Litter Inhibitory Effects on Soil Microbial Biomass, Activity, and Catabolic Diversity in Two Paired Stands of Robinia pseudoacacia L. and Pinus nigra Arn.

 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Location and Site Description
2.2. Newly-Shed Litter Sampling
2.3. Forest Floor and Mineral Soil Sampling
2.4. Chemical Analyses of Litter and Soil
2.5. Microbiological Analyses
2.6. Effect of Leaf-Litter Water Extracts on Microbial Activity and Biomass In Mineral Soil
2.7. Statistical Analysis
3. Results
3.1. Litter and Soil Chemical Characteristics
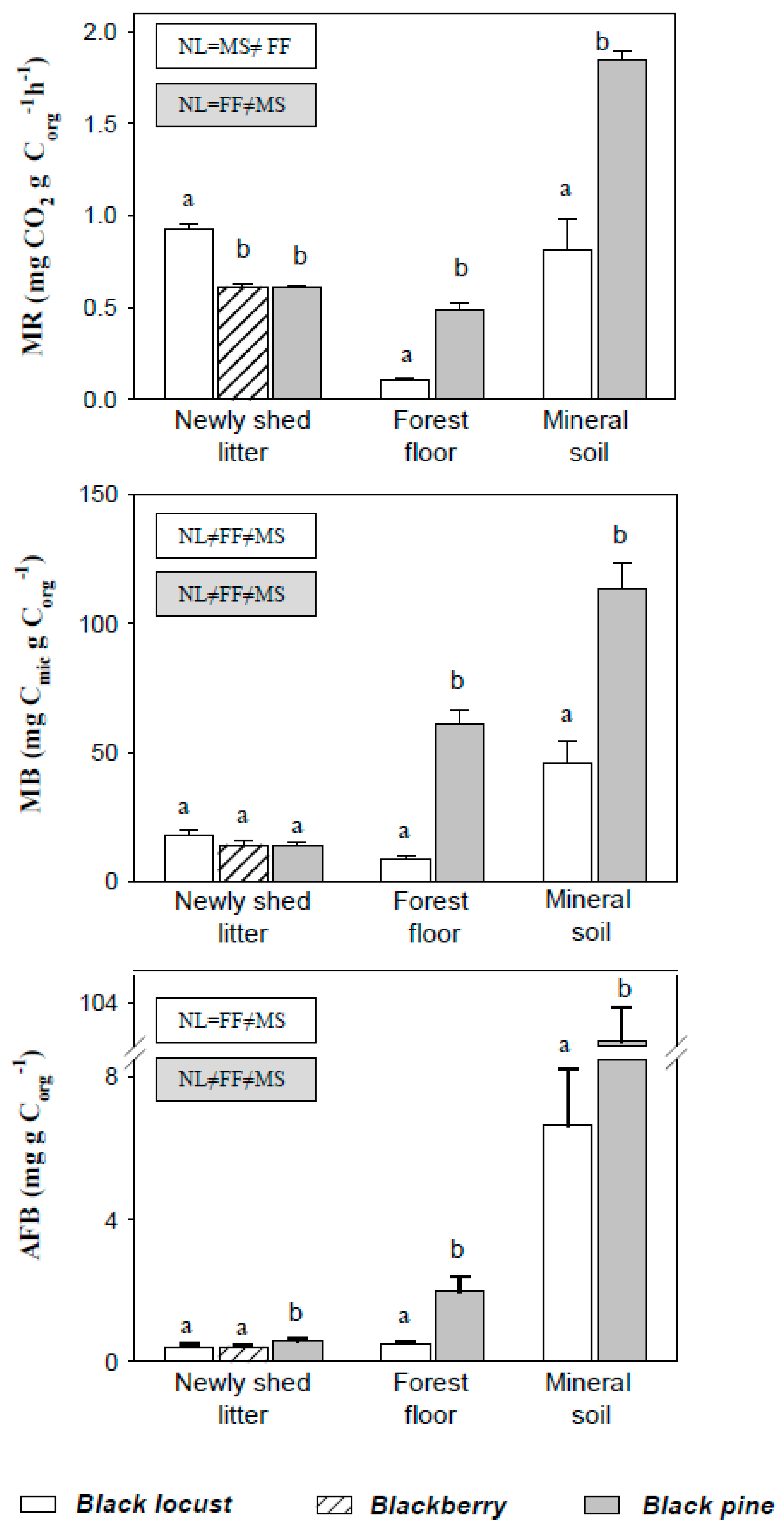
3.2. Mineralization Rate, Microbial Biomass, Active Fungal Biomass
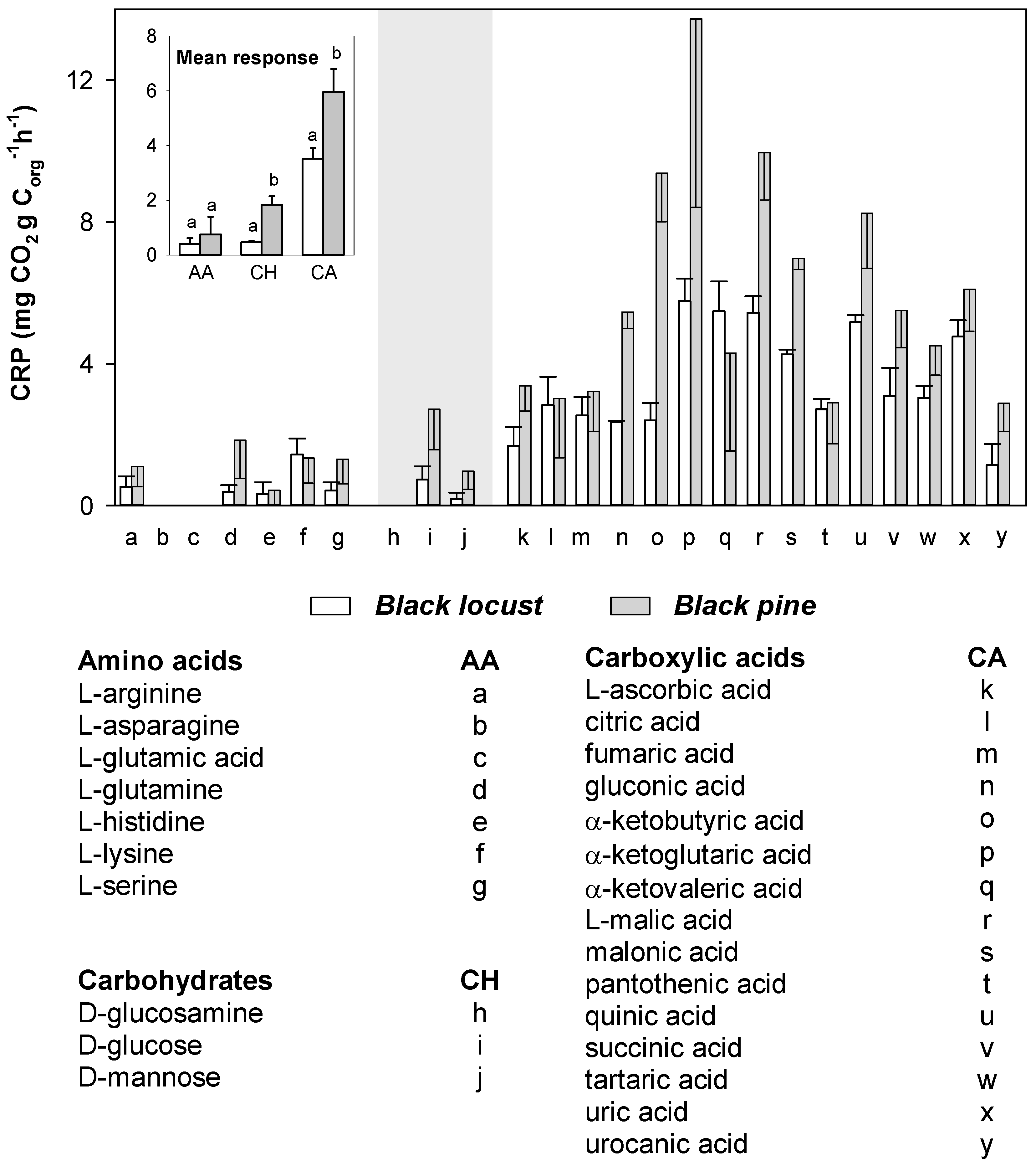
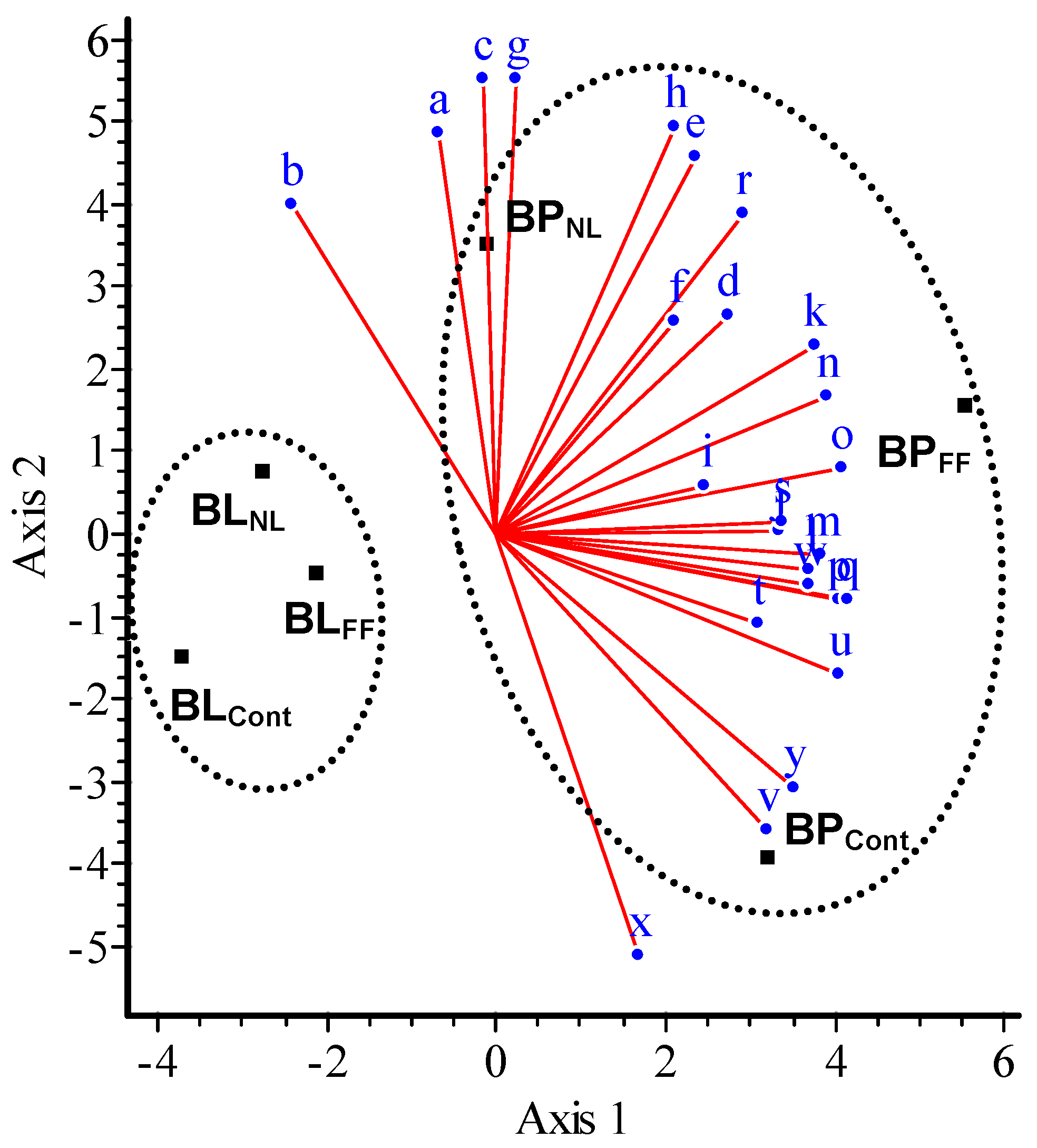
3.3. Catabolic Response Profile of Mineral Soil
3.4. Organic Components of Litter Water Extracts
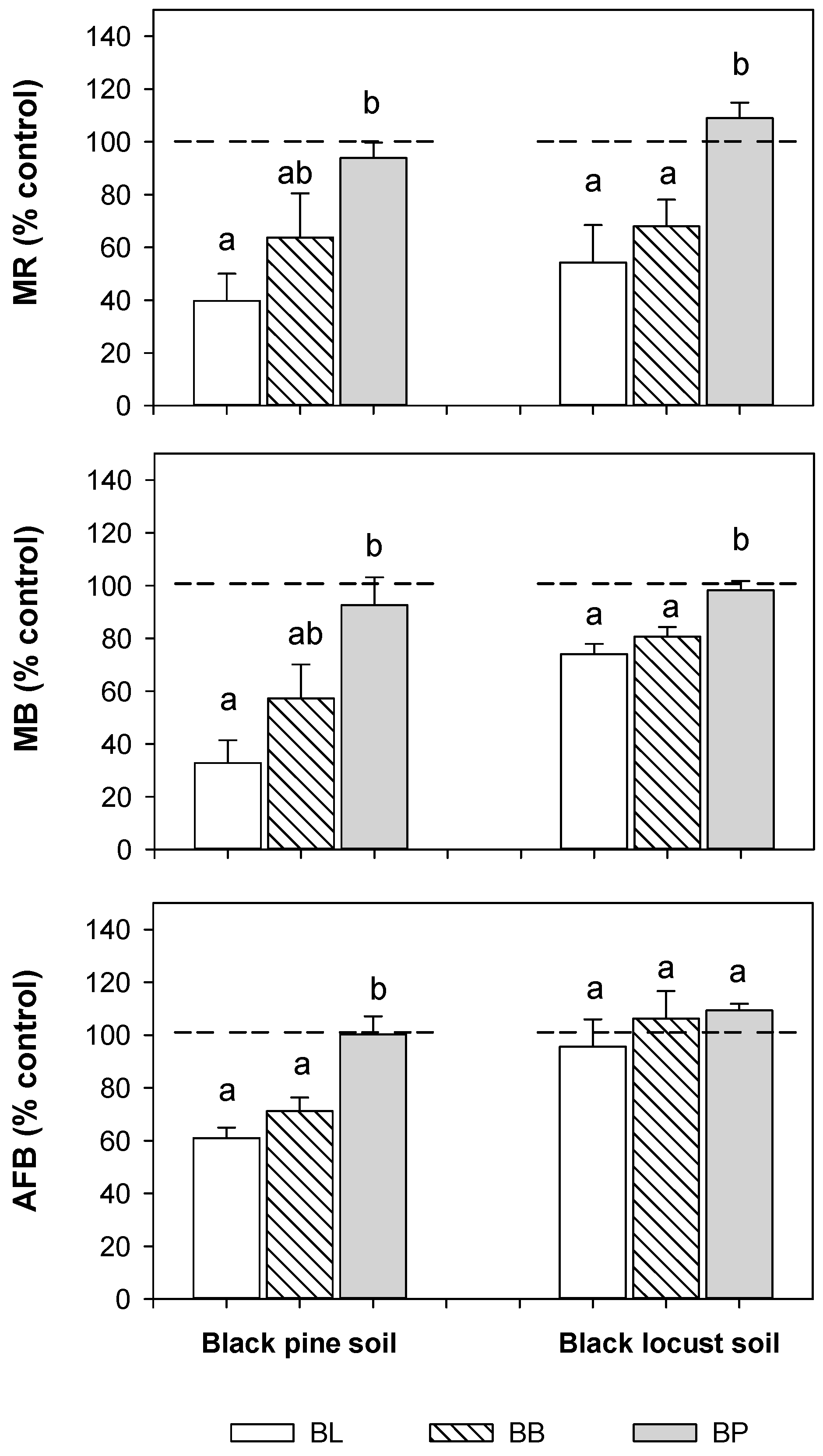
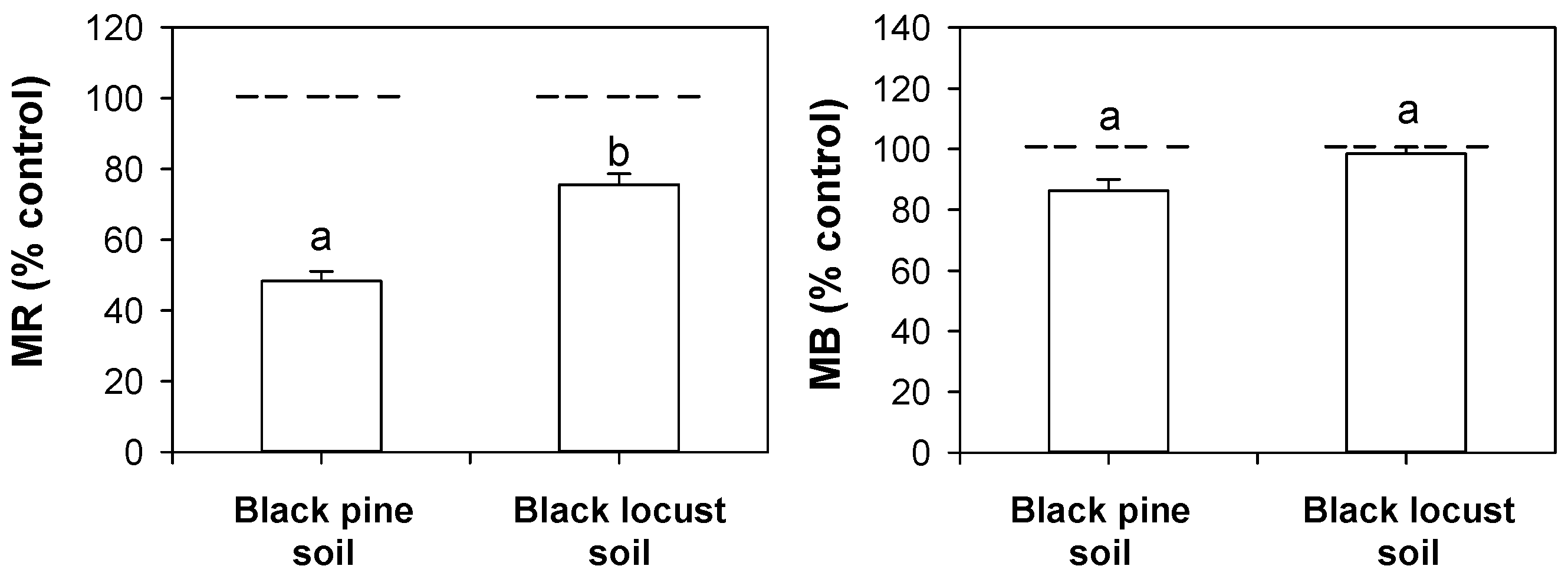
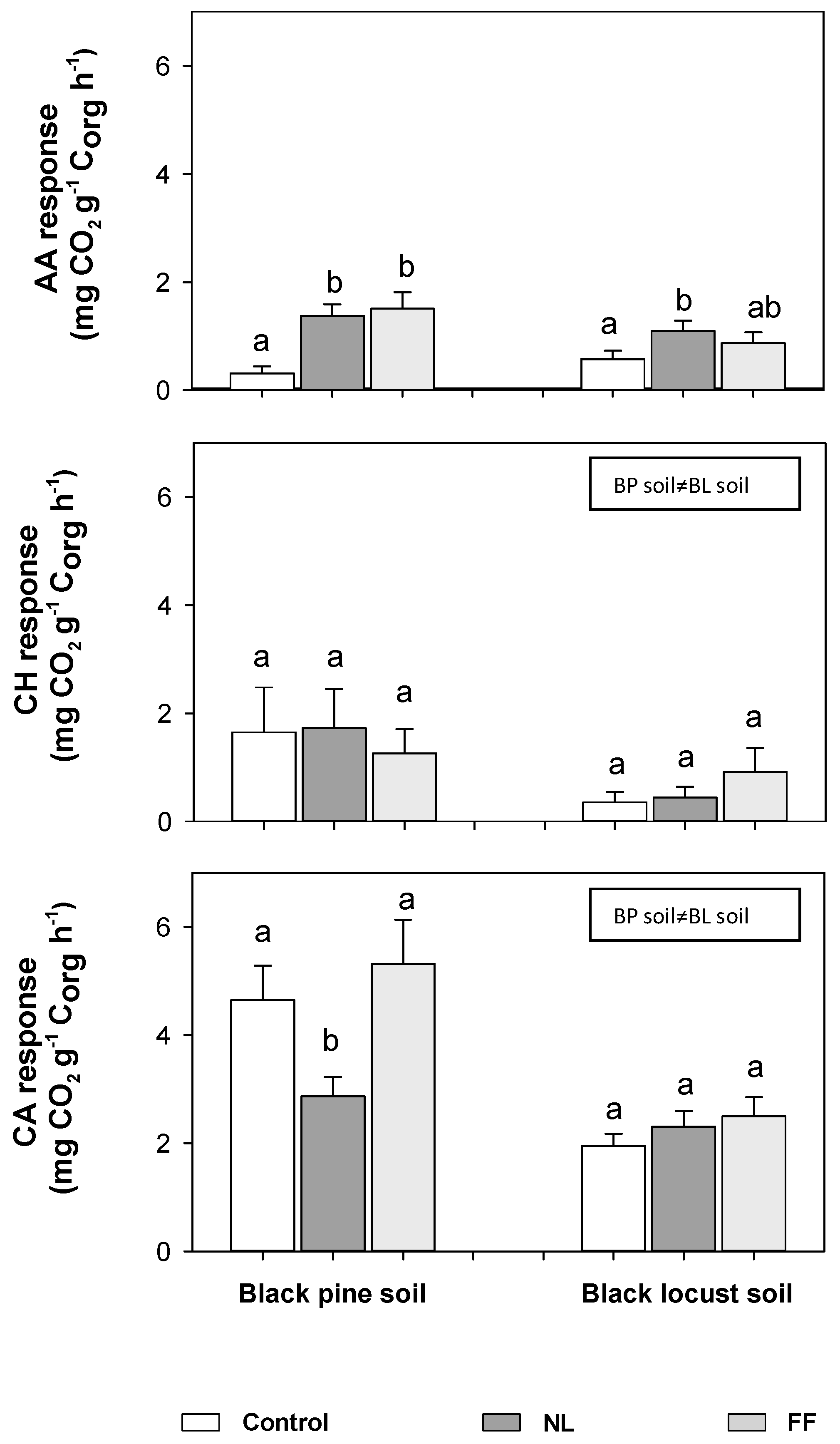
3.5. Effects of Litter Water Extracts on Mineralization Rates, Microbial Biomass, and Active Fungal Biomass of the Mineral Soil
3.6. Changes in the Catabolic Response Profile of Mineral Soil Amended with Litter Water Extracts
4. Discussion
4.1. Some Main Factors Influencing Carbon Sequestration in Forest Soils and Their Roles in The Present Study
4.2. Are There Additional Factors for Carbon Sequestration-Allelopathy?
5. Conclusions
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Rutigliano, F.A.; D’Ascoli, R.; Virzo De Santo, A. Soil microbial metabolism and nutrient status in a Mediterranean area as affected by plant cover. Soil Biol. Biochem. 2004, 36, 1719–1729. [Google Scholar] [CrossRef]
- Vesterdal, L.; Clarke, N.; Sigurdsson, B.D.; Gundersen, P. Do tree species influence soil carbon stocks in temperate and boreal forests? For. Ecol. Manag. 2013, 309, 4–18. [Google Scholar] [CrossRef]
- Augusto, L.; De Schrijver, A.; Vesterdal, L.; Smolander, A.; Prescott, C.; Ranger, J. Influences of evergreen gymnosperm and deciduous angiosperm tree species on the functioning of temperate and boreal forests. Biol. Rev. 2014, 90, 444–466. [Google Scholar] [CrossRef] [PubMed]
- Prescott, C.E.; Grayston, S.J. Tree species influence on microbial communities in litter and soil: Current knowledge and research needs. For. Ecol. Manag. 2013, 309, 19–27. [Google Scholar] [CrossRef]
- Schimel, J.P.; Van Cleve, K.; Cates, R.G.; Clausen, T.P.; Reichardt, P.B. Effects of balsam poplar (Populus balsamifera) tannins and low molecular weight phenolics on microbial activity in taiga floodplain soil: Implication for changes in N cycling during succession. Can. J. Bot. 1995, 74, 84–90. [Google Scholar] [CrossRef]
- Nilsson, M.C.; Gallet, C.; Wallstedt, A. Temporal variability of phenolics and batatasin III in Empetrum hermaphroditum leaves over an eight-year period: Interpretation of ecological function. Oikos 1998, 81, 6–16. [Google Scholar] [CrossRef]
- Coté, J.F.; Thibault, J.R. Allelopathic potential of raspberry foliar leachates on growth of ectomycorrhizal fungi associated with black spruce. Am. J. Bot. 1988, 75, 966–970. [Google Scholar] [CrossRef]
- Nilsson, M.C. The Mechanisms of Biological Interference by Empetrum Hermaphroditum on Tree Seedling Establishment in Boreal Forest Ecosystems. Ph.D. Thesis, Swedish University of Agricultural Science, Umeå, Sweden, 1992. [Google Scholar]
- Amarowicz, R.; Dykes, G.A.; Pegg, R.B. Antibacterial activity of tannin constituents from Phaseolus vulgaris, Fagoypyrum esculentum, Corylus avellana and Juglans nigra. Fitoterapia 2008, 79, 217–219. [Google Scholar] [CrossRef] [PubMed]
- Souto, C.; Pellissier, F.; Chiapusio, G. Relationships between phenolics and Soil Microorganisms in Spruce Forests: Significance for Natural Regeneration. J. Chem. Ecol. 2000, 26, 2025–2034. [Google Scholar] [CrossRef]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K.A. How strongly can forest management influence soil carbon sequestration? Geoderma 2007, 137, 253–268. [Google Scholar] [CrossRef]
- De Marco, A.; Spaccini, R.; Vittozzi, P.; Esposito, F.; Berg, B.; Virzo De Santo, A. Decomposition of black locust and black pine leaf litter in two coeval forest stands on Mount Vesuvius and dynamics of organic components assessed through proximate analysis and NMR spectroscopy. Soil Biol. Biochem. 2012, 51, 1–15. [Google Scholar] [CrossRef]
- De Marco, A.; Esposito, F.; Berg, B.; Giordano, M.; Virzo De Santo, A. Soil C and N sequestration in organic and mineral layers of two coeval forest stands implanted on pyroclastic material (Mount Vesuvius, South Italy). Geoderma 2013, 209–210, 128–135. [Google Scholar] [CrossRef]
- Berg, B.; Davey, M.P.; De Marco, A.; Emmett, B.; Faituri, M.; Hobbie, S.E.; Johansson, M.-B.; Liu, C.; McClaugherty, C.; Norell, L.; et al. Factors influencing limit values for pine needle litter decomposition: A synthesis for boreal and temperate pine forest systems. Biogeochemistry 2010, 100, 57–73. [Google Scholar] [CrossRef]
- Vesterdal, L.; Schmidt, I.K.; Callesen, I.; Nilsson, L.O.; Gundersen, P. Carbon and nitrogen in forest floor and mineral soil under six common European tree species. For. Ecol. Manag. 2008, 255, 35–48. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C.; Virzo De Santo, A. Practicalities of estimating carbon sequestration in boreal coniferous forests. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2008. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Bonanomi, G.; Sicurezza, M.G.; Caporaso, S.; Esposito, A.; Mazzoleni, S. Phytotoxicity dynamics of decaying plant materials. New Phytol. 2006, 169, 571–578. [Google Scholar] [CrossRef]
- Nasir, H.; Iqbal, Z.; Hiradate, S.; Fujii, Y. Allelopathic potential of Robinia pseudo-acacia L. J. Chem. Ecol. 2005, 31, 2179–2192. [Google Scholar] [CrossRef]
- Esposito, F.; De Marco, A.; Zarrelli, A.; Virzo De Santo, A. Allelopathic effects of Black locust (Robinia pseudoacacia L.) litter on soil microbial activity. In Proceedings of the 5th World Congress on Allelopathy: Growing Awareness of The Role of Allelopathy in Ecological, Agricultural, and Environmental Processes, Saratoga Springs, NY, USA, 21–25 September 2008; p. 87. [Google Scholar]
- Von Holle, B.; Neill, C.; Largay, E.F.; Budreski, K.A.; Ozimec, B.; Clark, S.A.; Lee, K. Ecosystem legacy of the introduced N2-fixing tree Robinia pseudoacacia in a coastal forest. Oecologia 2013, 172, 915–924. [Google Scholar] [CrossRef]
- De Marco, A.; Arena, C.; Giordano, M.; Virzo De Santo, A. Impact of the invasive tree black locust on soil properties of Mediterranean stone pine-holm oak forests. Plant Soil 2013, 372, 473–486. [Google Scholar] [CrossRef]
- Panizzi, L.; Caponi, C.; Catalano, S.; Cioni, P.L.; Morelli, I. In vitro antimicrobial activity of extracts and isolated constituents of Rubus ulmifolius. J. Ethnopharmacol. 2002, 79, 165–168. [Google Scholar] [CrossRef]
- Riaz, M.; Ahmad, M.; Rahman, N. Antimicrobial screening of fruit, leaves, root and stem of Rubus fruticosus. J. Med. Plants Res. 2011, 5, 5920–5924. [Google Scholar]
- Cseresnyés, I.; Tamás, J. Evaluation of Austrian pine (Pinus nigra) plantations in Hungary with respect to nature conservation—A review (Review). J. Landsc. Ecol. 2014, 12, 267–284. [Google Scholar]
- Berg, B.; Hannus, K.; Popoff, T.; Theander, O. Chemical components of Scots pine needles and needle litter and inhibition of fungal species by extractives. In Structure and Function of Northern Coniferous Forests—An Ecosystem Study; Persson, T., Ed.; Swedish Natural Science Reasearch Council: Stockholm, Sweden, 1980; pp. 391–400. [Google Scholar]
- Selvakumar, G.; Supradip, S.; Kundu, S. Inhibitory activity of pine needle tannin extracts on some agriculturally resourceful microbes. Indian J. Microbiol. 2007, 47, 267–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzoleni, S.; Ricciardi, M. Primary succession on the cone of Vesuvius. In Primary Succession on Land; Miles, J., Walton, D.W.H., Eds.; The British Ecological Society: London, UK, 1993; pp. 101–112. [Google Scholar]
- di Gennaro, A. I Sistemi di Terre Della Campania; S.EL.CA.: Firenze, Italy, 2002. [Google Scholar]
- Dick, W.A.; Tabatabai, M.A. Hydrolysis of organic and inorganic phosphorus compounds added to soil. Geoderma 1978, 21, 175–182. [Google Scholar] [CrossRef]
- García, C.; Hernández, T.; Costa, F. Study on water extract of sewage sludge composts. Soil Sci. Plant Nutr. 1991, 37, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Violante, P. (Ed.) Ministero delle Politiche Agricole e Forestali–Osservatorio Nazionale Pedologico e per la Qualità del Suolo, Metodi ufficiali di analisi chimica del suolo; International Union of Soil Science and Società Italiana della Scienza del Suolo; Franco Angeli: Milano, Itay, 2000. [Google Scholar]
- Mathers, N.J.; Jalota, R.K.; Dalal, R.C.; Boyd, S.E. 13C-NMR analysis of decomposing litter and fine roots in the semiarid Mulga Lands of southern Queensland. Soil Biol. Biochem. 2007, 39, 993–1006. [Google Scholar] [CrossRef]
- Degens, B.P.; Schipper, L.A.; Sparling, G.P.; Vojvodic Vukovic, M. Decreases in organic C reserves in soils can reduce the catabolic diversity of soil microbial communities. Soil Biol. Biochem. 2000, 32, 189–196. [Google Scholar] [CrossRef]
- Degens, B.P.; Schipper, L.A.; Sparling, G.P.; Duncan, L.C. Is the microbial community in a soil with reduced catabolic diversity less resistant to stress or disturbance? Soil Biol. Biochem. 2001, 33, 1143–1153. [Google Scholar] [CrossRef]
- Sundman, V.; Sivelä, S. A comment on the membrane filter technique for estimation of length of fungal hyphae in soil. Soil Biol. Biochem. 1978, 10, 399–401. [Google Scholar] [CrossRef]
- Söderström, B. Vital staining of fungi in pure cultures and in soil with fluorescein-diacetate. Soil Biol. Biochem. 1977, 11, 237–246. [Google Scholar] [CrossRef]
- Berg, B.; Söderström, B. Fungal biomass and nitrogen in decomposing Scots pine needle litter. Soil Biol. Biochem. 1979, 11, 339–341. [Google Scholar] [CrossRef]
- D’Abrosca, B.; DellaGreca, M.; Fiorentino, A.; Monaco, P.; Natale, A.; Oriano, P.; Zarrelli, A. Structural characterization of phytotoxic terpenoids from Cestrum parqui. Phytochemistry 2005, 66, 2681–2688. [Google Scholar] [CrossRef] [PubMed]
- Podani, J. SYN-Tax-pc. Computer Programs for Multivariate Data Analysis in Ecology and Systematics, Version 5.0; Scientia Publishing: Budapest, Hungary, 1993.
- Berg, B.; McClaugherty, C. Plant Litter. Decomposition, Humus Formation, Carbon Sequestration; Springer: Berlin Heidelberg, Germany, 2014. [Google Scholar]
- Güsewell, S.; Verhoeven, T.A. Litter N:P ratios indicate whether N or P limits the decomposability of graminoid leaf litter. Plant Soil 2006, 287, 131–143. [Google Scholar] [CrossRef]
- Gui, H.; Hyde, K.; Xu, J.; Mortimer, P. Arbuscular mycorrhiza enhance the rate of litter decomposition while inhibiting soil microbial community development. Sci. Rep. 2017, 7, 42184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, M.E.; Turner, B.L.; Liang, C.; Clay, K.; Johnson, D.J.; Phillips, R.P. Tree mycorrhizal type predicts within-site variability in the storage and distribution of soil organic matter. Glob. Chang. Biol. 2018, 24, 3317–3330. [Google Scholar] [CrossRef] [PubMed]
- Wilson, G.W.T.; Rice, C.W.; Rillig, M.C.; Springer, A.; Hartnett, D.C. Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: Results from long-term field experiments. Ecol. Lett. 2009, 12, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Uselman, S.M.; Qualls, R.G.; Thomas, R.B. Effects of increased atmospheric CO2, temperature, and soil N availability on root exudation of dissolved organic carbon by a N-fixing tree (Robinia pseudoacacia L.). Plant Soil 2000, 222, 191–202. [Google Scholar] [CrossRef]
- Lukić, S.; Pantić, D.; Belanović Simić, S.; Borota, D.; Tubić, B.; Djukić, M.; Djunisijević-Bojović, D. Effects of black locust and black pine on extremely degraded sites 60 years after afforestation—A case study of the Grdelica Gorge (Southeastern Serbia). iForest 2015. [Google Scholar] [CrossRef]
- Mueller, K.E.; Hobbie, S.E.; Chorover, J.; Reich, P.B.; Eisenhauer, N.; Castellano, M.J.; Chadwick, O.A.; Dobies, T.; Hale, C.M.; Jagodzin′ski, A.M.; et al. Effects of litter traits, soil biota, and soil chemistry on soil carbon stocks at a common garden with 14 tree species. Biogeochemistry 2015, 123, 313–327. [Google Scholar] [CrossRef] [Green Version]
- Adamczyk, B.; Karonen, M.; Adamczyk, S.; Engström, M.T.; Laakso, T.; Saranpää, P.; Kitunen, V.; Smolander, A.; Simon, J. Tannins can slow-down but also speed-up soil enzymatic activity in boreal forest. Soil Biol. Biochem. 2017, 107, 60–67. [Google Scholar] [CrossRef]
- Sosa, T.; Valares, C.; Alías, J.C.; Chaves Lobón, N. Persistence of flavonoids in Cistus ladanifer soils. Plant Soil 2010, 337, 51–63. [Google Scholar] [CrossRef]
- Medina-Villar, S.; Álvaro, A.; Castro-Díez, P.; Pérez-Corona, M.E. Allelopathic potentials of exotic invasive and native trees over coexisting understory species: The soil as modulator. Plant Ecol. 2017, 218, 579–594. [Google Scholar] [CrossRef]
- Kuzyakov, Y. Priming effects: Interactions between living and dead organic matter. Soil Biol. Biochem. 2010, 42, 1363–1371. [Google Scholar] [CrossRef]
- Jones, D. Organic acids in the rhizosphere—A critical review. Plant Soil 1998, 205, 25–44. [Google Scholar] [CrossRef]
- Van Hees, P.A.W.; Jones, D.L.; Godbold, D.L. Biodegradation of low molecular weight organic acids in coniferous forest podzolic soils. Soil Biol. Biochem. 2002, 34, 1261–1272. [Google Scholar] [CrossRef]
- Zhang, J.; Guang-Cai, C.; Xing, S.; Sun, Q.; Shan, Q.; Zhou, J.; Wang, Y. Carbon sequestration of black locust forests in the Yellow River Delta region, China. Int. J. Sustain. Dev. World Ecol. 2010, 17, 475–480. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH-H2O | Ash/CEC | P | Mn | N | C-to-N | N-to-P | Soluble C | Soluble N | Soluble N | Soluble C-to-N | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| mg g−1 | µg g−1 | mg g−1 | mg g−1 | mg g−1 | % of total N | ||||||
| Newly-shed | Ash (mg g−1) | ||||||||||
| Black locust | - | § 7.0 ± 0.3 a | 1.7 ± 0.6 a | § 42.1 ± 7.9 a | § 24.6 ± 0.4 a | § 19.5 ± 0.9 a | 14.5 ± 0.8 a | 122.3 ± 4.4 a | 10.6 ± 0.9 a | 43.1 ± 1.5 a | 11.7 ± 0.9 a |
| Blackberry | - | §§ 2.0 ± 0.3 b | 0.8 ± 0.07 a | §§ 65.2 ± 1.9 b | §§ 16.9 ± 0.3 b | §§ 27.4 ± 1.3 b | 21.2 ± 0.5 b | 107.3 ± 3.5 a | 7.1 ± 0.3 b | 28.8 ± 1.1 b | 15.2 ± 0.5 ab |
| Black pine | - | § 2.8 ± 0.9 b | 2.56 ± 0.8 b | § 29.0 ± 2.0 a | § 23.0 ± 1.8 a | § 22.1 ± 1.0 a | 9.0 ± 1.8 c | 57.7 ± 5.8 b | 3.4 ± 0.4 c | 14.0 ± 1.4 c | 17.0 ± 1.3 b |
| Forest floor | |||||||||||
| Black locust | 6.03 ± 0.13 a | 4.5 ± 0.6 a | 0.51 ± 0.11 a | 57.4 ± 3.0 a | 23.01 ± 0.61 a | 16.20 ± 0.98 a | 45.1 ± 0.8 a | 81.54 ± 2.31 a | 9.05 ± 1.03 a | 39.46 ± 1.56 a | 8.90 ± 1.33 a |
| Black pine | 4.75 ± 0.17 b | 6.9 ± 0.8 b | 1.11 ± 0.18 b | 62.1 ± 13.1 a | 13.40 ± 1.52 b | 31.57 ± 3.15 b | 12.1 ± 1.1 b | 16.40 ± 0.82 b | 2.49 ± 0.62 b | 18.67 ± 1.63 b | 5.96 ± 1.25 a |
| Mineral soil | CEC (cmolc Kg−1) | ||||||||||
| Black locust | § 5.59 ± 0.10 a | §§ 12.19 | 0.43 ± 0.06 a | § 18.6 ± 0.9 a | § 5.20 ± 0.58 a | 6.48 ± 0.79 a | 12.1 ± 0.8 a | 9.11 ± 0.90 a | 3.22 ± 0.13 a | 61.92 ± 1.24 a | 2.81 ± 1.11 a |
| Black pine | § 4.17 ± 0.18 b | §§ 4.92 | 0.28 ± 0.04 b | § 12.1 ± 1.4 b | § 3.10 ± 0.18 b | 8.86 ± 1.19 a | 11.1 ± 0.6 a | 5.77 ± 1.21 a | 1.12 ± 0.10 b | 36.13 ± 0.98 b | 5.24 ± 0.89 b |
| Newly-Shed Litter | Forest-Floor Litter | ||||
|---|---|---|---|---|---|
| Black Locust | Blackberry | Black Pine | Black Locust Stand | Black Pine Stand | |
| Total C (mg g−1) | 480.5 ± 10.0 a,A | 463.4 ± 10.3 a,B | 509.1 ± 15.0 b,A | 425.9 ± 11.2 a,B | 471.0 ± 18.1 b,A |
| Carboxyl C (200–160 ppm) | 29.3 ± 0.1 a,A | 26.9 ± 0.1 a,A | 18.8 ± 0.1 b,A | 39.2 ± 1.3 a,B | 24.5 ± 1.2 b,B |
| Phenol C (160–145 ppm) | 34.1 ± 0.6 a,A | 27.3 ± 0.1 b,B | 25.5 ± 0.2 b,A | 32.6 ± 1.1 a,A | 29.2 ± 1.4 a,B |
| Aryl C (145–110 ppm) | 48.5 ± 0.8 a,A | 53.8 ± 0.3 b,A | 44.8 ± 0.3 a,A | 44.1 ± 1.4 a,B | 39.6 ± 1.9 b,B |
| O – Alkyl C (110–60 ppm) | 258.5 ± 4.2 a,A | 243.3 ± 1.2 a,A | 312.6 ± 1.8 b,A | 199.1 ± 6.4 a,B | 263.4 ± 9.9 b,B |
| Methoxyl C (60–45 ppm) | 29.8 ± 0.5 a,A | 30.1 ± 0.1 a,A | 25.5 ± 0.2 b,A | 33.8 ± 1.1 a,B | 34.8 ± 1.6 a,B |
| Alkyl C (45–0 ppm) | 80.2 ± 1.3 a,A | 82.0 ± 0.4 a,A | 82.0 ± 0.6 a,A | 76.9 ± 2.5 a,A | 86.6 ± 4.1 b,A |
| Methoxyl C to Phenol C | 0.87 ± 0.008 a,A | 1.11 ± 0.001 b,A | 1.00 ± 0.0001c,A | 1.03 ± 0.007 a,B | 1.19 ± 0.0001 b,B |
| Aromaticity index | 0.32 ± 0.002 a,A | 0.33 ± 0.001 a,A | 0.22 ± 0.002 b,A | 0.38 ± 0.003 a,B | 0.26 ± 0.007 b,B |
| Hydrophobicity index | 0.56 ± 0.0001 a,A | 0.61 ± 0.005 b,A | 0.46 ± 0.006 c,A | 0.65 ± 0.004 a,B | 0.55 ± 0.003 b,B |
| Water Extract Components (%) | Newly-Shed Litter | Forest-Floor Litter | Mineral Soil | ||||
|---|---|---|---|---|---|---|---|
| Black Locust | Blackberry | Black Pine | Black Locust Stand | Black Pine Stand | Black Locust Stand | Black Pine Stand | |
| Carboxylic ac./Aldehydes (12–8 ppm) | 8.11 | 15.39 | 4.71 | 12.44 | 4.54 | 5.35 | 3.27 |
| Aromatic compounds (8–6 ppm) | 5.41 | 12.31 | 5.88 | 9.61 | 14.50 | 8.23 | 5.23 |
| Carbohydrates (6–4 ppm) | 59.46 | 49.23 | 58.83 | 51.16 | 44.56 | 53.50 | 45.75 |
| O-Alkyls (4–3 ppm) | 5.41 | 6.15 | 7.06 | 7.80 | 4.73 | 12.35 | 6.54 |
| Alkyls (3–0 ppm) | 21.61 | 16.92 | 23.53 | 18.99 | 31.67 | 20.58 | 39.22 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Marco, A.; Esposito, F.; Berg, B.; Zarrelli, A.; Virzo De Santo, A. Litter Inhibitory Effects on Soil Microbial Biomass, Activity, and Catabolic Diversity in Two Paired Stands of Robinia pseudoacacia L. and Pinus nigra Arn. Forests 2018, 9, 766. https://doi.org/10.3390/f9120766
De Marco A, Esposito F, Berg B, Zarrelli A, Virzo De Santo A. Litter Inhibitory Effects on Soil Microbial Biomass, Activity, and Catabolic Diversity in Two Paired Stands of Robinia pseudoacacia L. and Pinus nigra Arn. Forests. 2018; 9(12):766. https://doi.org/10.3390/f9120766
Chicago/Turabian StyleDe Marco, Anna, Fabrizio Esposito, Björn Berg, Armando Zarrelli, and Amalia Virzo De Santo. 2018. "Litter Inhibitory Effects on Soil Microbial Biomass, Activity, and Catabolic Diversity in Two Paired Stands of Robinia pseudoacacia L. and Pinus nigra Arn." Forests 9, no. 12: 766. https://doi.org/10.3390/f9120766
APA StyleDe Marco, A., Esposito, F., Berg, B., Zarrelli, A., & Virzo De Santo, A. (2018). Litter Inhibitory Effects on Soil Microbial Biomass, Activity, and Catabolic Diversity in Two Paired Stands of Robinia pseudoacacia L. and Pinus nigra Arn. Forests, 9(12), 766. https://doi.org/10.3390/f9120766