [Google Scholar] [CrossRef]
- Royal Roads University. Google Maps. Available online: https://maps.googleapis.com/maps/api/staticmap?center=43.8,-109.8&zoom=8&size=640x640&scale=2&maptype=terrain&language=en-EN&key=AIzaSyCLSu2p9wprZ6_Uf8qSb1HsE6ENlB_nJGU (accessed on 10 October 2018).
- Kahle, D.; Wickham, H. ggmap: Spatial Visualization with ggplot2. R J. 2013, 5, 144–161. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis (Use R!); Springer: New York, NY, USA, 2009; ISBN 978-0-387-98140-6. [Google Scholar]
- Slowikowski, K. ggrepel: Repulsive Text and Label Geoms for ‘ggplot2′. R. Package Version 0.8.0. Available online: https://cran.r-project.org/web/packages/ggrepel/index.html (accessed on 10 October 2018).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Love, J.D.; Reed, J.C., Jr.; Christiansen, A.C. Geologic Map of Grand Teton National Park, Teton County, Wyoming; U.S. Geological Survey: Teton County, WY, USA, 1992.
- Bevan, A. Summary of the geology of the Beartooth Mountains, Montana. J. Geol. 1923, 31, 441–465. [Google Scholar] [CrossRef]
- Frost, B.R.; Chamberlain, K.R.; Swapp, S.; Frost, C.D.; Hulsebosch, T.P. Late Archean structural and metamorphic history of the Wind River Range: Evidence for a long-lived active margin on the Archean Wyoming craton. Geol. Soc. Am. Bull. 2000, 112, 564–578. [Google Scholar] [CrossRef]
- ESRI. ArcGIS; Environmental Systems Research Institute: Redlands, CA, USA, 2012. [Google Scholar]
- Rowlingson, B.D.; Diggle, P. Splancs: Spatial and Space-Time Point Pattern Analysis—Version 2.01-40. Available online: https://rdrr.io/cran/splancs/ (accessed on 16 April 2017).
- Linhart, Y.B.; Tomback, D.F. Seed dispersal by nutcrackers causes multi-trunk growth form in pines. Oecologia 1985, 67, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Holtmeier, F.-K.; Broll, G. Layering in the Rocky Mountain treeline ecotone: Clonal conifer groups’ distribution, structure, and functional role. Trees 2017, 31, 953–965. [Google Scholar] [CrossRef]
- Burns, K.S.; Schoettle, A.W.; Jacobi, W.R.; Mahalovich, M.F. Options for the Management of White Pine Blister Rust in the Rocky Mountain Region; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ft. Collins, CO, USA, 2008.
- Frich, P.; Alexander, L.V.; Della-Marta, P.; Gleason, B.; Haylock, M.; Klein Tank, A.M.G.; Peterson, T. Observed coherent changes in climatic extremes during the second half of the twentieth century. Clim. Res. 2002, 19, 193–212. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Uria, P.; Körner, C. Low temperature limits of root growth in deciduous and evergreen temperate tree species. Funct. Ecol. 2007, 21, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Calmettes, G.; Drummond, G.B.; Vowler, S.L. Making do with what we have: Use your bootstraps. J. Physiol. 2012, 590, 3403–3406. [Google Scholar] [CrossRef] [PubMed]
- McCune, B. Ecological diversity in North American pines. Am. J. Bot. 1988, 75, 353–368. [Google Scholar] [CrossRef]
- Callaway, R.M.; Sala, A.; Keane, R.E. Succession may maintain high leaf area: Sapwood ratios and productivity in old subalpine forests. Ecosystems 2000, 3, 254–268. [Google Scholar] [CrossRef]
- White, M.A.; Thornton, P.E.; Running, S.W. A continental phenology model for monitoring vegetation responses to interannual climatic variability. Glob. Biogeochem. Cycles 1997, 11, 217–234. [Google Scholar] [CrossRef] [Green Version]
- Manabe, S.; Broccoli, A.J. Mountains and arid climates of middle latitudes. Science 1990, 247, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Griggs, R.F. Timberlines in the Northern Rocky Mountains. Ecology 1938, 19, 548–564. [Google Scholar] [CrossRef]
- Hurlbert, S.H. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 1984, 54, 187–211. [Google Scholar] [CrossRef]
- Little, R.L.; Peterson, D.L.; Conquest, L.L. Regeneration of subalpine fir (Abies lasiocarpa) following fire: Effects of climate and other factors. Can. J. For. Res. 1994, 24, 934–944. [Google Scholar] [CrossRef]
- Bockino, N.K.; Tinker, D.B. Interactions of white pine blister rust and mountain pine beetle in whitebark pine ecosystems in the southern Greater Yellowstone Area. Nat. Areas J. 2012, 32, 31–40. [Google Scholar] [CrossRef]
- Smith, C.M.; Wilson, B.; Rasheed, S.; Walker, R.C.; Carolin, T.; Shepherd, B. Whitebark pine and white pine blister rust in the Rocky Mountains of Canada and northern Montana. Can. J. For. Res. 2008, 38, 982–995. [Google Scholar] [CrossRef]
- Keane, R.E.; Morgan, P.; Menakis, J. Landscape assessment of the decline of whitebark pine (Pinus albicaulis) in the Bob Marshall Wilderness Complex, Montana, USA. Northwest Sci. 1994, 68, 213–229. [Google Scholar]
- Van Arsdel, E. Environment in Relation to White Pine Blister Rust Infection; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 1972; pp. 479–493.
- Ellison, A.W.; Farnsworth, E.J.; Ellison, A.M.; Farnsworth, E.J. Mangrove communities. In Marine Community Ecology; Bertness, M.D., Gaines, S.D., May, M.E., Eds.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2001. [Google Scholar]
- Keane, R.E.; Tomback, D.F.; Aubry, C.A.; Bower, A.D.; Campbell, E.M.; Cripps, C.L.; Jenkins, M.B.; Mahalovich, M.F.; Manning, M.; McKinney, S.T.; et al. A Range-Wide Restoration Strategy for Whitebark Pine (Pinus albicaulis); General Technical Report RMRS-GTR-279; USDA Forest Service: Fort Collins, CO, USA, 2012.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Area | Area (ha) | Elevation (m) | Aspects | Parent Material | Vegetation |
|---|---|---|---|---|---|
| Paintbrush Divide | 121 | 3055–3289 | NE NW SW | Early Proterozoic granite a Quaternary sedimentary deposits a | Arctostaphylos uva-ursi (L.) Spreng Myosotis asiatica (Vestergr.) Schischk. & Serg. Silene acaulis (L.) Jacq. |
| Hurricane Pass | 92 | 3045–3078 | NE SE SW | Middle Cambrian limestone a Early Proterozoic granite a Quaternary sedimentary deposits a | Dryas octopetala L. Silene acaulis Graminoids |
| Tibbs Butte | 4 | 2983–3238 | NE | Precambrian granite b | Geum rossii (R.Br.) Ser. Potentilla diversifolia Lehm. Saxifraga spp. |
| Christina Mountain | 117 | 3200–3400 | NE SE SW | Precambrian granite c Granodiorite c Porphyritic quartz monzonite c Magmatic gneiss c | Geum rossii Pteryxia hendersonii (J.M. Coult & Rose) Mathias & Constance Phlox pulvinata (Wherry) Cronquist Silene acaulis |
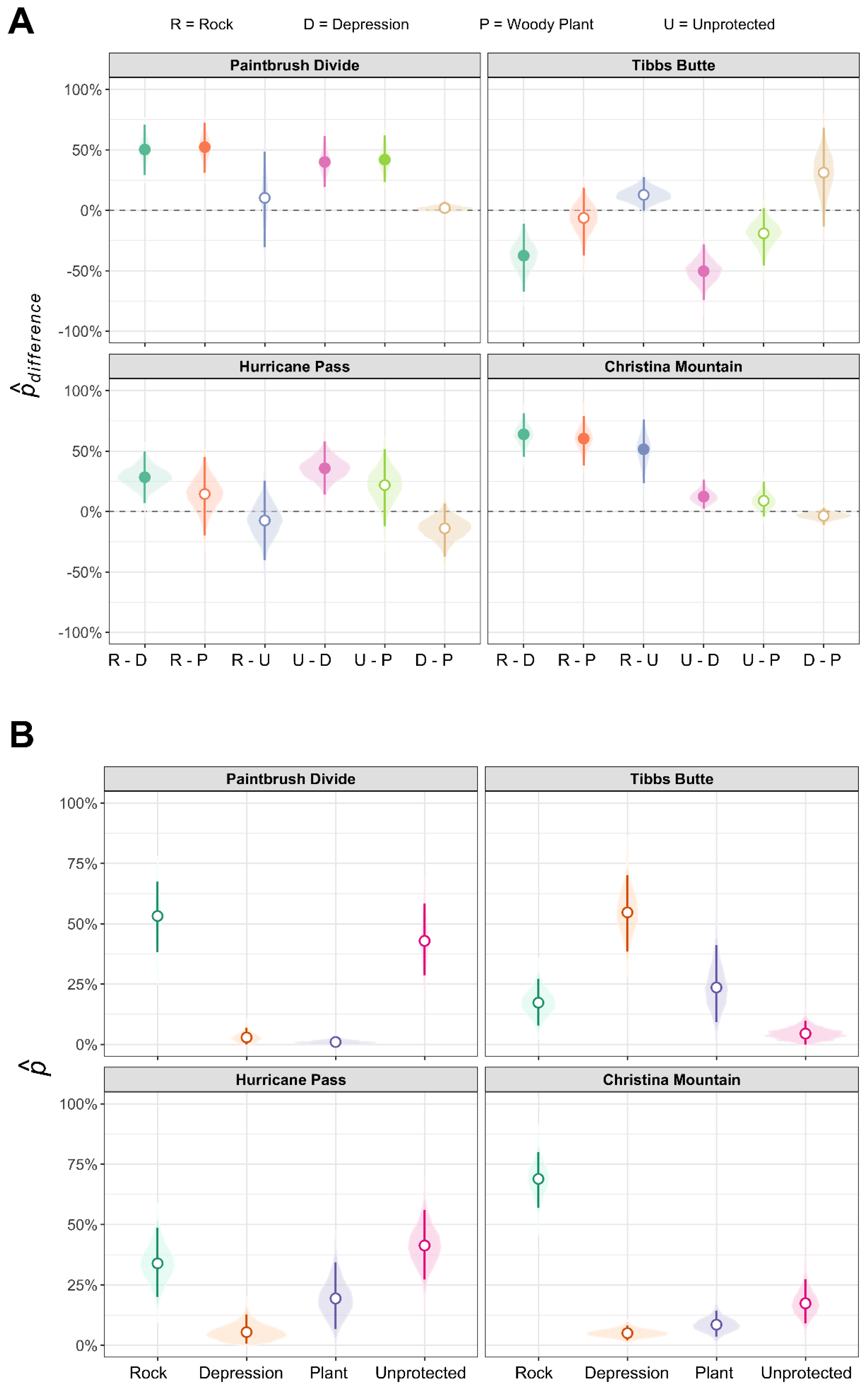
| Study Area | solitary (95% CI) | tree island (95% CI) | solitary-tree island (95% CI) |
|---|---|---|---|
| Paintbrush Divide | 62.0% (48.2%, 75.0%) | 38.0% (25.0%, 51.8%) | 24.1% (−3.6%, 49.9%) |
| Hurricane Pass | 53.8% (40.8%, 67.6%) | 46.2% (32.4%, 59.2%) | 7.6% (−18.4%, 35.1%) |
| Tibbs Butte | 95.5% (88.0%, 100.0%) | 4.5% (0.0%, 12.0%) | 91.0% * (76.0%, 100.0%) |
| Christina Mountain | 74.5% (61.5%, 85.9%) | 25.5% (14.1%, 38.5%) | 48.9% * (23.0%, 71.8%) |
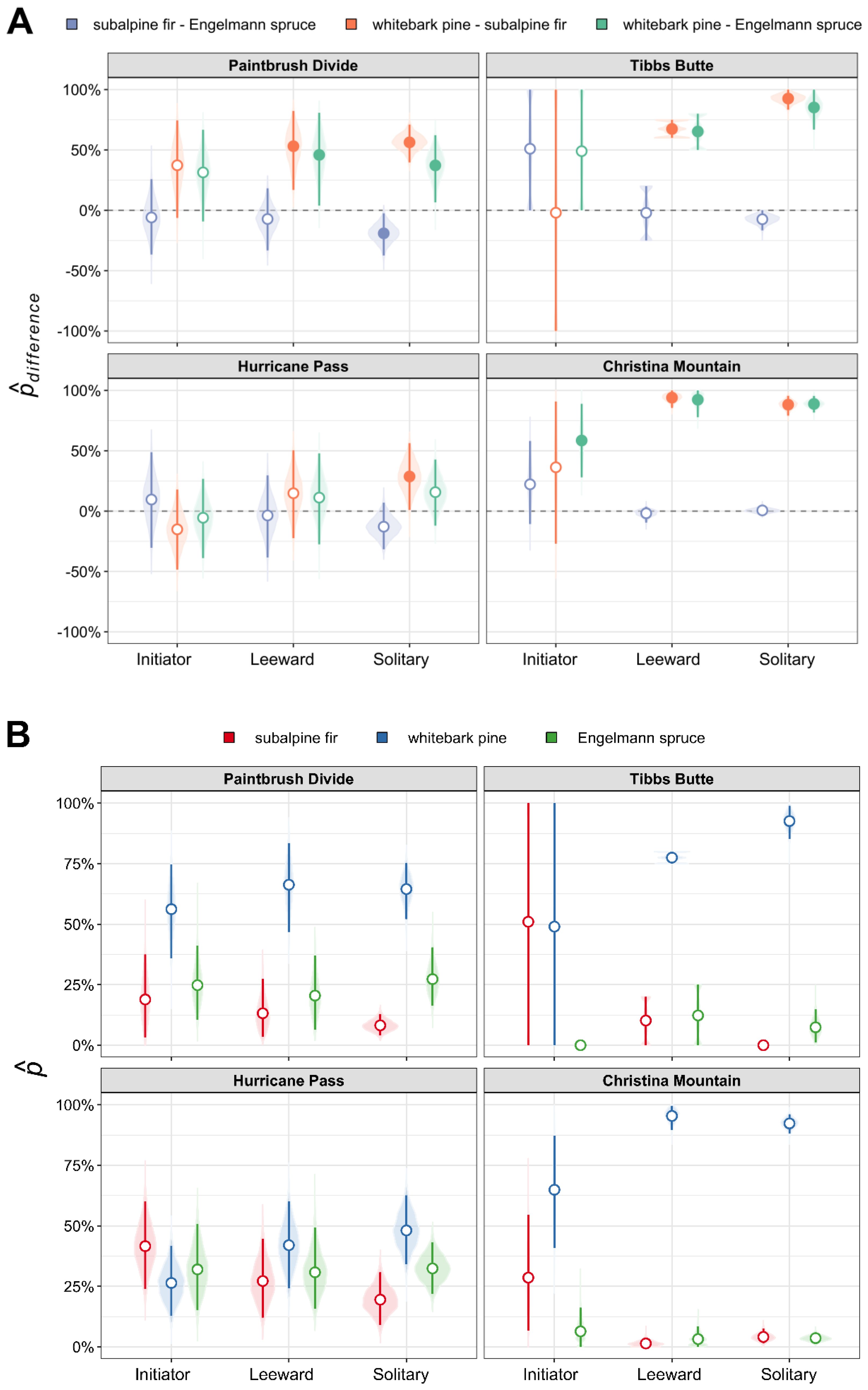
| Study Area | whitebark pine-subalpine fir (98.3% CI) | whitebark pine-Engelmann spruce (98.3% CI) | subalpine fir-Engelmann spruce (98.3% CI) |
|---|---|---|---|
| Paintbrush Divide | 43.4% * (19.1%, 64.0%) | 30.1% * (0.2%, 56.4%) | −13.3% (−31.3%, 3.1%) |
| Hurricane Pass | 16.6% (−4.5%, 38.8%) | 9.4% (−15.0%, 32.7%) | −7.2% (−25.3%, 9.5%) |
| Tibbs Butte | 90.2% * (81.0%, 98.1%) | 83.7% * (65.9%, 97.5%) | −6.6% (−16.1%, 1.7%) |
| Christina Mountain | 83.9% * (72.6%, 92.8%) | 87.1% * (80.4%, 93.2%) | 3.3% (−1.7%, 9.7%) |
| Study Area | whitebark pine (95% CI) | subalpine fir (95% CI) | Engelmann spruce (95% CI) |
|---|---|---|---|
| Paintbrush Divide | 57.9% (43.8%, 70.3%) | 14.4% (7.2%, 22.8%) | 27.7% (17.4%, 39.5%) |
| Hurricane Pass | 42.0% (30.4%, 54.4%) | 25.4% (17.1%, 34.1%) | 32.6% (23.3%, 43.1%) |
| Tibbs Butte | 91.3% (84.3%, 97.5%) | 1.1% (0.0%, 3.1%) | 7.6% (1.4%, 15.0%) |
| Christina Mountain | 90.3% (85.6%, 94.5%) | 6.5% (2.8%, 10.9%) | 3.2% (1.5%, 5.2%) |
| Study Area | Whitebark Pine initiator-solitary (95% CI) | Subalpine Fir initiator-solitary (95% CI) | Engelmann Spruce initiator-solitary (95% CI) |
|---|---|---|---|
| Paintbrush Divide | −1.3% (−23.9%, 20.1%) | 4.1% (−5.3%, 16.7%) | −2.8% (−19.2%, 15.9%) |
| Hurricane Pass | −20.9% (−39.9%, −3.1%) * | 17.6% (2.5%, 32.8%) * | 3.3% (−16.7%, 24.5%) |
| Tibbs Butte | −51.0% (−100.0%, 0.0%) | 51.0% (0.0%, 100.0%) | 0.0% (0.0%, 0.0%) |
| Christina Mountain | −15.3% (−38.2%, 4.3%) | 12.4% (−5.6%, 36.8%) | 3.0% (−5.6%, 13.4%) |
| Study Area | (95% CI) | (95% CI) | (95% CI) |
|---|---|---|---|
| Paintbrush Divide | 136 days (129 days, 142 days) | 9.9 °C (9.5 °C, 10.3 °C) | 27.6 cm (23.8 cm, 31.5 cm) |
| Hurricane Pass | 128 days (120 days, 136 days) | 9.0 °C (8.5 °C, 9.4 °C) | 28.1 cm (23.4 cm, 33.0 cm) |
| Tibbs Butte | 115 days (107 days, 123 days) | 8.7 °C (8.3 °C, 9.2 °C) | 19.2 cm (16.3 cm, 22.1 cm) |
| Christina Mountain | 136 days (129 days, 143 days) | 9.1 °C (8.7 °C, 9.5 °C) | 19.4 cm (16.3 cm, 22.5 cm) |
| Comparison | (99.2% CI) | (99.2% CI) | (99.2% CI) |
|---|---|---|---|
| PD-HP | 8 days (−6 days, 21 days) | 1.0 °C * (0.1 °C, 1.8 °C) | −0.5 cm (−9.1 cm, 7.5 cm) |
| PD-TB | 21 days * (6 days, 34 days) | 1.2 °C * (0.4 °C, 1.9 °C) | 8.5 cm * (2.2 cm, 15.1 cm) |
| PD-CM | 0 days (−13 days, 13 days) | 0.8 °C (0.0 °C, 1.5 °C) | 8.3 cm * (1.7 cm, 14.9 cm) |
| CM-HP | 8 days (−6 days, 22 days) | 0.2 °C (−0.6 °C, 1.0 °C) | −8.8 cm * (−16.8 cm, −1.4 cm) |
| CM-TB | 21 days * (6 days, 35 days) | 0.4 °C (−0.4 °C, 1.2 °C) | 0.2 cm (−5.3 cm, 5.7 cm) |
| HP-TB | 13 days (−2 days, 28 days) | 0.2 °C (−0.6 °C, 1.0 °C) | 8.9 cm * (1.7 cm, 16.5 cm) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner, A.C.; Tomback, D.F.; Resler, L.M.; Pansing, E.R. Whitebark Pine Prevalence and Ecological Function in Treeline Communities of the Greater Yellowstone Ecosystem, U.S.A.: Potential Disruption by White Pine Blister Rust. Forests 2018, 9, 635. https://doi.org/10.3390/f9100635
Wagner AC, Tomback DF, Resler LM, Pansing ER. Whitebark Pine Prevalence and Ecological Function in Treeline Communities of the Greater Yellowstone Ecosystem, U.S.A.: Potential Disruption by White Pine Blister Rust. Forests. 2018; 9(10):635. https://doi.org/10.3390/f9100635
Chicago/Turabian StyleWagner, Aaron C., Diana F. Tomback, Lynn M. Resler, and Elizabeth R. Pansing. 2018. "Whitebark Pine Prevalence and Ecological Function in Treeline Communities of the Greater Yellowstone Ecosystem, U.S.A.: Potential Disruption by White Pine Blister Rust" Forests 9, no. 10: 635. https://doi.org/10.3390/f9100635
APA StyleWagner, A. C., Tomback, D. F., Resler, L. M., & Pansing, E. R. (2018). Whitebark Pine Prevalence and Ecological Function in Treeline Communities of the Greater Yellowstone Ecosystem, U.S.A.: Potential Disruption by White Pine Blister Rust. Forests, 9(10), 635. https://doi.org/10.3390/f9100635