Fungal Communities Associated with Bark Beetles in Pinus radiata Plantations in Northern Spain Affected by Pine Pitch Canker, with Special Focus on Fusarium Species

 , , and
, , and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Molecular and Morphological Identification of Fungi
2.3. Molecular Identification of Insects
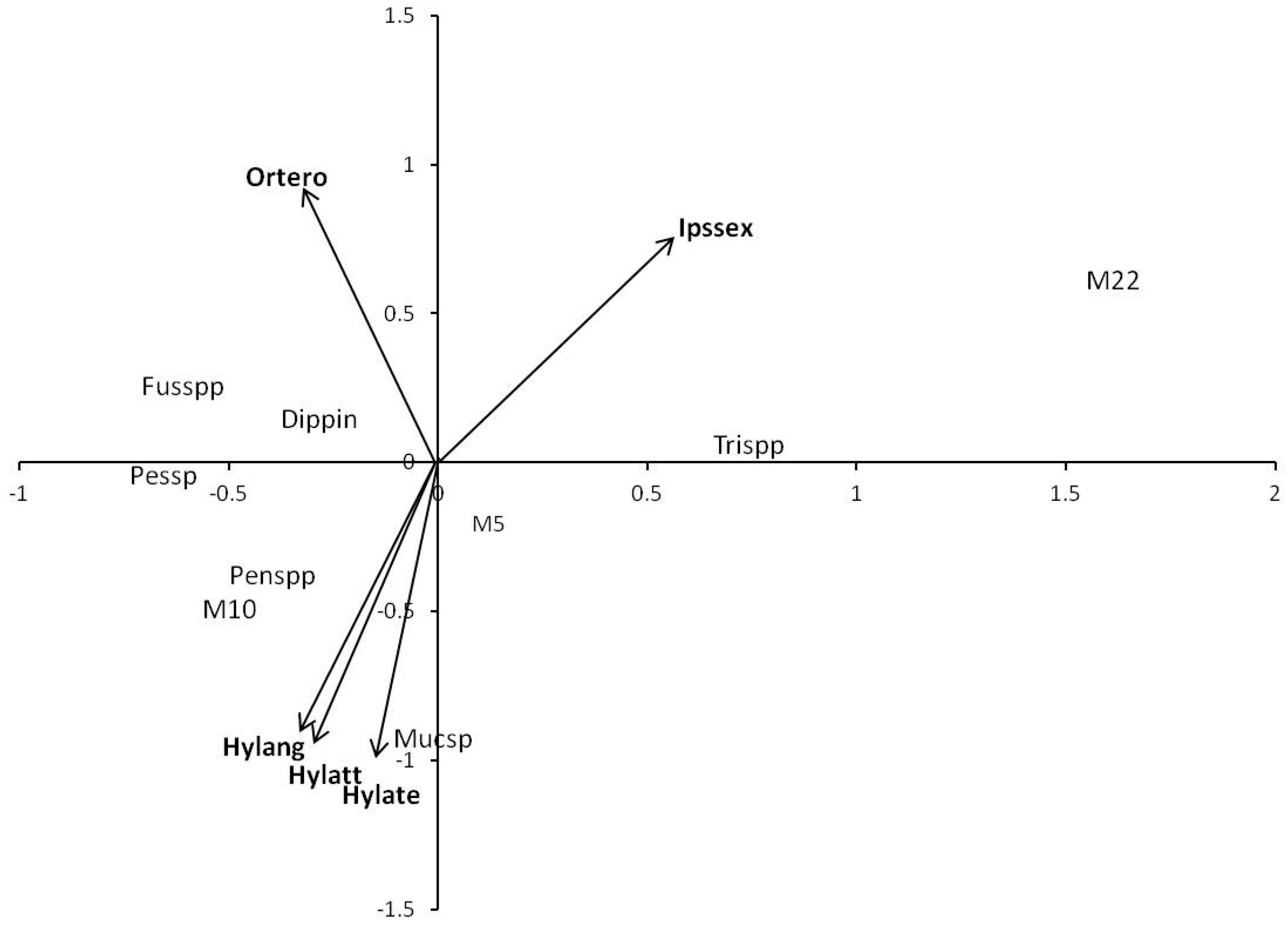
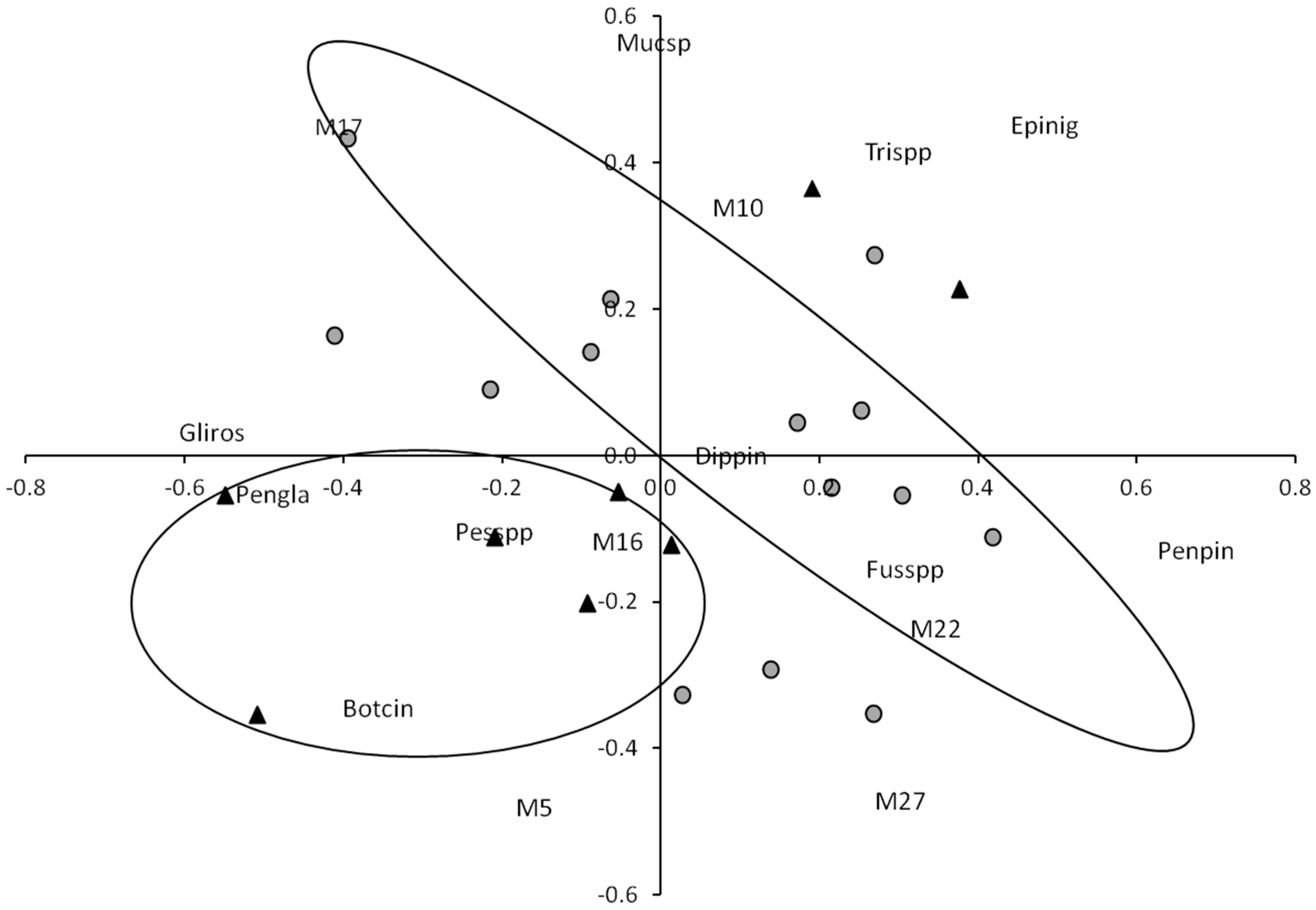
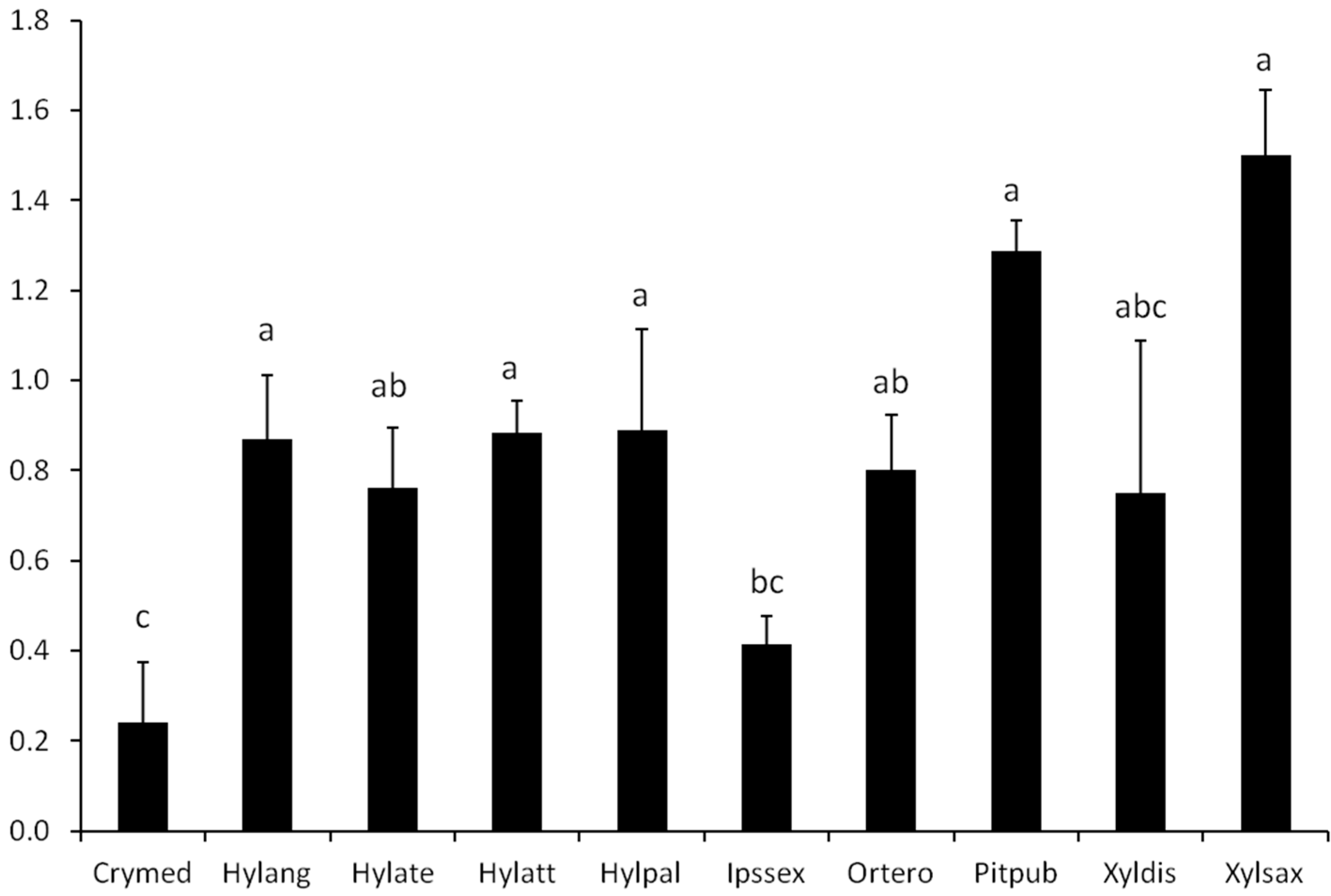
2.4. Statistical Analysis
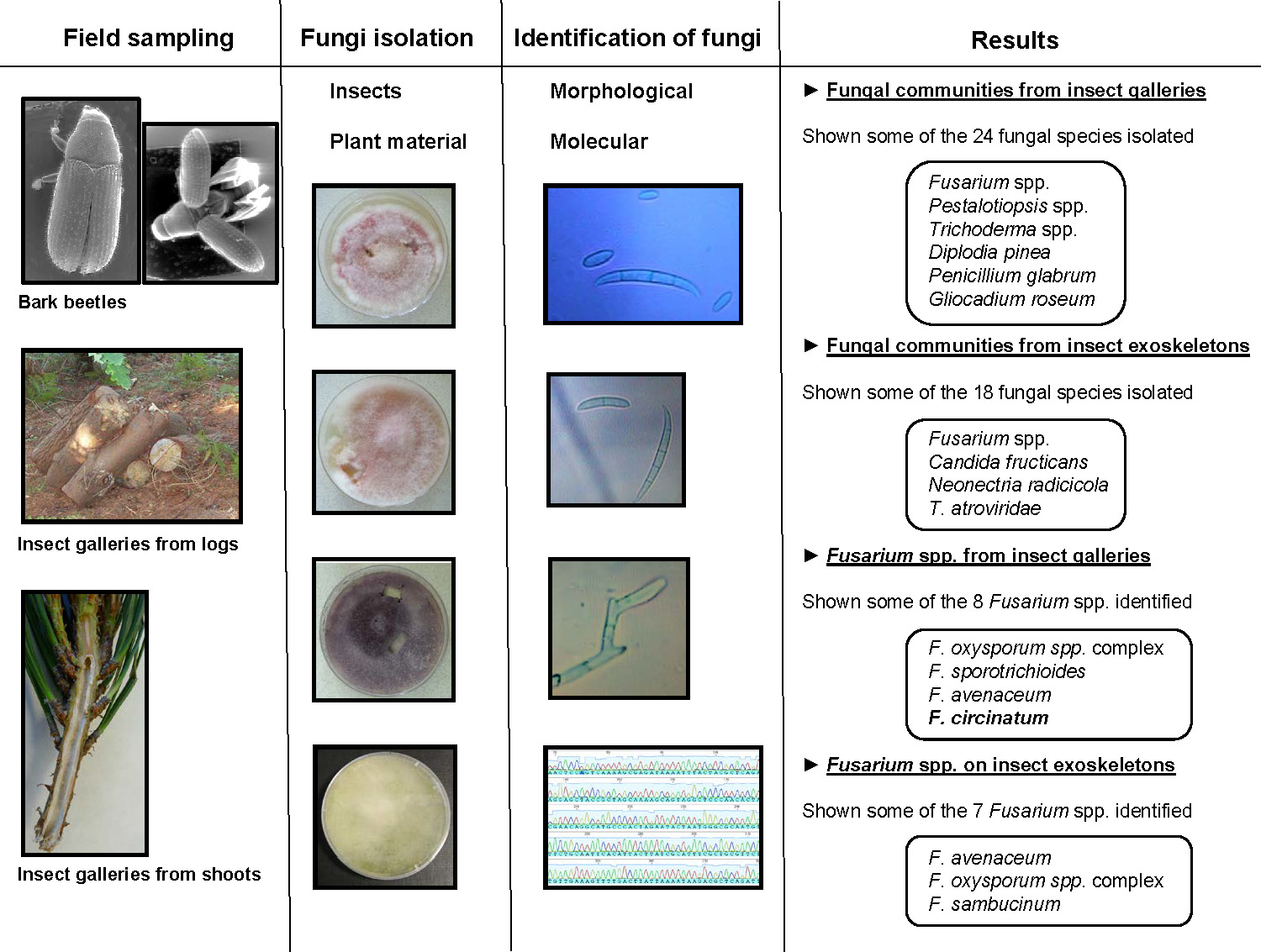
3. Results
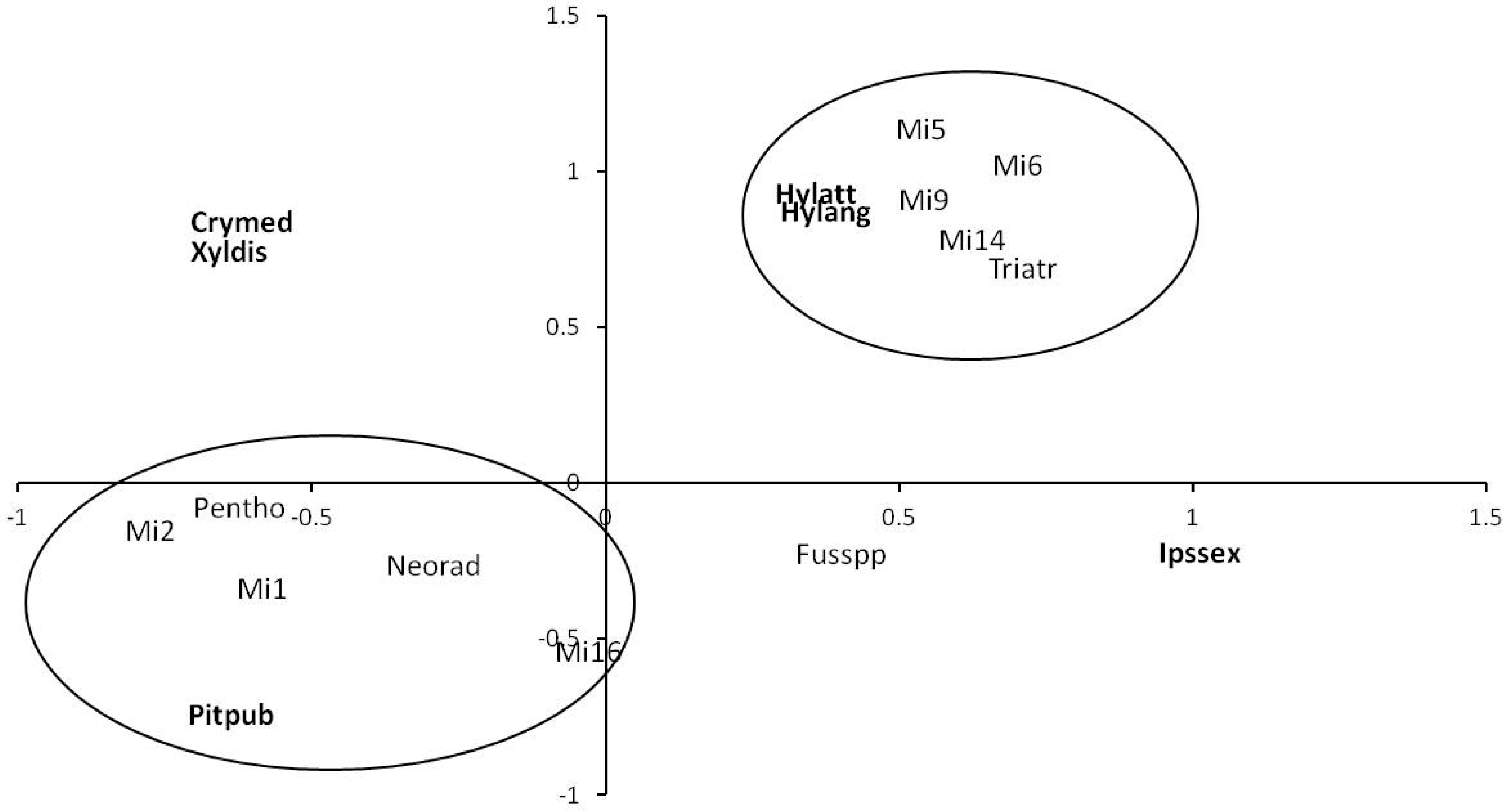
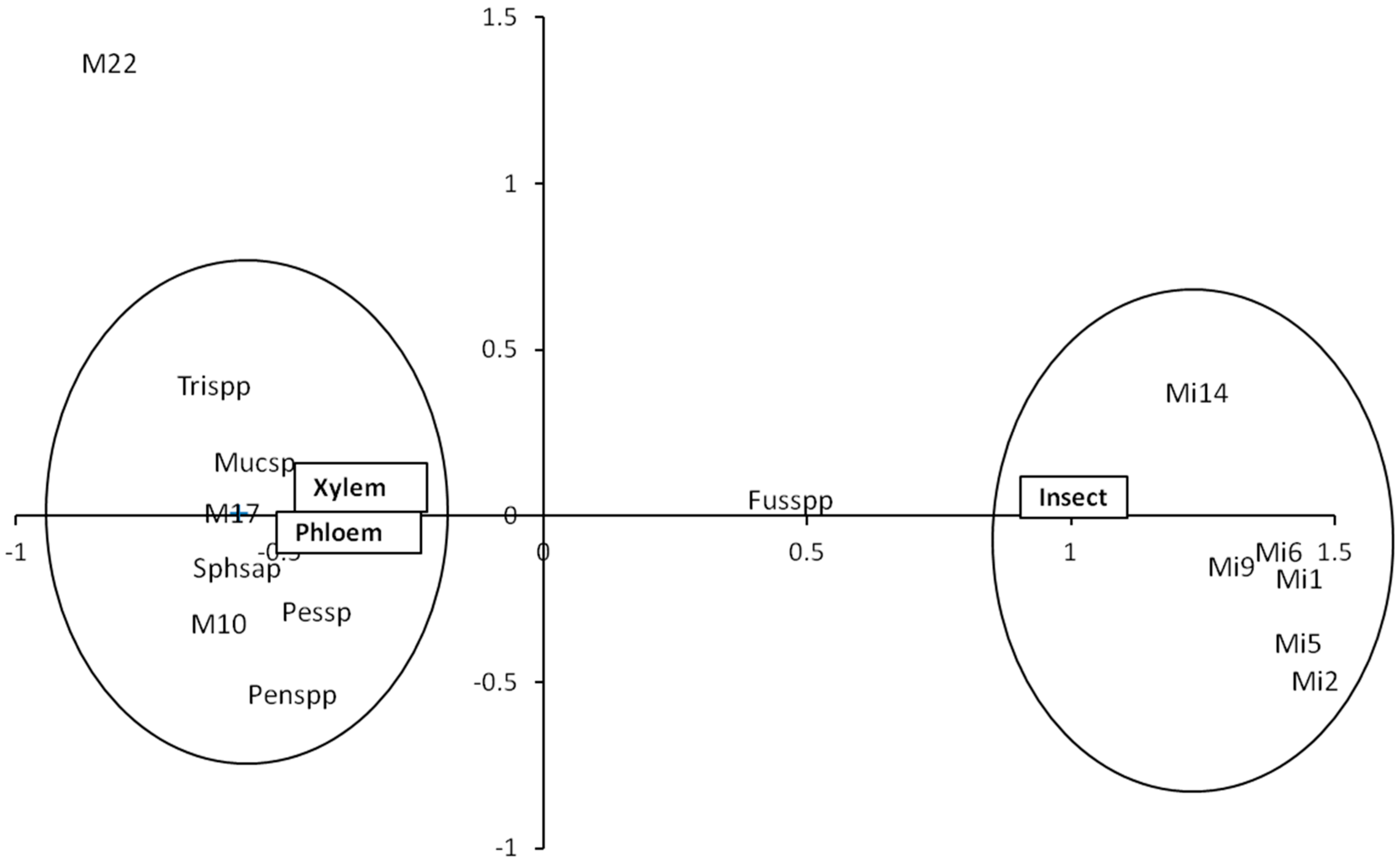
3.1. Fungal Communities from Insect Galleries
3.2. Fungal Communities from Insect Exoskeletons
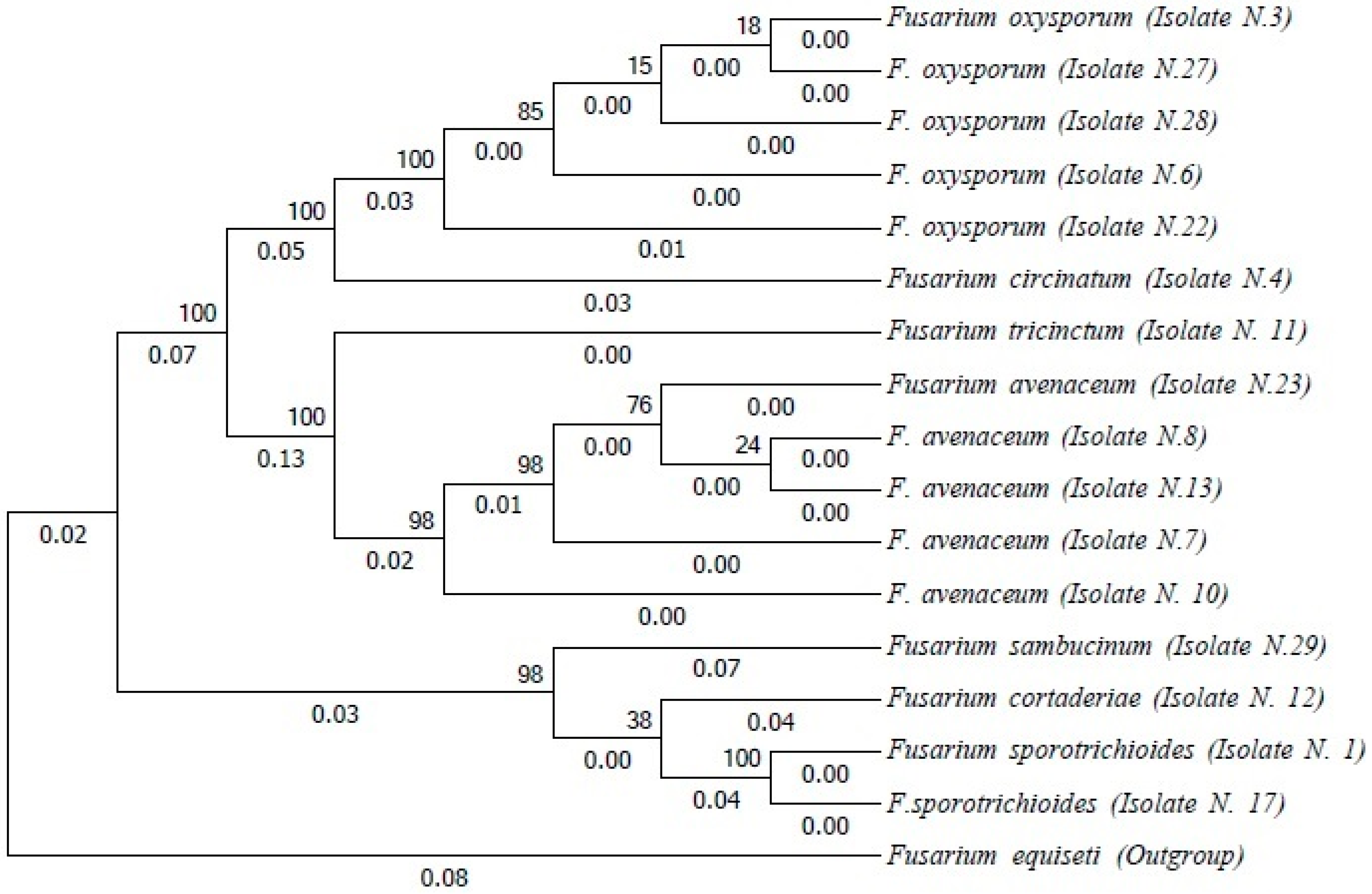
3.3. Fusarium spp. from Insect Galleries
3.4. Fusarium spp. on Insect Exoskeletons
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arnold, A.E. Understanding the diversity of foliar endophytic fungi: Progress, challenges, and frontiers. Fungal Biol. Rev. 2007, 21, 51–66. [Google Scholar] [CrossRef]
- Teetor-Barsch, G.H.; Roberts, D.W. Entomogenous Fusarium species. Mycopatbologia 1983, 84, 3–16. [Google Scholar] [CrossRef]
- Summerell, B.; Leslie, J. Introducing the genus Fusarium. In Control of Fusarium Species; Álves-Santos, F., Diez Casero, J., Eds.; Research Point: Kerala, India, 2012. [Google Scholar]
- Mirete, S.; Patiño, B.; Vázquez, C.; Jiménez, M.; Hinojo, M.J.; Soldevilla, C.; González-Jaén, M.T. Fumonisin production by Gibberella fujikuroi strains from Pinus species. Int. J. Food Microbiol. 2003, 89, 213–221. [Google Scholar] [CrossRef]
- Summerell, B.; Leslie, J. Introducing the genus Fusarium. In Control of Fusarium Diseases; Alves-Santos, F., Diez, J., Eds.; Research Signpoin: Kerala, India, 2012; pp. 1–16. [Google Scholar]
- Román-Avilés, B.; Lewis, J.; Kelly, J. Fusarium genetic control: A long term strategy. In Control of Fusarium Diseases; Alves-Santos, F., Díez, J., Eds.; Research Sing Post: Kerala, India, 2011; pp. 65–108. [Google Scholar]
- Nirenberg, H.I.; O’Donnell, K. New Fusarium species and combinations within the Gibberella fujikuroi species complex. Mycologia 1998, 90, 434–458. [Google Scholar] [CrossRef]
- Wikler, K.; Storer, A.J.; Newman, W.; Gordon, T.R.; Wood, D.L. The dynamics of an introduced pathogen in a native Monterey pine (Pinus radiata) forest. For. Ecol. Manag. 2003, 179, 209–221. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Hammerbacher, A.; Ganley, R.J.; Steenkamp, E.T.; Gordon, T.R.; Wingfield, B.D.; Coutinho, T.A. Pitch canker caused by Fusarium circinatum—A growing threat to pine plantations and forests worldwide. Australas. Plant Pathol. 2008, 37, 319–334. [Google Scholar] [CrossRef]
- Viljoen, A.; Wingfield, M.J.; Kemp, G.H.J.; Marasas, W.F.O. Susceptibility of pines in South Africa to the pitch canker fungus Fusarium subglutinans f.sp. pini. Plant Pathol. 2018, 44, 877–882. [Google Scholar] [CrossRef]
- Bezos, D.; Martínez-Alvarez, P.; Fernández, M.; Diez, J.J. Epidemiology and management of pine pitch canker disease in Europe—A review. Balt. For. 2017, 23, 279–293. [Google Scholar]
- Gordon, T.R.; Storer, A.J.; Wood, D.L. The pitch canker epidemic in California. Plant Dis. 2001, 85, 1128–1139. [Google Scholar] [CrossRef]
- Landeras, E.; García, P.; Fernández, Y.; Braña, M.; Fernández-Alonso, O.; Méndez-Lodos, S.; Pérez-Sierra, A.; León, M.; Abad-Campos, P.; Berbegal, M.; et al. Outbreak of pitch canker caused by Fusarium circinatum on Pinus spp. in Northern Spain. Plant Dis. 2005, 89, 1015. [Google Scholar] [CrossRef]
- Romón, P.; Troya, M.; Fernández de Gamarra, M.E.; Eguzkitza, A.; Iturrondobeitia, J.C.; Goldarazena, A. Fungal communities associated with pitch canker disease of Pinus radiata caused by Fusarium circinatum in northern Spain: Association with insects and pathogen-saprophyte antagonistic interactions. Can. J. Plant Pathol. 2008, 30, 241–253. [Google Scholar] [CrossRef]
- Alves-Santos, F.; Diez, J. Biological control of Fusarium. In Control of Fusarium Diseases; Alves-Santos, F., Diez, J., Eds.; Research Singpoin: Kerala, India, 2012; pp. 131–158. [Google Scholar]
- Martínez-Álvarez, P.; Manuel Alves-Santos, F.; Martínez-Álvarez, J.D. In vitro and in vivo interactions between Trichoderma viride and Fusarium circinatum. Silva Fenn. 2012, 46, 303–316. [Google Scholar] [CrossRef]
- Martín-García, J.; Paraschiv, M.; Flores-Pacheco, J.A.; Chira, D.; Diez, J.J.; Fernández, M. Susceptibility of several northeastern conifers to Fusarium circinatum and strategies for biocontrol. Forests 2017, 8, 318. [Google Scholar] [CrossRef]
- Howell, C.R. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef]
- Deacon, J.W. Fungal Biology; Blackwell Publishing: Malden, MA, USA, 2006. [Google Scholar]
- García-Serna, I. Diplodia pinea (Desm.) Kickx y Fusarium circinatum Niremberg & O’Donell; Universidad del País Vasco: Lejona, Spain, 2011. [Google Scholar]
- Romón, P.; Iturrondobeitia, J.C.; Gibson, K.; Lindgren, B.S.; Goldarazena, A. Quantitative association of bark beetles with pitch canker fungus and effects of verbenone on their semiochemical communication in monterey pine forests in Northern Spain. Environ. Entomol. 2007, 36, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Bezos, D.; Martínez-Álvarez, P.; Diez, J.J.; Fernández, M.M. The pine shoot beetle Tomicus piniperda as a plausible vector of Fusarium circinatum in northern Spain. Ann. For. Sci. 2015, 72, 1079–1088. [Google Scholar] [CrossRef]
- Bezos, D.; Martínez-Álvarez, P.; Diez, J.J.; Fernández, M. Association levels between Pityophthorus pubescens and Fusarium circinatum in pitch canker disease affected plantations in northern Spain. Entomol. Gen. 2016, 36, 43–54. [Google Scholar] [CrossRef]
- Sakamoto, J.M.; Gordon, T.R.; Storer, A.J.; Wood, D.L. The role of Pityophthorus spp. as vectors of pitch canker affecting Pinus radiata. Can. Entomol. 2007, 139, 864–871. [Google Scholar] [CrossRef]
- López, S.; Ochoa, P.R.; Carlos Iturrondobeitia, J.; Goldarazena, A. Los Escolítidos de las Coníferas del País Vasco; Servicio Central de Publicaciones del Gobierno Vasco: Vitoria-Gasteiz, Spain, 2007; ISBN 978-84-457-2650-1. [Google Scholar]
- Raffa, K.; Berryman, A. The role of host plant resistance in the colonization behaviour and ecology of bark beetles (Coleoptera: Scolitidae). Ecol. Monogr. 1983, 53, 27–49. [Google Scholar] [CrossRef]
- López, S.; Quero, C.; Iturrondobeitia, J.C.; Guerrero, Á.; Goldarazena, A. Evidence for (E)-pityol as an aggregation pheromone of Pityophthorus pubescens (Coleoptera: Curculionidae: Scolytinae). Can. Entomol. 2011, 143, 447–454. [Google Scholar] [CrossRef]
- Lacap, D.C.; Hyde, K.D.; Liew, E.C.Y. An evaluation of the fungal “morphotype” concept based on ribosomal DNA sequences. Fungal Divers. 2003, 12, 53–66. [Google Scholar]
- Vainio, E.J.; Korhonen, K.; Hantule, J. Genetic variation in Phlebiopsis gigantea as detected with random amplified microsatellite (RAMS) markers. Mycol. Res. 1998, 102, 187–192. [Google Scholar] [CrossRef]
- Gardes, M.; Burns, T. ITS primers with enhanced specificity for basidiomycetes application to the identification of mycorrhizae and rust. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Leslie, J.; Summerell, B. The Fusarium Laboratury Manual; Blackwell Publishing: Ames, IA, USA, 2006; ISBN 978-0-813-81919-8. [Google Scholar]
- O’donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed]
- Geiser, D.M.; Jiménez-Gasco, M.D.M.; Kang, S.; Makalowska, I.; Veeraraghavan, N.; Ward, T.J.; Zhang, N.; Kuldau, G.A.; O’donnell, K. FUSARIUM-ID v. 1.0: A DNA sequence database for identifying Fusarium. Eur. J. Plant Pathol. 2004, 110, 473–479. [Google Scholar] [CrossRef]
- Pérez-Sierra, A.; Landeras, E.; León, M.; Berbegal, M.; García-Jiménez, J.; Armengol, J. Characterization of Fusarium circinatum from Pinus spp. in northern Spain. Mycol. Res. 2007, 111, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence Limits on phylogenies: An aproach using the botts trap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980. [Google Scholar] [CrossRef]
- Schweigkofler, W.; O’Donnell, K.; Garbelotto, M. Detection and quantification of airborne conidia of Fusarium circinatum, the causal agent of pine pitch canker, from two California sites by using a real-time PCR approach combined with a simple spore trapping method. Appl. Environ. Microbiol. 2004, 70, 3512–3520. [Google Scholar] [CrossRef] [PubMed]
- Gallego, D.; Galián, J. The internal transcribed spacers (ITS1 and ITS2) of the rDNA differentiates the bark beetle forest pests Tomicus destruens and T. piniperda. Insect Mol. Biol. 2001. [Google Scholar] [CrossRef]
- García Pérez, A. Métodos Avanzados De Estadística Aplicada: Métodos Robustos Y De Remuestreo; UNED: Madrid, Spain, 2010. [Google Scholar]
- Wilcox, R.R.; Schönbrodt, F.D. The WRS Package for Robust Statistics in R (Version 0.24). 2014. Available online: https://github.com/nicebread/WRS (accessed on 9 November 2018).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Henry, M.; Stevens, H.; et al. Package “vegan” Title Community Ecology Package. 2015. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 9 November 2018).
- Whitehill, J.G.A.; Lehman, J.S.; Bonello, P. Ips pini (Curculionidae: Scolytinae) is a vector of the fungal pathogen, Sphaeropsis sapinea (Coelomycetes), to Austrian Pines, Pinus nigra (Pinaceae). Environ. Entomol. 2007, 36, 114–120. [Google Scholar] [CrossRef]
- Zamora, P.; Martínez-Ruiz, C.; Diez, J.J. Fungi in needles and twigs of pine plantations from northern Spain. Fungal Divers. 2008, 30, 171–184. [Google Scholar]
- Sanz-Ros, A.V.; Müller, M.M.; San Martín, R.; Diez, J.J. Fungal endophytic communities on twigs of fast and slow growing Scots pine (Pinus sylvestris L.) in northern Spain. Fungal Biol. 2015, 119, 870–883. [Google Scholar] [CrossRef] [PubMed]
- Jeewon, R.; Liew, E.; Hyde, K. Phylogenetic evaluation of species nomenclature of Pestalotiopsis in relation to host association. Fungal Divers. 2004, 17, 39–55. [Google Scholar]
- Bajo, J.; Santamaría, O.; Diez, J.J. Cultural characteristics and pathogenicity of Pestalotiopsis funerea on Cupressus arizonica. For. Pathol. 2008, 38, 263–274. [Google Scholar] [CrossRef]
- Pinto, P.M.; Alonso, J.A.P.; Fernández, V.P.; Casero, J.J.D. Fungi isolated from diseased nursery seedlings in Spain. New For. 2006, 31, 41–56. [Google Scholar] [CrossRef]
- Olmedo-Monfil, V.; Mendoza-Mendoza, A.; Gómez, I.; Cortés, C.; Herrera-Estrella, A. Multiple environmental signals determine the transcriptional activation of the mycoparasitism related gene prb1 in Trichoderma atroviride. Mol. Genet. Genom. 2002, 267, 703–712. [Google Scholar] [CrossRef]
- Hofstetter, R.W.; Dinkins-Bookwalter, J.; Davis, T.S.; Klepzig, K.D. Symbiotic Associations of Bark Beetles. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F., Hostetter, R., Eds.; Academic Press: Cambridge, MA, USA, 2015; ISBN 9780124171565. [Google Scholar]
- Ndobe, N.E. Fungi Associated with Roots of Healthy-Looking Scots Pines and Norway Spruce Seedlings Grown in Nine Swedish Forest Nurseries; SLU: Uppsala, Sweden, 2012. [Google Scholar]
- Talgø, V.; Thomsen, I.M.; Nielsen, U.B.; Brurberg, M.B.; Stensvand, A. Neonectria canker on subalpine fir (Abies lasiocarpa) in Denmark. In Proceedings of the 10th International Christmas Tree Research & Extension Conference, Eichgraben, Austria, 21–27 August 2011; pp. 92–96. [Google Scholar]
- Morales, R. Detección de Neonectria fuckeliana en Chile, asociado a cancros y malformaciones fustales en plantaciones de Pinus radiata. Bosque 2009, 30, 106–110. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Rojas, M.G.; Sittertz-Bhatkar, H.; Saldaña, G.; Saldaña, S. Symbiotic Relationship Between Hypothenemus hampei (Coleoptera: Scolytidae) and Fusarium solani (Moniliales: Tuberculariaceae). Ann. Entomol. Soc. Am. 2000, 93, 541–547. [Google Scholar] [CrossRef]
- Dick, M.A.; Dobbie, K. Species of Fusarium on Pinus radiata in New Zeland. N. Z. Plant Prot. 2002, 55, 58–62. [Google Scholar]
- Satyaprasad, K.; Bateman, G.L.; Ward, E. Comparisons of isolates of Fusarium avenaceum from white lupin and other crops by pathogenicity tests, DNA analyses and vegetative compatibility tests. J. Phytopathol. 2000, 148, 211–219. [Google Scholar] [CrossRef]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Mycotoxins Plant Dis. 2002, 108, 611–624. [Google Scholar]
- Niemira, B.A.; Hammerschmidt, R.; Safir, G.R. Postharvest suppression of potato dry rot (Fusarium sambucinum) in prenuclear minitubers by arbuscular mycorrhizal fungal inoculum. Am. Potato J. 1996, 73, 509–515. [Google Scholar] [CrossRef]
- Muñoz-Adalia, E.J.; Sanz-Ros, A.V.; Flores-Pacheco, J.A.; Hantula, J.; Diez, J.J.; Vainio, E.J.; Fernández, M. Sydowia polyspora dominates fungal communities carried by two Tomicus species in pine plantations threatened by Fusarium circinatum. Forests 2017, 8, 127. [Google Scholar] [CrossRef]
- Lieutier, F.; Langstrom, B.; Faccoli, M. The genus Tomicus. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F., Hofstetter, R., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 371–426. ISBN 9780124171565. [Google Scholar]
- Jacobs, K.; Bergdahl, D.R.; Wingfield, M.J.; Halik, S.; Seifert, K.A.; Bright, D.E.; Wingfield, B.D. Leptographium wingfieldii introduced into North America and found associated with exotic Tomicus piniperda and native bark beetles. Mycol. Res. 2004, 108, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Lieutier, F.; Yart, A.; Garcia, J.; Ham, M.C.; Morelet, M.; Levieux, J. Champignons phytopathogènes associés à deux coléoptères scolytidae du pin sylvestre (Pinus sylvestris L.) et étude préliminaire de leur agressivité envers l’hôte. Ann. Sci. For. 1989, 46, 201–216. [Google Scholar] [CrossRef]
- Reay, S.D.; Walsh, P.J.; Ram, A.; Farrell, R.L. The invasion of Pinus radiata seedlings by sapstain fungi, following attack by the black pine bark beetle, Hylastes ater (Coleoptera: Scolytidae). For. Ecol. Manag. 2002, 165, 47–56. [Google Scholar] [CrossRef]
- Bonello, P.; Mcnee, W.R.; Storer, A.J.; Wood, D.L.; Gordon, T.R. The role of olfactory stimuli in the location of weakened hosts by twig-infesting Pityophthorus spp. Ecol. Entomol. 2001, 26, 8–15. [Google Scholar] [CrossRef]
- Etxebeste, I.; Pajares, J.A. Verbenone protects pine trees from colonization by the six-toothed pine bark beetle, Ips sexdentatus Boern. (Col.: Scolytinae). J. Appl. Entomol. 2011, 135, 258–268. [Google Scholar] [CrossRef]
- Fox, J.W.; Wood, D.L.; Koehler, C.S.; O’keefe, S.T. Engraver beetles (Scolytidae: Ips species) as vectors of the pitch canker fungus, Fusarium subglutinans. Can. Entomol. 1991, 123, 1355–1367. [Google Scholar] [CrossRef]
- Lieutier, F.; Yart, A.; Salle, A. Stimulation of tree defenses by Ophiostomatoid fungi can explain attack success of bark beetles on conifers. Ann. For. Sci. 2009, 66, 801. [Google Scholar] [CrossRef]
- Six, D.L.; Wingfield, M.J. The role of phytopathogenicity in bark beetle–fungus symbioses: A challenge to the classic paradigm. Annu. Rev. Entomol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.M.; Norris, D.M. A complex of fungi mutualistically involved in the nutrition of the ambrosia beetle Xyleborus ferrugineus. J. Invertebr. Pathol. 1968, 11, 246–250. [Google Scholar] [CrossRef]
- Dvořák, M.; Janoš, P.; Botella, L.; Rotková, G.; Zas, R. Spore dispersal patterns of Fusarium circinatum on an infested monterey pine forest in north-western Spain. Forests 2017, 8, 432. [Google Scholar] [CrossRef]

 = plots sampled during autumn, ▲ = plots sampled during summer).
= plots sampled during autumn, ▲ = plots sampled during summer).
= plots sampled during autumn, ▲ = plots sampled during summer).
= plots sampled during autumn, ▲ = plots sampled during summer).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insect Species | Total Number | Logs | Ethanol | (E)-Pityol | Shoots |
|---|---|---|---|---|---|
| Ips sexdentatus | 116 | 116 | 0 | 0 | 0 |
| Pityophthorus pubescens | 97 | 0 | 0 | 97 | 0 |
| Hylastes attenuatus Erichson | 86 | 83 | 2 | 0 | 1 |
| Orthotomicus erosus (Wollaston) | 30 | 30 | 0 | 0 | 0 |
| Crypturgus mediterraneus (Eichhoff) | 26 | 26 | 0 | 0 | 0 |
| Hylastes ater (Paykull) | 25 | 20 | 5 | 0 | 0 |
| Hylastes angustatus (Herbest) | 23 | 23 | 0 | 0 | 0 |
| Xyleborinus saxeseni Ratzenburg | 22 | 1 | 21 | 0 | 0 |
| Tomicus piniperda | 19 | 0 | 0 | 0 | 19 |
| Hylurgops palliatus (Gyllenhal) | 9 | 9 | 0 | 0 | 0 |
| Xyleborus dispar F. | 4 | 0 | 4 | 0 | 0 |
| Hylastes linearis Erichson | 1 | 1 | 0 | 0 | 0 |
| Total | 458 | 309 | 32 | 97 | 20 |
| Species | Shoots | Logs | Accession Number | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tomicus piniperda | Ips sexdentatus | Hylastes attenuatus | Orthotomicus erosus | Hylastes ater | Hylastes angustatus | Hylurgops palliatus | ||||||||
| Xylem | Phloem | Xylem | Phloem | Xylem | Phloem | Xylem | Phloem | Xylem | Phloem | Xylem | Phloem | |||
| Diplodia sapinea | 88.4 | 62.2 | 74.1 | 54.8 | 67.1 | 100.0 | - | 45.0 | 50.0 | 84.2 | 89.4 | 60.0 | 100.0 | KP900724 |
| Pestalotiopsis sp. | 24.5 | 2.5 | 5.4 | 4.11 | 2.7 | - | - | 10.0 | 5.0 | 21.05 | - | - | - | KP900723 |
| Mucor sp. | 3.8 | 0.0 | 1.8 | 15.1 | 6.9 | - | - | 50.0 | 15.0 | 5.26 | 10.5 | - | 16.6 | KP900722 |
| Trichoderma spp. | 9.1 | 19.3 | 21.4 | 11.0 | 15.0 | - | - | 15.0 | 10.0 | 10.53 | 15.7 | - | - | KP900738 |
| Fusarium spp. | 21.05 | 2.5 | 9.8 | 30.1 | 13.7 | 100.0 | - | 40.0 | 10.5 | 26.32 | 15.78 | 20.0 | 16.6 | - |
| Fusarium circinatum | 3.5 | - | 0.9 | - | 1.3 | - | - | - | - | 5.2 | 5.5 | - | - | - |
| Penicillium glabrum | 11.2 | 3.4 | - | 16.4 | 23.3 | - | - | 15.0 | - | 26.3 | 21.0 | - | 16.6 | KP900733 |
| Trichoderma harzianum | 11.9 | 46.2 | 52.7 | 20.5 | 20.5 | - | - | 5.0 | 30.0 | - | 10.5 | - | - | KP900736 |
| Trichoderma atroviride Bissett | 1.4 | - | - | - | - | - | - | - | - | - | - | - | - | KP900725 |
| Peniophora sp. | - | 1.7 | - | - | - | - | - | - | - | - | - | - | - | KP900735 |
| Gliocladium roseum Bainier | 12.2 | - | - | - | - | - | - | - | - | - | - | - | - | KP900726 |
| Botrytis cinerea Pers. | 6.3 | - | - | - | - | - | - | - | - | - | - | - | - | KP900730 |
| Epicoccum nigrum Link | 1.7 | - | - | - | - | - | - | - | - | - | - | - | - | KP900729 |
| M5 | 11.93 | 16.0 | 17.0 | 27.4 | 22.0 | - | - | 30.0 | 20.0 | 21.5 | 21.0 | 60.0 | 33.0 | - |
| M10 | 3.16 | 0.8 | 10.7 | 12.33 | 4.1 | - | 100.0 | 15.0 | 25.0 | 10.53 | 26.0 | 60.0 | - | - |
| M16 | 1.4 | 0.8 | - | - | 1.4 | - | - | - | - | - | - | - | - | - |
| M17 | 3.51 | 2.5 | - | 2.74 | 5.5 | - | - | - | 5.0 | 10.53 | 5.3 | - | - | - |
| M22 | 2.81 | 3.4 | 1.8 | - | - | - | - | - | - | - | - | - | - | - |
| M27 | 8.77 | 0.8 | - | - | - | - | - | - | - | - | - | - | - | - |
| Species | Pityophthorus pubescens | Ips sexdentatus | Hylastes attenuatus | Orthotomicus erosus | Crypturgus mediterraneus | Hylastes ater | Hylastes angustatus | Xyleborinus saxeseni | Hylurgops palliatus | Xileborus dispar | Accesion Number |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Candida fructus | 25.7 | 0.9 | 8.1 | 10 | - | 8 | 4.3 | 27.3 | 11.1 | 25 | KP900741 |
| Fusarium spp. | 20.6 | 34.48 | 32.5 | 56.6 | 12 | 64 | 39.1 | 31.8 | 55.5 | 25 | |
| Fusarium circinatum | 1.05 | 0.9 | 1.6 | - | - | - | - | - | - | - | |
| Gliocadium roseum | - | - | 2.3 | - | - | - | - | - | - | - | KP900740 |
| Neonectria radicícola | 5.1 | - | 2.3 | - | - | - | 4.3 | 22.3 | - | - | KP900737 |
| Penicillium sp. | 16.5 | - | - | - | - | - | 8.7 | 4.5 | - | - | KP900731 |
| Trichoderma atroviride | 1.03 | 0.9 | 3.48 | 3.3 | 8 | - | 4.3 | - | - | - | KP900728 |
| Mi1 | 25.7 | 0.9 | 8.1 | 10 | - | 8 | 4.3 | 27.3 | 11.1 | 25 | - |
| Mi2 | 1.03 | - | 11.6 | - | - | - | 8.7 | - | - | - | - |
| Mi5 | - | - | 4.6 | - | - | 8 | 4.3 | - | 11.1 | - | - |
| Mi6 | 1.03 | 1.7 | 1.16 | - | - | - | - | - | - | - | - |
| Mi9 | 1.03 | 1.7 | 6.96 | - | - | - | 8.7 | - | - | - | - |
| Mi14 | - | 1.7 | 2.3 | 6.6 | - | 4 | - | 4.5 | - | - | - |
| Mi16 | 2.06 | 0 | 5.81 | - | 4 | - | - | 4.5 | - | - | - |
| Isolate | Origin | Collected From | ITS Region | Morphology | TEF Region | Consensus Species | Accession Number TEF |
|---|---|---|---|---|---|---|---|
| 1 | Phloem | Logs I.s | Fusarium sp. | F. sporotrichioides | F. sporotrichioides | F. sporotrichioides | KR002044 |
| 2 | Phloem | Logs H. att | Fusarium sp. | F. beomiforme | - | F. beomiforme | - |
| 3 | Phloem | Logs I.s | F. oxysporum | F. oxysporum | F. oxysporum | F. oxysporum | KR002062 |
| 4 | Phloem | Logs H. ang | - | F. circinatum | F. circinatum | F. circinatum | KR002060 |
| 5 | Xylem | Logs H. att | Fusarium sp. | F. beomiforme | - | F. beomiforme | - |
| 6 | Xylem | Logs H.att | Fusarium sp. | F. oxysporum | F. oxysporum | F. oxysporum | KR002050 |
| 7 | Xylem | Logs H. a | Fusarium sp. | F. avenaceum | F. avenaceum | F. avenaceum | KR002057 |
| 8 | Xylem | Logs H. a | Fusarium sp. | F. avenaceum | F. avenaceum | F. avenaceum | KR002049 |
| 9 | Xylem | Logs Hy. p | Fusarium sp. | F. avenaceum | F. avenaceum | F. avenaceum | KR002058 |
| 10 | Shoot | Shoots T.p. | Fusarium sp. | F. avenaceum | F. avenaceum | F. avenaceum | KR002063 |
| 11 | Shoot | Shoots T.p. | Fusarium sp. | F. tricinctum | F. tricinctum | F. tricinctum | KR002064 |
| 12 | Shoot | Shoots T.p. | Fusarium sp. | F. cortaderiae | F. cortaderiae | F. cortaderiae | KR002048 |
| 13 | Shoot | Shoots T.p. | Fusarium sp. | F. avenaceum | F. avenaceum | F. avenaceum | KR002046 |
| 14 | Shoot | Shoots T.p. | Fusarium sp. | F. avenaceum | F. avenaceum | F. avenaceum | KR002051 |
| 15 | Shoot | Shoots T.p. | Fusarium sp. | F. tricinctum | F. tricinctum | F. tricinctum | KR002054 |
| 16 | Shoot | Shoots T.p. | Fusarium sp. | F. tricinctum | F. tricinctum | F. tricinctum | KR002047 |
| 17 | Shoot | Shoots T.p. | Fusarium sp. | F. sporotrichioides | F. sporotrichioides | F. sporotrichioides | - |
| 18 | Hylastes ater | Logs | F. lateritium | F. avenaceum | - | F. avenaceum | KR002059 |
| 19 | Hylastes attenuatus | Logs | Fusarium sp. | F. avenaceum | F. avenaceum | F. avenaceum | KR002052 |
| 20 | H. attenuatus | Logs | Fusarium sp. | F. avenaceum | F. avenaceum | F. avenaceum | - |
| 21 | H. attenuatus | Logs | Fusarium sp. | F. anthophilum | F. avenaceum | Fusarium sp. | KR002055 |
| 22 | H. attenuatus | Logs | Fusarium sp. | F. oxysporum | F. oxysporum | F. oxysporum | KR002065 |
| 23 | Ips sexdentatus | Logs | Fusarium sp. | F. avenaceum | F. avenaceum | F. avenaceum | KR002056 |
| 24 | I. sexdentatus | Logs | Fusarium sp. | F. avenaceum | F. avenaceum | F. avenaceum | KR002046 |
| 25 | I. sexdentatus | Logs | Fusarium sp. | F. avenaceum | F. avenaceum | F. avenaceum | - |
| 26 | I. sexdentatus | Logs | Fusarium sp. | F. tricinctum | - | F. tricinctum | - |
| 27 | I. sexdentatus | Logs | Fusarium sp. | F. oxysporum | F. oxysporum | F. oxysporum | KR002066 |
| 28 | Orthotomicus erosus | Logs | Fusarium sp. | F. oxysporum | F. oxysporum | F. oxysporum | KR002043 |
| 29 | Pityophthorus pubescens | Funnel | Fusarium sp. | F. sambucinum | F. sambucinum | F. sambucinum | KR002053 |
| 30 | Xyleborinus saxesni | Funnel | Fusarium sp. | F. konzum | - | F. konzum | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bezos, D.; Martínez-Álvarez, P.; Sanz-Ros, A.V.; Martín-García, J.; Fernandez, M.M.; Diez, J.J. Fungal Communities Associated with Bark Beetles in Pinus radiata Plantations in Northern Spain Affected by Pine Pitch Canker, with Special Focus on Fusarium Species. Forests 2018, 9, 698. https://doi.org/10.3390/f9110698
Bezos D, Martínez-Álvarez P, Sanz-Ros AV, Martín-García J, Fernandez MM, Diez JJ. Fungal Communities Associated with Bark Beetles in Pinus radiata Plantations in Northern Spain Affected by Pine Pitch Canker, with Special Focus on Fusarium Species. Forests. 2018; 9(11):698. https://doi.org/10.3390/f9110698
Chicago/Turabian StyleBezos, Diana, Pablo Martínez-Álvarez, Antonio V. Sanz-Ros, Jorge Martín-García, M. Mercedes Fernandez, and Julio J. Diez. 2018. "Fungal Communities Associated with Bark Beetles in Pinus radiata Plantations in Northern Spain Affected by Pine Pitch Canker, with Special Focus on Fusarium Species" Forests 9, no. 11: 698. https://doi.org/10.3390/f9110698
APA StyleBezos, D., Martínez-Álvarez, P., Sanz-Ros, A. V., Martín-García, J., Fernandez, M. M., & Diez, J. J. (2018). Fungal Communities Associated with Bark Beetles in Pinus radiata Plantations in Northern Spain Affected by Pine Pitch Canker, with Special Focus on Fusarium Species. Forests, 9(11), 698. https://doi.org/10.3390/f9110698