Refining Species Traits in a Dynamic Vegetation Model to Project the Impacts of Climate Change on Tropical Trees in Central Africa

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Studied Species
2.2. Climate Data and Future Scenarios
2.3. The CARAIB Dynamic Vegetation Model
- A constant atmospheric CO2 mixing ratio of 330 ppmv ([330] configuration) in the hypothesis that there will be no fertilization effect due to nutrient limitation. Maintaining CO2 concentrations constant in the model is indeed the only way to test this hypothesis since nutrient limitation in the model is only induced by the species C:N ratio which is a constant input;
- An increasing atmospheric CO2 mixing ratio ([var] configuration) according to the projections of RCP4.5 and RCP8.5 scenarios which respectively reach 538 ppmv and 936 ppmv in 2100. This, then, is consistent with the hypothesis that there will be a fertilizing effect and no nutrient limitation;
- An increasing atmospheric CO2 mixing ratio equal to those of the [var] configuration but accompanied by a downregulation of photosynthetic activity ([down] configuration) in an attempt to take the influence of decreasing nitrogen availability on the CO2 fertilizing effect into account. Whereas the maximum carboxylation rate (Vcmax) for the first two is calculated as a function of foliage C/N and SLA [74], the third CO2 configuration consists of a calibration of the photosynthesis based on empirical values of Vcmax and maximum rate of electron transport (Jmax) measured in free-air CO2 enrichment (FACE) experiments following the meta-analysis of Ainsworth and Rogers [90]. These experiments were conducted mostly in temperate ecosystems. Their results are thus not directly transferable to the tropical species studied here. Nonetheless, we used them to construct an intermediate scenario of CO2 fertilization, as a sensitivity test, since such experiments have essentially not been conducted up to now in the tropics [91].
3. Results
3.1. CARAIB Simulations for the Present
3.1.1. Refinement of Morpho-Physiological Traits
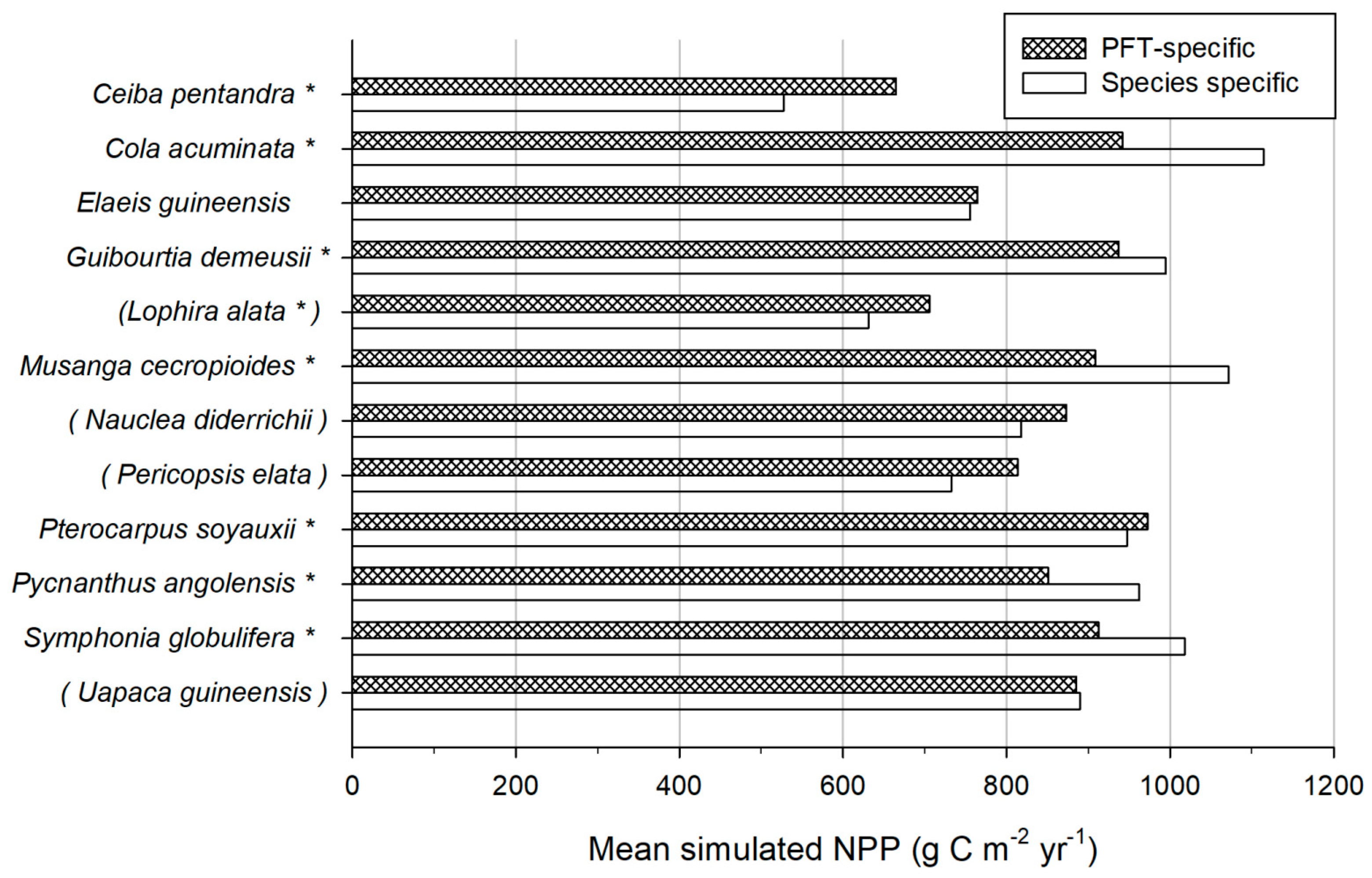
3.1.2. Modelling Validation
3.2. CARAIB Simulations for the Future
3.2.1. Climate Scenarios
3.2.2. Changes in Soil Water Content
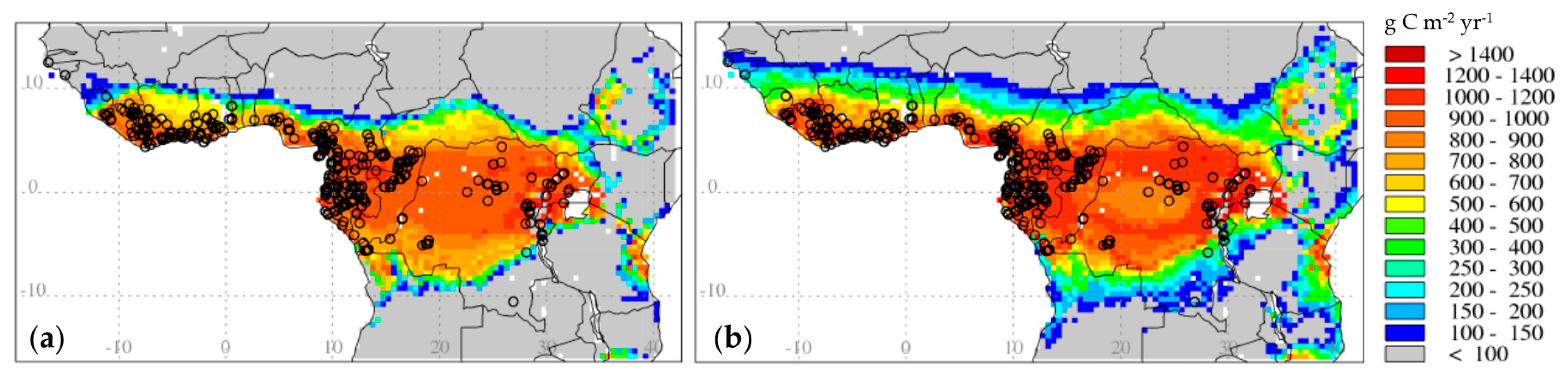
3.2.3. Changes in Productivity and Distribution
3.2.4. Changes in Biomass
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brink, A.B.; Eva, H.D. Monitoring 25 years of land cover change dynamics in Africa: A sample based remote sensing approach. Appl. Geogr. 2009, 29, 501–512. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of the United Nations (FAO). State of the World’s Forests; FAO: Rome, Italy, 2012; p. 56. ISBN 978-9251072929. [Google Scholar]
- O’Farrell, P.J.; Anderson, P.M.L.; Le Maitre, D.C.; Holmes, P.M. Insights and opportunities offered by a rapid ecosystem service assessment in promoting a conservation agenda in an urban biodiversity hotspot. Ecol. Soc. 2012, 17, 27. [Google Scholar] [CrossRef]
- Grace, J.; San José, J.; Meir, P.; Miranda, H.S.; Montes, R.A. Productivity and carbon fluxes of tropical savannas. J. Biogeogr. 2006, 33, 387–400. [Google Scholar] [CrossRef]
- Norris, K.; Asase, A.; Collen, B.; Gockowksi, J.; Mason, J.; Phalan, B.; Wade, A. Biodiversity in a forest-agriculture mosaic—The changing face of West African rainforests. Biol. Conserv. 2010, 143, 2341–2350. [Google Scholar] [CrossRef]
- Boko, M.; Niang, I.; Nyong, A.; Vogel, C.; Githeko, A.; Medany, M.; Osman-Elasha, B.; Tabo, R.; Yanda, P. Africa. In Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Parry, M.L., Canziani, O.F., Palutikof, J.P., van der Linden, P.J., Hanson, C.E., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 433–467. [Google Scholar]
- Brodie, J.; Post, E.; Laurance, W.F. Climate change and tropical biodiversity: A new focus. Trends Ecol. Evol. 2012, 27, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Iloweka, E.M. The Deforestation of Rural Areas in the Lower Congo Province. Environ. Monit. Assess. 2004, 99, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Ahrends, A.; Burgess, N.D.; Milledge, S.A.H.; Bulling, M.T.; Fisher, B.; Smart, J.C.R.; Clarke, G.P.; Mhoro, B.E.; Lewis, S.L. Predictable waves of sequential forest degradation and biodiversity loss spreading from an African city. Proc. Natl. Acad. Sci. USA 2010, 107, 14556–14561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keenan, R.J.; Reams, G.A.; Achard, F.; de Freitas, J.V.; Grainger, A.; Lindquist, E. Dynamics of global forest area: Results from the FAO Global Forest Resources Assessment 2015. For. Ecol. Manag. 2015, 352, 9–20. [Google Scholar] [CrossRef]
- Yadav, P.K.; Sarma, K.; Kumar, R. A framework for assessing the impact of urbanization and population pressure on Garo hills landscape of north-east India. Int. J. Conserv. Sci. 2013, 4, 213–222. [Google Scholar]
- Kukkonen, M.; Käyhkö, N. Spatio-temporal analysis of forest changes in contrasting land use regimes of Zanzibar, Tanzania. Appl. Geogr. 2014, 55, 193–202. [Google Scholar] [CrossRef]
- Thilagavathi, N.; Subramani, T.; Suresh, M. Land use/land cover change detection analysis in Salem Chalk Hills, South India using remote sensing and GIS. Disaster Adv. 2015, 8, 44–52. [Google Scholar]
- Pessoa, M.S.; Rocha-Santos, L.; Talora, D.C.; Faria, D.; Mariano-Neto, E.; Hambuckers, A.; Cazetta, E. Fruit biomass availability along a forest cover gradient. Biotropica 2016, 49, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Pütz, S.; Groeneveld, J.; Alves, L.F.; Metzger, J.P.; Huth, A. Fragmentation drives tropical forest fragments to early successional states: A modelling study for Brazilian Atlantic forests. Ecol. Model. 2011, 222, 1986–1997. [Google Scholar] [CrossRef]
- Tabarelli, M.; Peres, C.A.; Melo, F.P.L. The ‘few winners and many losers’ paradigm revisited: Emerging prospects for tropical forest biodiversity. Biol. Conserv. 2012, 155, 136–140. [Google Scholar] [CrossRef]
- Niang, I.; Ruppel, O.C.; Abdrabo, M.A.; Essel, A.; Lennard, C.; Padgham, J.; Urquhart, P. Africa. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Barros, V.R., Field, C.B., Dokken, D.J., Mastrandrea, M.D., Mach, K.J., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 1199–1265. [Google Scholar]
- Gonzalez, P.; Neilson, R.P.; Lenihan, J.M.; Drapek, R.J. Global patterns in the vulnerability of ecosystems to vegetation shifts due to climate change. Glob. Ecol. Biogeogr. 2010, 19, 755–768. [Google Scholar] [CrossRef]
- Lewis, S.L.; Sonké, B.; Sunderland, T.; Begne, S.K.; Lopez-Gonzalez, G.; van der Heijden, G.M.F.; Phillips, O.L.; Affum-Baffoe, K.; Baker, T.R.; Banin, L.; et al. Above-ground biomass and structure of 260 African tropical forests. Philos. Trans. R. Soc. B 2013, 368, 20120295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhi, Y.; Adu-Bredu, S.; Asare, R.A.; Lewis, S.L.; Mayaux, P. African rainforests: Past, present and future. Philos. Trans. R. Soc. B 2013, 368, 20120312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunting, E.L.; Fullman, T.; Kiker, G.; Southworth, J. Utilization of the SAVANNA model to analyze future patterns of vegetation cover in Kruger National Park under changing climate. Ecol. Model. 2016, 342, 147–160. [Google Scholar] [CrossRef]
- Hély, C.; Bremond, L.; Alleaume, S.; Smith, B.; Sykes, M.T.; Guiot, J. Sensitivity of African biomes to changes in the precipitation regime. Glob. Ecol. Biogeogr. 2006, 15, 258–270. [Google Scholar] [CrossRef]
- Williams, C.A.; Hanan, N.P.; Neff, J.C.; Scholes, R.J.; Berry, J.A.; Denning, A.S.; Baker, D.F. Africa and the global carbon cycle. Carbon Balance Manag. 2007, 2, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, S.L.; Lopez-Gonzalez, G.; Sonké, B.; Affum-Baffoe, K.; Baker, T.R.; Ojo, L.O.; Phillips, O.L.; Reitsma, J.M.; White, L.; Comiskey, J.A.; et al. Increasing carbon storage in intact African tropical forests. Nature 2009, 457, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Chambers, L.E.; Altwegg, R.; Barbraud, C.; Barnard, P.; Beaumont, L.J.; Crawford, R.J.M.; Durant, J.M.; Hughes, L.; Keatley, M.R.; Low, M.; et al. Phenological Changes in the Southern Hemisphere. PLoS ONE 2013, 8, e75514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grace, J.; Mitchard, E.; Gloor, E. Perturbations in the carbon budget of the tropics. Glob. Chang. Biol. 2014, 20, 3238–3255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cusack, D.F.; Karpman, J.; Ashdown, D.; Cao, Q.; Ciochina, M.; Halterman, S.; Lydon, S.; Neupane, A. Global change effects on humid tropical forests: Evidence for biogeochemical and biodiversity shifts at an ecosystem scale. Rev. Geophys. 2016, 54, 523–610. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, F.; Franssen, W.; Jans, W.; Beyenne, T.; Kruijt, B.; Supit, I. Climate change impacts on the Congo Basin region. In Climate Change Scenarios for the Congo Basin; Report No. 11; Haensler, A., Jacob, D., Kabat, P., Ludwig, F., Eds.; Climate Service Centre: Hamburg, Germany, 2013. [Google Scholar]
- Moncrieff, G.R.; Scheiter, S.; Slingsby, J.A.; Higgins, S.I. Understanding global change impacts on South African biomes using Dynamic Vegetation Models. S. Afr. J. Bot. 2015, 101, 16–23. [Google Scholar] [CrossRef]
- Erfanian, A.; Wang, G.; Yu, M.; Anyah, R. Multimodel ensemble simulations of present and future climates over West Africa: Impacts of vegetation dynamics. J. Adv. Model. Earth Syst. 2016, 8, 1411–1431. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Yu, M.; Pal, J.S.; Mei, R.; Bonan, G.B.; Levis, S.; Thornton, P.E. On the development of a coupled regional climate–vegetation model RCM–CLM–CN–DV and its validation in Tropical Africa. Clim. Dyn. 2016, 46, 515–539. [Google Scholar] [CrossRef]
- Thuiller, W.; Midgley, G.F.; Hughes, G.O.; Bomhard, B.; Drew, G.; Rutherford, M.C.; Woodward, F.I. Endemic species and ecosystem sensitivity to climate change in Namibia. Glob. Chang. Biol. 2006, 12, 759–776. [Google Scholar] [CrossRef]
- Mbatudde, M.; Mwanjololo, M.; Kakudidi, E.K.; Dalitz, H. Modelling the potential distribution of endangered Prunus africana (Hook.f.) Kalkm. in East Africa. Afr. J. Ecol. 2012, 50, 393–403. [Google Scholar] [CrossRef]
- Idohou, R.; Assogbadjo, A.E.; Kakaï, R.G.; Peterson, A.T. Spatio-temporal dynamic of suitable areas for species conservation in West Africa: Eight economically important wild palms under present and future climates. Agrofor. Syst. 2017, 91, 527–540. [Google Scholar] [CrossRef]
- Sarmento Cabral, J.; Jeltsch, F.; Thuiller, W.; Higgins, S.; Midgley, G.F.; Rebelo, A.G.; Rouget, M.; Schurr, F.M. Impacts of past habitat loss and future climate change on the range dynamics of South African Proteaceae. Divers. Distrib. 2013, 19, 363–376. [Google Scholar] [CrossRef]
- Frenette-Dussault, C.; Shipley, B.; Meziane, D.; Hingrat, Y. Trait-based climate change predictions of plant community structure in arid steppes. J. Ecol. 2013, 101, 484–492. [Google Scholar] [CrossRef]
- Tabet, S.; Belhemra, M.; François, L.; Arar, A. Evaluation by prediction of the natural range shrinkage of Quercus ilex L. in eastern Algeria. Forestist 2018, 68, 7–15. [Google Scholar] [CrossRef]
- Ouled Belgacem, A.; Louhaichi, M. The vulnerability of native rangeland plant species to global climate change in the West Asia and North African regions. Clim. Chang. 2013, 119, 451–463. [Google Scholar] [CrossRef] [Green Version]
- Hartig, F.; Dyke, J.; Hickler, T.; Higgins, S.I.; O’Hara, R.B.; Scheiter, S.; Huth, A. Connecting dynamic vegetation models to data—An inverse perspective. J. Biogeogr. 2012, 39, 2240–2252. [Google Scholar] [CrossRef]
- Morin, X.; Thuiller, W. Comparing niche- and process-based models to reduce prediction uncertainty in species range shifts under climate change. Ecology 2009, 90, 1301–1313. [Google Scholar] [CrossRef] [PubMed]
- Delire, C.; Ngomanda, A.; Jolly, D. Possible impacts of 21st century climate on vegetation in Central and West Africa. Glob. Planet. Chang. 2008, 64, 3–15. [Google Scholar] [CrossRef]
- Zelazowski, P.; Malhi, Y.; Huntingford, C.; Sitch, S.; Fisher, J.B. Changes in the potential distribution of humid tropical forests on a warmer planet. Philos. Trans. R. Soc. Lond. A 2011, 369, 137–160. [Google Scholar] [CrossRef] [Green Version]
- Budde, K.B.; Gonzalez-Martinez, S.C.; Hardy, O.J.; Heuertz, M. The ancient tropical rainforest tree Symphonia globulifera L. f. (Clusiaceae) was not restricted to postulated Pleistocene refugia in Atlantic Equatorial Africa. Heredity 2013, 111, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Feeley, K.; Silman, M. The data void in modelling current and future distributions of tropical species. Glob. Chang. Biol. 2011, 17, 626–630. [Google Scholar] [CrossRef]
- François, L.; Delire, C.; Warnant, P.; Munhoven, G. Modelling the glacial-interglacial changes in the continental biosphere. Glob. Planet. Chang. 1998, 16–17, 37–52. [Google Scholar] [CrossRef]
- François, L.; Ghislain, M.; Otto, D.; Micheels, A. Late Miocene vegetation reconstruction with the CARAIB model. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 238, 302–320. [Google Scholar] [CrossRef] [Green Version]
- Dury, M.; Hambuckers, A.; Warnant, P.; Henrot, A.; Favre, E.; Ouberdous, M.; François, L. Responses of European forest ecosystems to 21st century climate: Assessing changes in interannual variability and fire intensity. iForest 2011, 4, 82–99. [Google Scholar] [CrossRef]
- Dury, M. Modelling the Dynamics of European Ecosystems from the Early Holocene to the End of the 21st Century with the CARAIB Dynamic Vegetation Model. Ph.D. Thesis, Université de Liège, Liège, Belgium, 2015. [Google Scholar]
- Raghunathan, N.; François, L.; Huynen, M.-C.; Oliveira, L.C.; Hambuckers, A. Modelling the distribution of key tree species used by lion tamarins in the Brazilian Atlantic forest under a scenario of future climate change. Reg. Environ. Chang. 2015, 15, 683–693. [Google Scholar] [CrossRef]
- Raghunathan, N.; François, L.; Dury, M.; Hambuckers, A. Contrasting climate risks predicted by dynamic vegetation and ecological niche-based models applied to tree species in the Brazilian Atlantic Forest. Reg. Environ. Chang. 2018. [Google Scholar] [CrossRef]
- Minet, J.; Laloy, E.; Tychon, B.; François, L. Bayesian inversions of a dynamic vegetation model at four European grassland sites. Biogeosciences 2015, 12, 2809–2829. [Google Scholar] [CrossRef] [Green Version]
- Taylor, K.E.; Stouffer, R.J.; Meehl, G.A. An overview of CMIP5 and the experiment design. Bull. Am. Meteorol. Soc. 2012, 93, 485–498. [Google Scholar] [CrossRef]
- Moss, R.H.; Edmonds, J.A.; Hibbard, K.A.; Manning, M.R.; Rose, S.K.; van Vuuren, D.P.; Carter, T.R.; Emori, S.; Kainuma, M.; Kram, T.; et al. The next generation of scenarios for climate change research and assessment. Nature 2010, 463, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Hambuckers, A.; Huynen, M.-C.; Trolliet, F.; Jamar-Beudels, R.; Lafontaine, R.-M.; Robert, H.; Van Damme, P.; Baert, A.; Dendoncker, N.; Fontaine, C.; et al. BIOSERF: Sustainability of Tropical Forest Biodiversity and Services under Climate and Human Pressure; Final Report; Research Programme Science for a Sustainable Development; Belgian Science Policy: Brussels, Belgium, 2015.
- Dauby, G.; Zaiss, R.; Blach-Overgaard, A.; Catarino, L.; Damen, T.; Deblauwe, V.; Dessein, S.; Dransfield, J.; Droissart, V.; Duarte, M.C.; et al. RAINBIO: A mega-database of tropical African vascular plants distributions. PhytoKeys 2016, 74, 1–18. [Google Scholar] [CrossRef]
- Fayolle, A.; Swaine, M.D.; Bastin, J.-F.; Bourland, N.; Comiskey, J.A.; Dauby, G.; Doucet, J.-L.; Gillet, J.-F.; Gourlet-Fleury, S.; Hardy, O.J.; et al. Patterns of tree species composition across tropical African forests. J. Biogeogr. 2014, 41, 2320–2331. [Google Scholar] [CrossRef]
- Kattge, J.; Díaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Bönisch, G.; Garnier, E.; Westoby, M.; Reich, P.B.; Wright, I.J.; et al. TRY—A global database of plant traits. Glob. Chang. Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef] [Green Version]
- Kearsley, E.; de Haulleville, T.; Hufkens, T.; Kidimbu, K.; Toirambe, B.; Baert, G.; Huygens, D.; Kebede, Y.; Defourny, P.; Bogaert, J.; et al. Conventional tree height-diameter relationships significantly overestimate aboveground carbon stocks in the Central Congo Basin. Nat. Commun. 2013, 4, 2269. [Google Scholar] [CrossRef] [PubMed]
- Kearsley, E.; Verbeeck, H.; Hufkens, K.; Van de Perre, F.; Doetterl, S.; Baert, G.; Beeckman, H.; Boeckx, P.; Huygens, D. Functional community structure of African monodominant Gilbertiodendron dewevrei forest influenced by local environmental filtering. Ecol. Evol. 2017, 7, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P. The Economics of Ecosystems and Biodiversity: Ecological and Economic Foundations, 1st ed.; Earthscan: London, UK; Washington, DC, USA, 2010; p. 456. ISBN 9781136538803. [Google Scholar]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations—The CRU TS3.10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- Voldoire, A.; Sanchez-Gomez, E.; Salas y Mélia, D.; Decharme, B.; Cassou, C.; Sénési, S.; Valcke, S.; Beau, I.; Alias, A.; Chevallier, M.; et al. The CNRM-CM5.1 global climate model: Description and basic evaluation. Clim. Dyn. 2013, 40, 2091–2121. [Google Scholar] [CrossRef] [Green Version]
- Giorgetta, M.A.; Jungclaus, J.; Reick, C.H.; Legutke, S.; Bader, J.; Böttinger, M.; Brovkin, V.; Crueger, T.; Esch, M.; Fieg, K.; et al. Climate and carbon cycle changes from 1850 to 2100 in MPI-ESM simulations for the Coupled Model Intercomparison Project phase 5. J. Adv. Model. Earth Syst. 2013, 5, 572–597. [Google Scholar] [CrossRef] [Green Version]
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; et al. The community climate system model version 4. J. Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- Arora, V.K.; Scinocca, J.F.; Boer, G.J.; Christian, J.R.; Denman, K.L.; Flato, G.M.; Kharin, V.V.; Lee, W.G.; Merryfield, W.J. Carbon emission limits required to satisfy future representative concentration pathways of greenhouse gases. Geophys. Res. Lett. 2011, 38. [Google Scholar] [CrossRef] [Green Version]
- Gordon, H.B.; O’Farrell, S.; Collier, M.; Dix, M.; Rotstayn, L.; Kowalczyk, E.; Hirst, T.; Watterson, I. The CSIRO Mk3.5 Climate Model; CAWCR Technical Report No. 21; The Centre for Australian Weather and Climate Research: Melbourne, Australia, 2010; p. 74. ISBN 978-1-921605-666.
- Volodin, E.M.; Dianskii, N.A.; Gusev, A.V. Simulating present-day climate with the INMCM4.0 coupled model of the atmospheric and oceanic general circulations. Izv. Atmos. Ocean. Phys. 2010, 46, 414–431. [Google Scholar] [CrossRef]
- Warnant, P.; François, L.; Strivay, D.; Gérard, J.-C. CARAIB—A global model of terrestrial biological productivity. Glob. Biogeochem. Cycles 1994, 8, 255–270. [Google Scholar] [CrossRef]
- Gérard, J.-C.; Nemry, B.; François, L.; Warnant, P. The interannual change of atmospheric CO2: Contribution of subtropical ecosystems? Geophys. Res. Lett. 1999, 26, 243–246. [Google Scholar] [CrossRef]
- Henrot, A.-J.; Utescher, T.; Erdei, B.; Dury, M.; Hamon, N.; Ramstein, G.; Krapp, M.; Herold, N.; Goldner, A.; Favre, E.; et al. Middle Miocene climate and vegetation models and their validation with proxy data. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 467, 95–119. [Google Scholar] [CrossRef]
- Otto, D.; Rasse, D.; Kaplan, J.; Warnant, P.; François, L. Biospheric carbon stocks reconstructed at the Last Glacial Maximum: Comparison between general circulation models using prescribed and computed sea surface temperatures. Glob. Planet. Chang. 2002, 33, 117–138. [Google Scholar] [CrossRef]
- François, L.; Utescher, T.; Favre, E.; Henrot, A.; Warnant, P.; Micheels, A.; Erdei, B.; Suc, J.-P.; Cheddadi, R.; Mosbrugger, V. Modelling Late Miocene vegetation in Europe: Results of the CARAIB model and comparison with palaeovegetation data. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 304, 359–378. [Google Scholar] [CrossRef]
- Hubert, B.; François, L.; Warnant, P.; Strivay, D. Stochastic generation of meteorological variables and effects on global models of water and carbon cycles in vegetation and soils. J. Hydrol. 1998, 212–213, 317–334. [Google Scholar] [CrossRef]
- Walker, A.P.; Beckerman, A.P.; Gu, L.; Kattge, J.; Cernusak, L.A.; Domingues, T.F.; Scales, J.C.; Wohlfahrt, G.; Wullschleger, S.D.; Woodward, F.I. The relationship of leaf photosynthetic traits—Vcmax and Jmax—To leaf nitrogen, leaf phosphorus, and specific leaf area: A meta-analysis and modeling study. Ecol. Evol. 2014, 4, 3218–3235. [Google Scholar] [CrossRef] [PubMed]
- Laurent, J.-M.; François, L.; Bar-Hen, A.; Bel, L.; Cheddadi, R. European Bioclimatic Affinity Groups: Data-model comparisons. Glob. Planet. Chang. 2008, 61, 28–40. [Google Scholar] [CrossRef]
- Schenk, H.J.; Jackson, R.B. The global biogeography of roots. Ecol. Monogr. 2002, 72, 311–328. [Google Scholar] [CrossRef]
- McGroddy, M.E.; Daufresne, T.; Hedin, L.O. Scaling of C:N:P Stoichiometry in Forests Worldwide: Implications of Terrestrial Redfield-Type Ratios. Ecology 2004, 85, 2390–2401. [Google Scholar] [CrossRef]
- Warnant, P. Modélisation du Cycle du Carbone Dans la Biosphère Continentale à L’échelle Globale. Ph.D. Thesis, Université de Liège, Liège, Belgium, 1999. [Google Scholar]
- Shipley, B. Trade-offs between net assimilation rate and specific leaf area in determining relative growth rate: Relationship with daily irradiance. Funct. Ecol. 2002, 16, 682–689. [Google Scholar] [CrossRef]
- Meir, P.; Levy, P.E. Photosynthetic parameters from two contrasting woody vegetation types in West Africa. Plant Ecol. 2007, 192, 277–287. [Google Scholar] [CrossRef]
- Wright, I.J.; Ackerly, D.D.; Bongers, F.; Harms, K.E.; Ibarra-Manriquez, G.; MartinezRamos, M.; Mazer, S.J.; Muller-Landau, H.C.; Paz, H.; Pitman, N.C.A.; et al. Relationships among ecologically important dimensions of plant trait variation in seven Neotropical forests. Ann. Bot. 2007, 99, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Fyllas, N.M.; Patino, S.; Baker, T.R.; Bielefeld Nardoto, G.; Martinelli, L.A.; Quesada, C.A.; Paiva, R.; Schwarz, M.; Horna, V.; Mercado, L.M.; et al. Basin-wide variations in foliar properties of Amazonian forest: Phylogeny, soils and climate. Biogeosciences 2009, 6, 2677–2708. [Google Scholar] [CrossRef] [Green Version]
- Green, W. USDA PLANTS Compilation, version 1, 09-02-02. In NRCS: The PLANTS Database; National Plant Data Center: Baton Rouge, LA, USA, 2009. [Google Scholar]
- Reich, P.B.; Wright, I.J.; Lusk, C.H. Predicting leaf physiology from simple plant and climate attributes: A global GLOPNET analysis. Ecol. Appl. 2007, 17, 1982–1988. [Google Scholar] [CrossRef] [PubMed]
- Onoda, Y.; Westoby, M.; Adler, P.B.; Choong, A.M.F.; Clissold, F.J.; Cornelissen, J.H.C.; Diaz, S.; Dominy, N.J.; Elgart, A.; Enrico, L.; et al. Global patterns of leaf mechanical properties. Ecol. Lett. 2011, 14, 301–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kattge, J.; Knorr, W.; Raddatz, T.; Wirth, C. Quantifying photosynthetic capacity and its relationship to leaf nitrogen content for global-scale terrestrial biosphere models. Glob. Chang. Biol. 2009, 15, 976–991. [Google Scholar] [CrossRef]
- Baraloto, C.; Paine, C.E.T.; Poorter, L.; Beauchene, J.; Bonal, D.; Domenach, A.-M.; Herault, B.; Patino, S.; Roggy, J.-C.; Chave, J. Decoupled leaf and stem economics in rainforest trees. Ecol. Lett. 2010, 13, 1338–1347. [Google Scholar] [CrossRef] [PubMed]
- Domingues, T.F.; Meir, P.; Feldpausch, T.R.; Saiz, G.; Veenendaal, E.M.; Schrodt, F.; Bird, M.; Djagbletey, G.; Hien, F.; Compaore, H.; et al. Co-limitation of photosynthetic capacity by nitrogen and phosphorus in West Africa woodlands. Plant Cell Environ. 2010, 33, 959–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial gross carbon dioxide uptake: Global distribution and covariation with climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.G.; Scullion, J.; Ostle, N.; Levy, P.E.; Gwynn-Jones, D. Completing the FACE of elevated CO2 research. Environ. Int. 2014, 73, 252–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhi, Y. The productivity, metabolism and carbon cycle of tropical forest vegetation. J. Ecol. 2012, 100, 65–75. [Google Scholar] [CrossRef]
- Whittaker, R.H. Communities and Ecosystems, 2nd ed.; Collier-Macmillan: New York, NY, USA, 1975; p. 352. ISBN 978-0024273901. [Google Scholar]
- Martinez-Yrizar, A.; Maass, J.M.; Perez-Jimenez, L.A.; Sarukhan, J. Net primary productivity of a tropical deciduous forest ecosystem in western Mexico. J. Trop. Ecol. 1996, 12, 169–175. [Google Scholar] [CrossRef]
- Poorter, L.; Castilho, C.V.; Schietti, J.; Oliveira, R.S.; Costa, F.R.C. Can traits predict individual growth performance? A test in a hyperdiverse tropical forest. New Phytol. 2018, 219, 109–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singer, A.; Johst, K.; Banitz, T.; Fowler, M.S.; Groeneveld, J.; Gutierrez, A.G.; Hartig, F.; Krug, R.M.; Liess, M.; Matlack, G.; et al. Community dynamics under environmental change: How can next generation mechanistic models improve projections of species distributions? Ecol. Model. 2016, 326, 63–74. [Google Scholar] [CrossRef]
- Rogers, A.; Medlyn, B.E.; Dukes, J.S. Improving representation of photosynthesis in Earth System Models. New Phytol. 2014, 204, 12–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.Y.; van Dijk, A.; de Jeu, R.; Canadell, J.G.; McCabe, M.; Evans, J.; Wang, G. Recent reversal in loss of global terrestrial biomass. Nat. Clim. Chang. 2015, 5, 470–474. [Google Scholar] [CrossRef]
- Smith, N.G.; Dukes, J.S. Plant respiration and photosynthesis in global-scale models: Incorporating acclimation to temperature and CO2. Glob. Chang. Biol. 2013, 19, 45–63. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Schurgers, G.; Rummukainen, M.; Smith, B.; Samuelsson, P.; Jansson, C.; Siltberg, J.; May, W. Vegetation–Climate feedbacks modulate rainfall patterns in Africa under future climate change. Earth Syst. Dyn. 2016, 7, 627–647. [Google Scholar] [CrossRef]
- Haxeltine, A.; Prentice, I.C. A general model for the light-use efficiency of primary production. Funct. Ecol. 1996, 10, 551–561. [Google Scholar] [CrossRef]
- Bush, M.B.; Flenley, J.R.; Gosling, W.D. Tropical Rainforest Responses to Climate Change, 2nd ed.; Springer: Dordrecht, The Netherlands, 2011; p. 454. ISBN 978-3-642-05383-2. [Google Scholar]
- James, R.; Washington, R.; Rowell, D.P. Implications of global warming for the climate of African rainforests. Philos. Trans. R. Soc. B 2013, 368, 20120298. [Google Scholar] [CrossRef] [PubMed]
- Rammig, A.; Jupp, T.; Thonicke, K.; Tietjen, B.; Heinke, J.; Ostberg, S.; Lucht, W.; Cramer, W.; Cox, P. Estimating the risk of Amazonian forest dieback. New Phytol. 2010, 187, 694–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huntingford, C.; Zelazowski, P.; Galbraith, D.; Mercado, L.M.; Sitch, S.; Fisher, R.; Lomas, M.; Walker, A.P.; Jones, C.D.; Booth, B.B.B.; et al. Simulated resilience of tropical rainforests to CO2-induced climate change. Nat. Geosci. 2013, 6, 268–273. [Google Scholar] [CrossRef]
- Gopalakrishnan, R.; Jayaraman, M.; Swarnim, S.; Chaturvedi, R.; Bala, G.; Ravindranath, N. Impact of climate change at species level: A case study of teak in India. Mitig. Adapt. Strateg. Glob. Chang. 2011, 16, 199–209. [Google Scholar] [CrossRef]
- Lloyd, J.; Farquhar, G. Effects of rising temperatures and [CO2] on the physiology of tropical forest trees. Philos. Trans. R. Soc. Lond. B 2008, 363, 1811–1817. [Google Scholar] [CrossRef] [PubMed]
- Scheiter, S.; Higgins, S.I. Impacts of climate change on the vegetation of Africa: An adaptive dynamic vegetation modelling approach. Glob. Chang. Biol. 2009, 15, 2224–2246. [Google Scholar] [CrossRef]
- Malhi, Y.; Aragão, L.E.O.C.; Galbraith, D.; Huntingford, C.; Fisher, R.; Zelazowski, P.; Sitch, S.; McSweeney, C.; Meir, P. Exploring the likelihood and mechanism of a climate-change-induced dieback of the Amazon rainforest. Proc. Natl. Acad. Sci. USA 2009, 106, 20610–20615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Wang, G.; Parr, D.; Ahmed, K.F. Future changes of the terrestrial ecosystem based on a dynamic vegetation model driven with RCP8.5 climate projections from 19 GCMs. Clim. Chang. 2014, 127, 257–271. [Google Scholar] [CrossRef]
- Bastin, J.F. Estimating the Aboveground Biomass of Central African Tropical Forests at the Tree, the Canopy and the Region Level. Ph.D. Thesis, Université de Liège, Liège, Belgium, 2014. [Google Scholar]
- Santiago, L.S.; Bonal, D.; De Guzman, M.E.; Ávila-Lovera, E. Drought Survival Strategies of Tropical Trees. In Tropical Tree Physiology; Goldstein, G., Santiago, L., Eds.; Springer: Cham, Switzerland, 2016; Volume 6, pp. 243–258. ISBN 978-90-8585-534-7. [Google Scholar]
- Scheiter, S.; Savadogo, P. Ecosystem management can mitigate vegetation shifts induced by climate change in West Africa. Ecol. Model. 2016, 332, 19–27. [Google Scholar] [CrossRef]
- Davies-Barnard, T.; Valdes, P.J.; Singarayer, J.S.; Wiltshire, A.J.; Jones, C.D. Quantifying the relative importance of land cover change from climate and land use in the representative concentration pathways. Glob. Biogeochem. Cycles 2015, 29, 842–853. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, P. Introduction: The Social Sciences in a Cross-Disciplinary Age. J. Hist. Behav. Sci. 2015, 51, 1–9. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.L. Climate change and social transformations: Is it time for a quantum leap? Wiley Interdiscip. Rev. Clim. Chang. 2016, 7, 618–626. [Google Scholar] [CrossRef]
- Trolliet, F.; Forget, P.-M.; Huynen, M.-C.; Hambuckers, A. Forest cover, hunting pressure, and fruit availability influence seed dispersal in a forest-savanna mosaic in the Congo Basin. Biotropica 2017, 49, 337–345. [Google Scholar] [CrossRef]
- García, C.; Klein, A.K.; Jordano, P. Dispersal processes driving plant movement: Challenges for understanding and predicting range shifts in a changing world. J. Ecol. 2017, 105, 1–5. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (Family) | Ecosystem Services | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| food | fodder | med | rit/soc | envi | poison | constr | mat | fuel | ||
| 1 | Ceiba pentandra (Malvaceae) | x | x | x | x | x | x | |||
| 2 | Cola acuminata (Malvaceae) | x | x | x | x | |||||
| 3 | Elaeis guineensis (Arecaceae) | x | x | x | x | x | x | |||
| 4 | Guibourtia demeusei (Fabaceae) | x | x | x | x | x | x | x | x | |
| 5 | Lophira alata (Ochnaceae) | x | x | x | x | x | ||||
| 6 | Musanga cecropioides (Urticaceae) | x | x | x | x | x | ||||
| 7 | Nauclea diderrichii (Rubiaceae) | x | x | x | x | x | x | x | ||
| 8 | Pericopsis elata (Fabaceae) | x | x | x | ||||||
| 9 | Pterocarpus soyauxii (Fabaceae) | x | x | x | x | x | ||||
| 10 | Pycnanthus angolensis (Myristicaceae) | x | x | x | x | x | x | |||
| 11 | Symphonia globulifera (Clusiaceae) | x | x | x | x | x | ||||
| 12 | Uapaca guineensis (Euphorbiaceae) | x | x | x | x | x | x | |||
| Species | Height (m) | Root Depth (mm) | PFT 2 | Fabaceae 3 | Foliage C:N (g g−1) | SLA (m2 g−1 C) | |
|---|---|---|---|---|---|---|---|
| 1 | Ceiba pentandra | 55 4 (20) | 610 4,5 (910) | 1 | 0 | 30 | 0.037 ± 0.019 5 (0.020) |
| 2 | Cola acuminata | 20 4 (20) | 910 | 2 6,7 | 0 6,7 | 12 6,7 (30) | 0.041 6,7 (0.030) |
| 3 | Elaeis guineensis | 25 4 (20) | 600 4 (910) | 2 | 0 | 30 | 0.030 |
| 4 | Guibourtia demeusei | 35 4 (20) | 910 | 2 6 | 1 6 | 21 ± 0.7 6 (25) | 0.027 ± 0.006 6 (0.030) |
| 5 | Lophira alata | 55 4 (20) | 1440 | 1 | 0 | 30 | 0.020 |
| 6 | Musanga cecropioides | 30 4,5 (20) | 910 | 2 6 | 0 6 | 18 ± 3.6 6 (30) | 0.026 ± 0.009 5,6 (0.030) |
| 7 | Nauclea diderrichii | 45 4 (20) | 910 | 2 | 0 | 30 | 0.030 |
| 8 | Pericopsis elata | 45 4 (20) | 1140 | 1 | 1 | 25 | 0.020 |
| 9 | Pterocarpus soyauxii | 50 4 (20) | 910 | 2 6 | 1 6 | 16 ± 2.8 6 (25) | 0.048 ± 0.020 6 (0.030) |
| 10 | Pycnanthus angolensis | 30 4 (20) | 910 | 2 6 | 0 6 | 18 ± 4.5 6 (30) | 0.036 ± 0.011 5,6 (0.030) |
| 11 | Symphonia globulifera | 35 4 (20) | 910 | 2 6 | 0 6 | 28 6 (30) | 0.027 ± 0.013 5,6 (0.030) |
| 12 | Uapaca guineensis | 25 4 (20) | 910 | 2 | 0 | 30 | 0.030 |
| Species | Tmins (°C) | Swmins | GDD5ming (°C Day) | Swmaxg | |
|---|---|---|---|---|---|
| 1 | Ceiba pentandra | 6.6 | 0.021 | 5519 | 0.667 |
| 2 | Cola acuminata | 7.6 | 0.093 | 5437 | - |
| 3 | Elaeis guineensis | 6.2 | 0.020 | 5050 | - |
| 4 | Guibourtia demeusei | 10.4 | 0.162 | 6808 | - |
| 5 | Lophira alata | 6.8 | 0.600 | 5803 | 0.675 |
| 6 | Musanga cecropioides | 6.3 | 0.109 | 5408 | - |
| 7 | Nauclea diderrichii | 7.0 | 0.095 | 6144 | - |
| 8 | Pericopsis elata | 8.7 | 0.600 | 6868 | 1.025 |
| 9 | Pterocarpus soyauxii | 11.0 | 0.134 | 6640 | - |
| 10 | Pycnanthus angolensis | 7.2 | 0.069 | 5459 | - |
| 11 | Symphonia globulifera | 5.2 | 0.069 | 4039 | - |
| 12 | Uapaca guineensis | 6.3 | 0.039 | 5653 | - |
| Species | Observed Occurrences 1 | Simulation with PFT-Specific Traits | Simulation with Species-Specific Traits | |||
|---|---|---|---|---|---|---|
| Distribution Area (106 km2) | Sensitivity (%) | Distribution Area (106 km2) | Sensitivity (%) | |||
| 1 | Ceiba pentandra | 197 | 6.59 | 91 | 6.14 | 91 |
| 2 | Cola acuminata | 60 | 6.35 | 98 | 12.76 | 100 |
| 3 | Elaeis guineensis | 118 | 12.87 | 99 | 12.34 | 97 |
| 4 | Guibourtia demeusei | 41 | 5.09 | 100 | 5.42 | 100 |
| 5 | Lophira alata | 128 | 3.94 | 96 | 3.30 | 91 |
| 6 | Musanga cecropioides | 133 | 6.15 | 98 | 7.95 | 99 |
| 7 | Nauclea diderrichii | 126 | 6.12 | 99 | 5.85 | 99 |
| 8 | Pericopsis elata | 40 | 3.93 | 95 | 3.45 | 93 |
| 9 | Pterocarpus soyauxii | 153 | 5.40 | 100 | 8.47 | 99 |
| 10 | Pycnanthus angolensis | 199 | 7.16 | 98 | 10.28 | 100 |
| 11 | Symphonia globulifera | 154 | 7.84 | 99 | 7.52 | 99 |
| 12 | Uapaca guineensis | 162 | 9.25 | 98 | 9.09 | 98 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dury, M.; Mertens, L.; Fayolle, A.; Verbeeck, H.; Hambuckers, A.; François, L. Refining Species Traits in a Dynamic Vegetation Model to Project the Impacts of Climate Change on Tropical Trees in Central Africa. Forests 2018, 9, 722. https://doi.org/10.3390/f9110722
Dury M, Mertens L, Fayolle A, Verbeeck H, Hambuckers A, François L. Refining Species Traits in a Dynamic Vegetation Model to Project the Impacts of Climate Change on Tropical Trees in Central Africa. Forests. 2018; 9(11):722. https://doi.org/10.3390/f9110722
Chicago/Turabian StyleDury, Marie, Lenni Mertens, Adeline Fayolle, Hans Verbeeck, Alain Hambuckers, and Louis François. 2018. "Refining Species Traits in a Dynamic Vegetation Model to Project the Impacts of Climate Change on Tropical Trees in Central Africa" Forests 9, no. 11: 722. https://doi.org/10.3390/f9110722
APA StyleDury, M., Mertens, L., Fayolle, A., Verbeeck, H., Hambuckers, A., & François, L. (2018). Refining Species Traits in a Dynamic Vegetation Model to Project the Impacts of Climate Change on Tropical Trees in Central Africa. Forests, 9(11), 722. https://doi.org/10.3390/f9110722