Calcium and Potassium Imbalance Favours Leaf Blight and Defoliation Caused by Calonectria pteridis in Eucalyptus Plants

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Preparation and Implantation
2.2. Inoculum Preparation and Inoculation
2.3. Experimental Evaluation
2.3.1. Disease and Defoliation Assessment
2.3.2. Plant Growth and Chlorophyll Content
2.4. Determination of the Leaf Nutrient Contents
2.5. Statistical Analysis
3. Results
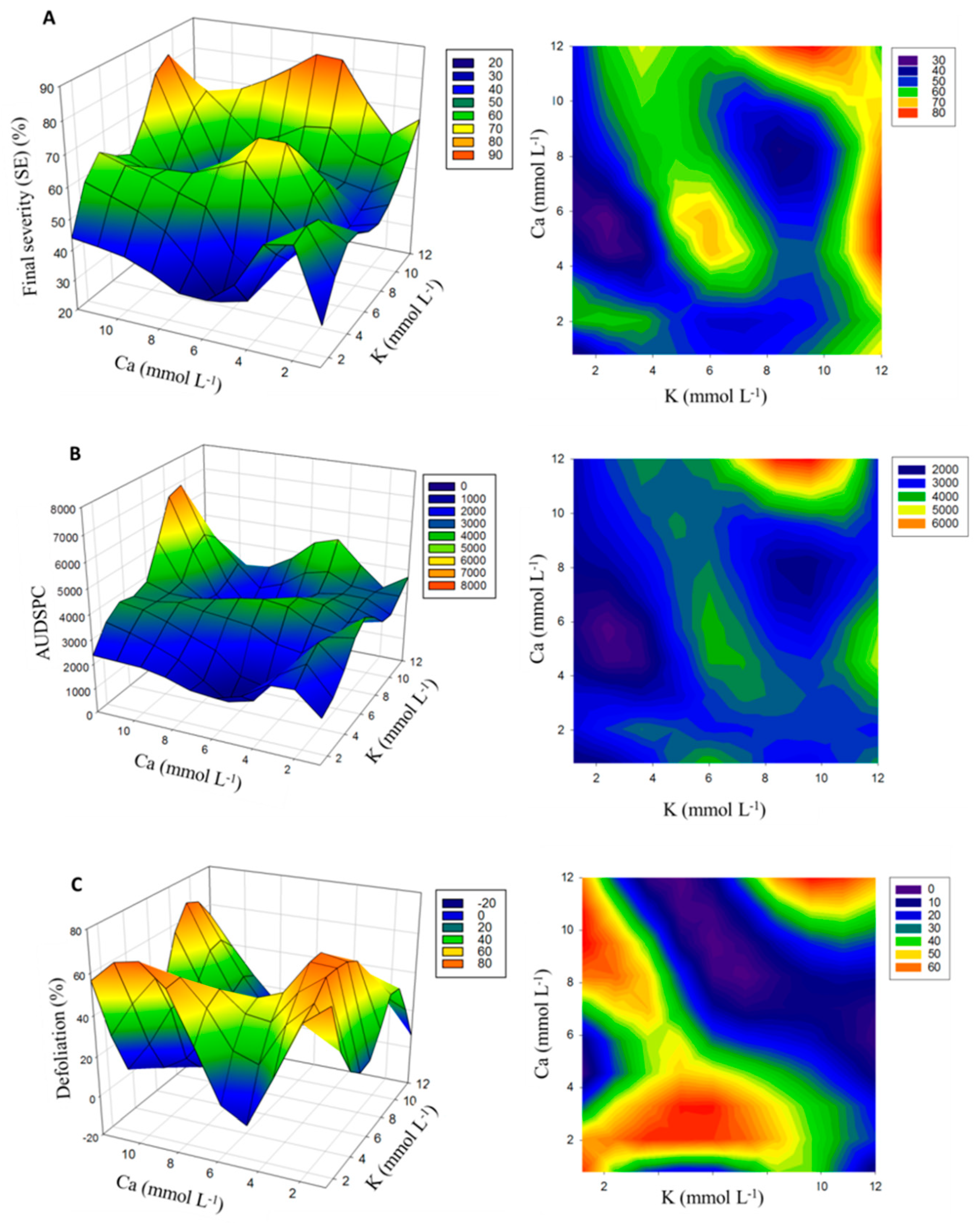
3.1. Disease Severity
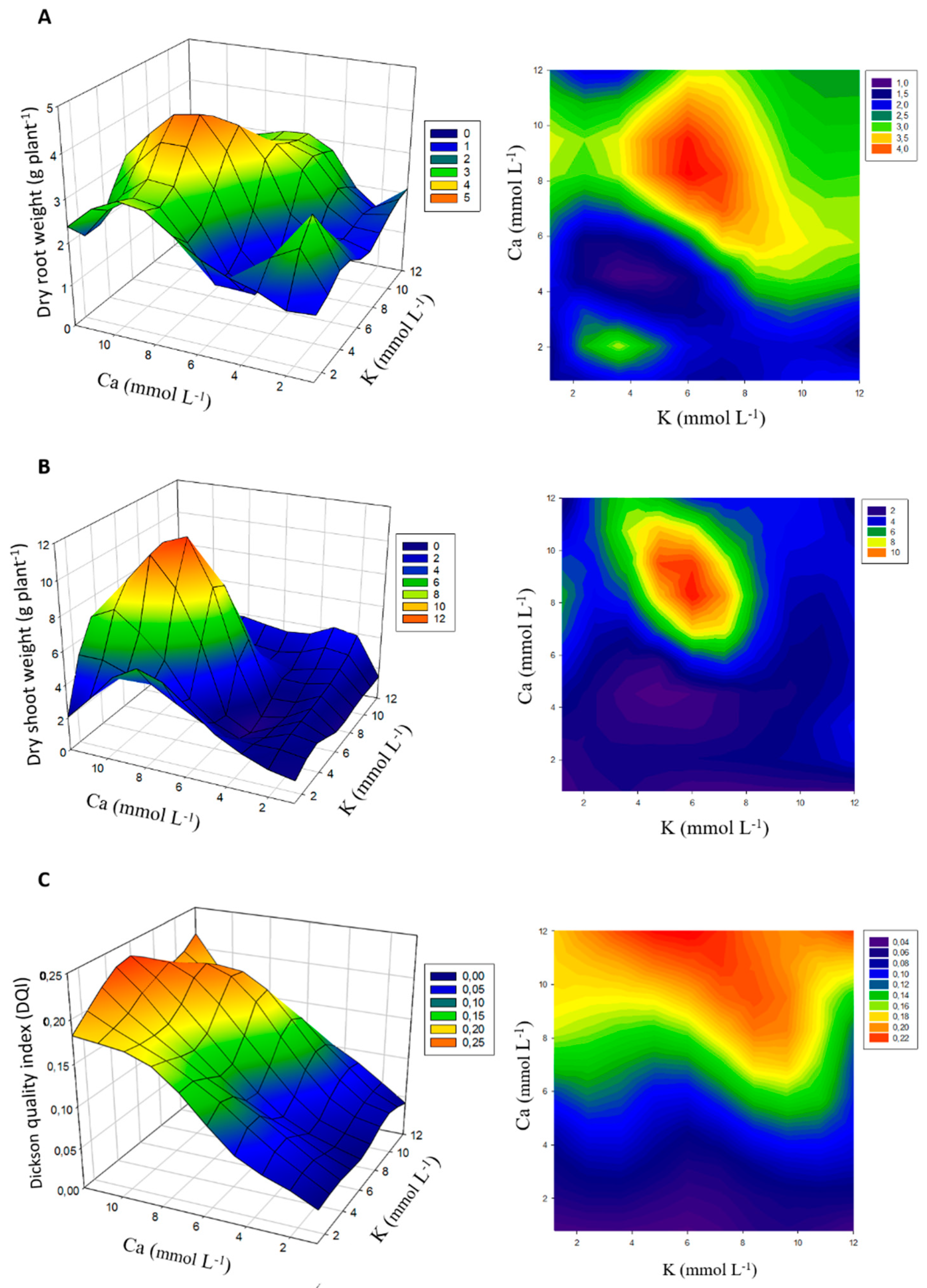
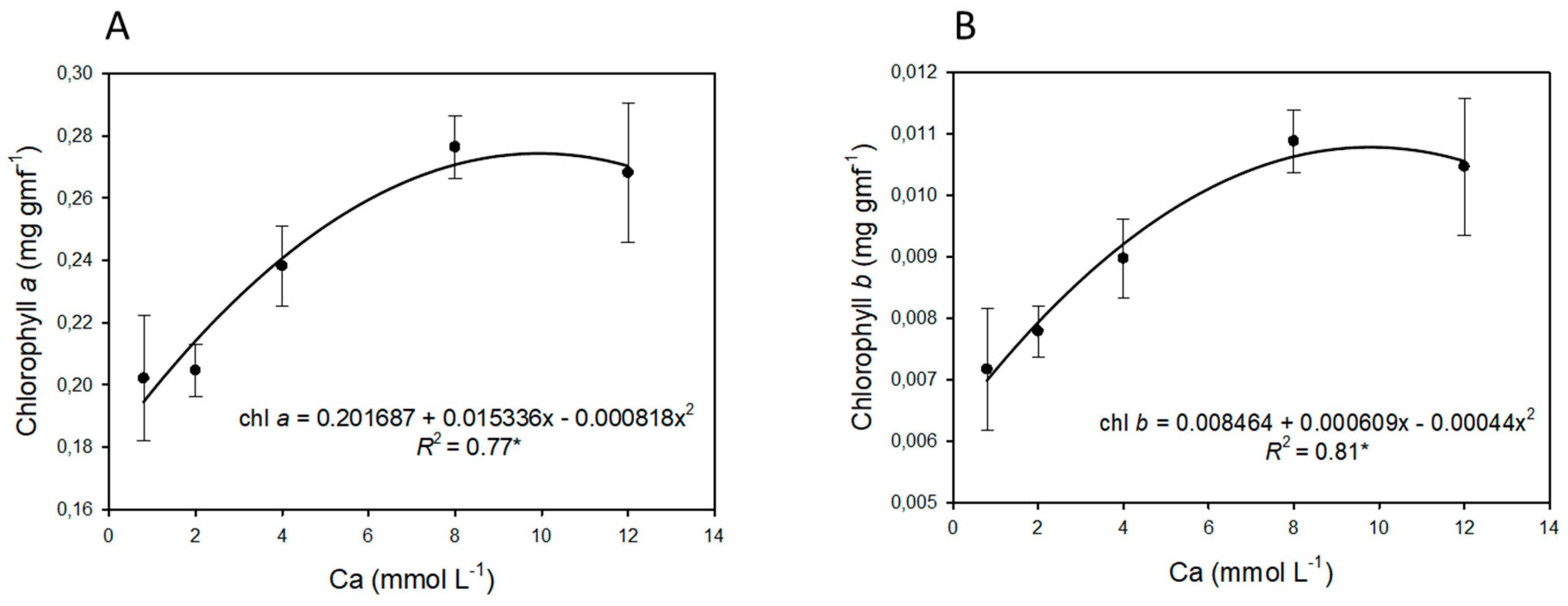
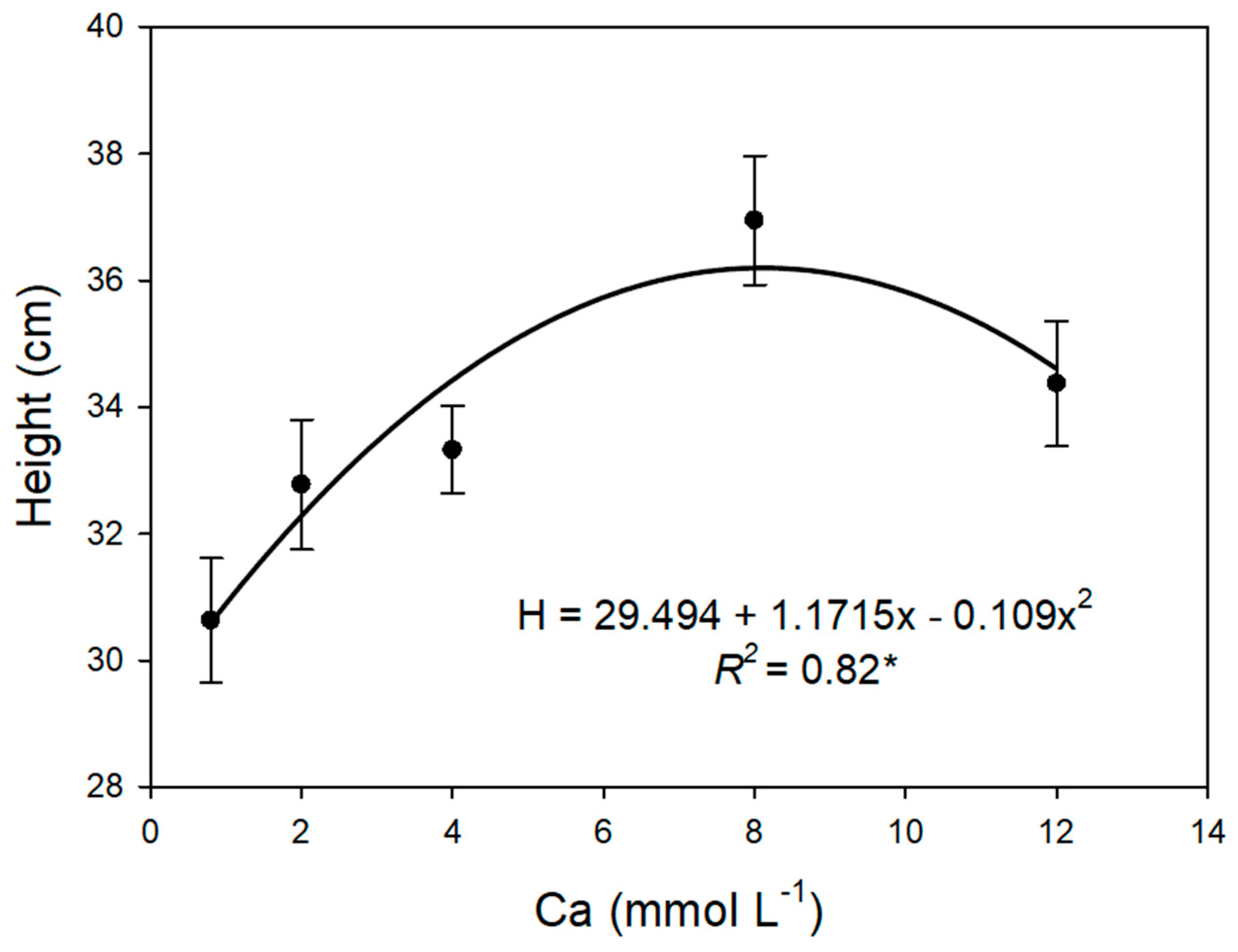
3.2. Plant Growth and Chlorophyll Content
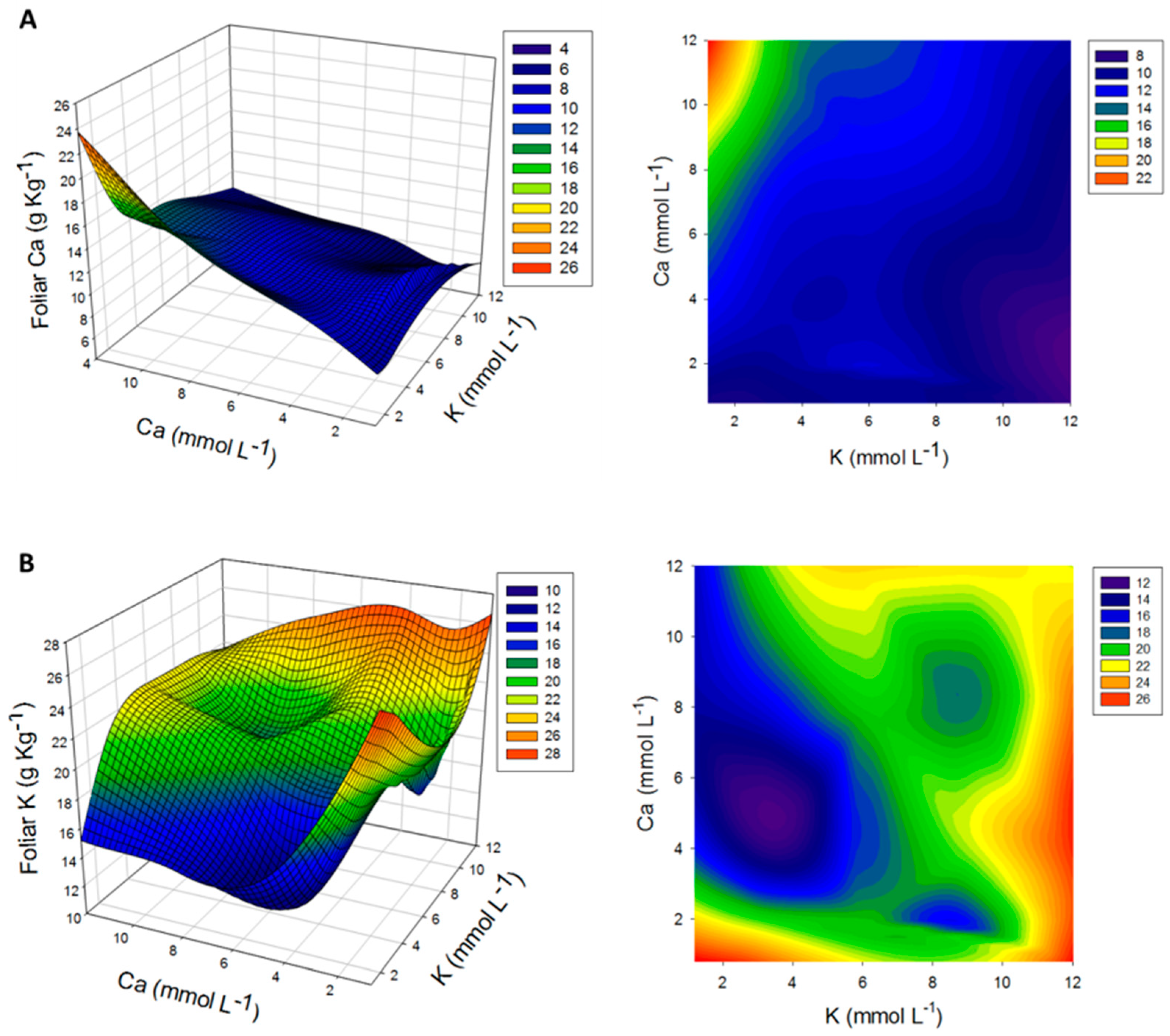
3.3. Determination of Leaf Nutrient Contents
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ibá—Indústria Brasileira de Árvores. Available online: http://iba.org/images/shared/Biblioteca/IBA_RelatorioAnual2017.pdf (accessed on 3 October 2017).
- Hinchee, M.; Rottmann, W.; Mullinax, L.; Zhang, C.; Chang, S.; Cunningham, M.; Pearson, L.; Nehra, N. Short-rotation woody crops for bioenergy and biofuels applications. In Biofuels; Springer: New York, NY, USA, 2009; pp. 139–156. [Google Scholar]
- Shepherd, M.; Bartle, J.; Lee, D.J.; Brawner, J.; Bush, D.; Turnbull, P. Eucalypts as a biofuel feedstock. Biogeosciences 2011, 2, 639–657. [Google Scholar] [CrossRef]
- Alfenas, A.C.; Zauza, E.A.V.; Mafia, R.G.; Assis, T.F. Clonagem e Doenças do Eucalipto, 2nd ed.; Editora, U.F.V., Viçosa, M.G., Eds.; Editora UFV: Viçosa, Brazil, 2009; p. 500. [Google Scholar]
- Rodas, C.A.; Lombard, L.; Gryzenhout, M.; Slippers, B.; Wingfield, M.J. Cylindrocladium blight of. Eucalyptus grandis in Colombia. Australas. Plant Pathol. 2005, 34, 143–149. [Google Scholar] [CrossRef]
- Alfenas, A.C.; Ferreira, F.A. A mancha de folha do eucalipto no Brasil causada por três espécies de Cylindrocladium—Uma revisão da descrição da doença. Revista Árvore 1979, 3, 47–56. [Google Scholar]
- Ferreira, F.A.; Alfenas, A.C.; Moreira, A.M.; Demuner, N.L. Mancha-depteridis doença foliar de eucalipto em areas tropicais brasileiras. Fitopatol. Bras. 1995, 20, 107–110. [Google Scholar]
- Alfenas, A.C.; Zauza, E.A.V.; Mafia, R.G.; Assis, T.F. Clonagem e Doenças do Eucalipto; Editora UFV: Viçosa, Brazil, 2004; p. 442. [Google Scholar]
- Ferreira, E.M.; Alfenas, A.C.; Mafia, L.A.; Mafia, R.G. Eficiência de fungicidas sistêmicos para o controle de Cylindrocladium candelabrum em eucalipto. Fitopatol. Bras. 2006, 31, 468–475. [Google Scholar] [CrossRef]
- Datnoff, L.E.; Rodrigues, F.A.; Seebold, K.W. Silicon and Plant Nutrition. In Mineral Nutrition and Plant Disease; Datnoff, L.E., Elmer, W.H., Huber, D.M., Eds.; APS Press: Saint Paul, MN, USA, 2007; pp. 233–246. [Google Scholar]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, T.J. Interaction between Plant Nutrients: 1. Theory and Analytical Procedures. Acta Agric. Scand. Section B–Soil Plant Sci. 1992, 42, 208–212. [Google Scholar] [CrossRef]
- Jakobsen, T.J. Interaction between Plant Nutrients: III. Antagonism between Potassium, Magnesium and Calcium. Acta Agric. Scand. Section B–Soil Plant Sci. 1993, 43, 1–5. [Google Scholar] [CrossRef]
- Zhang, L.; Du, L.; Poovaiah, B.W. Calcium signaling and biotic defense responses in plants. Plant Signal. Behav. 2014, 9, e973818. [Google Scholar] [CrossRef] [Green Version]
- Armengaud, P.; Breitling, R.; Amtmann, A. The potassium-dependent transcriptome of Arabidopsis reveals a prominent role of jasmonic acid in nutrient signaling. Plant Physiol. 2004, 136, 2556–2576. [Google Scholar] [CrossRef]
- Ma, W.; Qi, Z.; Smigel, A.; Walker, R.K.; Verma, R.; Berkowitz, G.A. Ca2+, cAMP, and transduction of nonself perception during plant immune responses. Proc. Nat. Acad. Sci. USA 2009, 106, 20995–21000. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Punja, Z.K. Calcium and plant disease. In Mineral Nutrition and Plant Disease; Datnoff, L.E., Elmer, W.H., Huber, D.M., Eds.; American Phytopathological Society Press: Saint Paul, MN, USA, 2007; Volume 1, pp. 79–93. [Google Scholar]
- Clark, R.B. Characterization of phosphates in intact maize roots. J. Agric. Food Chem. 1975, 23, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Alfenas, R.F. Produção de Inóculo de Cylindrocladium pteridis em Condições Controladas. Master’s Thesis, Federal University of Viçosa, Viçosa, Brazil, October 2009. [Google Scholar]
- Vale, F.X.R.; Fernandes, F.E.I.F.; Liberato, J.R. QUANT—A software for plant disease severity assessment. In Proceedings of the Anais International Congress of Plant Pathology, Christchurch, New Zealand, 2–7 February 2003. [Google Scholar]
- Dickson, A.; Leaf, A.L.; Hosner, J.F. Quality appraisal of white spruce and white pine seedling stock in nurseries. For. Chron. 1960, 36, 10–13. [Google Scholar] [CrossRef]
- Shaner, G.; Finney, R.E. The effect of nitrogen fertilization on the expression of slow-mildew resistance in Knox wheat. Phytopathology 1977, 67, 1051–1056. [Google Scholar] [CrossRef]
- Matsumoto, S.N.; Carvalho, F.M.; Viana, A.E.S.; Malta, M.R.; Castro, L.G. Initial growth of coffee plants (Coffea Arabica L.) submitted to different phosphate doses in nutritive solution. Coffee Sci. 2008, 31, 58–67. [Google Scholar]
- Malavolta, E.; Vitti, G.C.; Oliveira, S.A. Avaliação do Estado Nutricional das Plantas: Princípios e Aplicações, 2nd ed.; POTAFOS: Piracicaba, Brazil, 1997; p. 319. [Google Scholar]
- Martinez, H.E.P.; Carvalho, J.G.; Souza, R.B. Diagnose foliar. In Recomendações para Uso de Corretivos e Fertilizantes em Minas Gerais: 5 Aproximação; Ribeiro, A.C., Guimarães, P.T.G., Alvarez, V.V.H., Eds.; CFSEMG: Viçosa, Brazil, 1999; pp. 143–168. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Pulrolnik, K.; Reis, G.G.; Reis, M.G.F.; Monte, M.A.; Fontan, I.C.I. Crescimento de plantas de clone de Eucalyptus grandis [hillexmaiden] submetidas a diferentes tratamentos de desrama artificial, na região de cerrado. Revista Árvore 2005, 29, 495–505. [Google Scholar] [CrossRef]
- Hillocks, R.J.; Chindoya, R. The relationship between Alternaria leaf spot and potassium deficiency causing premature defoliation of cotton. Plant Pathol. 1989, 38, 502–508. [Google Scholar] [CrossRef]
- Hu, W.; Lv, X.; Yang, J.; Chen, B.; Zhao, W.; Meng, Y.; Oosterhuis, D.M. Effects of potassium deficiency on antioxidant metabolism related to leaf senescence in cotton (Gossypium hirsutum L.). Field Crop. Res. 2016, 191, 139–149. [Google Scholar] [CrossRef]
- Tsialtas, J.T.; Shabala, S.; Matsi, T. A prominent role for leaf calcium as a yield and quality determinan in upland cotton (Gossypium hirsutum L.) varieties grown under irrigated Mediterranean conditions. J. Agron. Crop Sci. 2016, 202, 161–173. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Regulation of potassium transport in plants under hostile conditions: Implications for abiotic and biotic stress tolerance. Physiol. Plant. 2014, 151, 257–279. [Google Scholar] [CrossRef]
- Reddy, K.R.; Hodges, H.F.; Varco, J. Potassium nutrition of cotton. Miss. Agric. For. Exp. Stn. 2000, 1094, 1–10. [Google Scholar]
- Pervez, H.; Ashraf, M.; Makhdum, M.I.; Mahmood, T. Potassium nutrition of cotton (Gossypium hirsutum L.) in relation to cotton leaf curl virus disease in aridisols. Pak. J. Bot. 2007, 39, 529–539. [Google Scholar]
- Graça, R.N.; Alfenas, A.C.; Maffia, L.A.; Titon, M.; Alfenas, R.F.; Lau, D.; Rocabado, J.M.A. Factors influencing infection of eucalypts by Cylindrocladium pteridis. Plant Pathol. 2009, 58, 971–981. [Google Scholar] [CrossRef]
- Smith, S.; Stewart, G.R. Effects of potassium levels on the stomatal behavior of the hemi-parasite Striga hermonthica. Plant Physiol. 1990, 94, 1472–1476. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, F.A.V. Crescimento de Eucalipto em Idade Jovem e Movimentação de Cálcio e Magnésio no Solo em Resposta à Aplicação de Calcário e Gesso Agrícola. Ph.D. Thesis, Universidade Federal de Viçosa, Viçosa, Brazil, April 2013. [Google Scholar]
- Ivashuta, S.; Liu, J.; Lohar, D.P.; Haridas, S.; Bucciarelli, B.; VandenBosch, K.A.; Vance, C.P.; Harrison, M.J.; Gantt, J.S. RNA interference identifies a calcium-dependent protein kinase involved in Medicago truncatula root development. Plant Cell 2005, 17, 2911–2921. [Google Scholar] [CrossRef] [PubMed]
- Fromm, J. Wood formation of trees in relation to potassium and calcium nutrition. Tree Physiol. 2010, 30, 1140–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fageria, V.D. Nutrient interactions in crop plants. J. Plant Nutr. 2001, 24, 1269–1290. [Google Scholar] [CrossRef]
- Laclau, J.P.; Almeida, J.C.R.; Gonçalves, J.L.M.; Saint-Andre, L.; Ventura, M.; Ranger, J.; Moreira, R.M.; Nouvellon, Y. Influence of nitrogen and potassium fertilization on leaf life span and allocation of above-ground growth in Eucalyptus plantations. Tree Physiol. 2009, 29, 111–124. [Google Scholar] [CrossRef]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, T.P.F.; Pozza, E.A.; Pozza, A.A.A.; Mafia, R.G.; Ferreira, M.A. Calcium and Potassium Imbalance Favours Leaf Blight and Defoliation Caused by Calonectria pteridis in Eucalyptus Plants. Forests 2018, 9, 782. https://doi.org/10.3390/f9120782
Soares TPF, Pozza EA, Pozza AAA, Mafia RG, Ferreira MA. Calcium and Potassium Imbalance Favours Leaf Blight and Defoliation Caused by Calonectria pteridis in Eucalyptus Plants. Forests. 2018; 9(12):782. https://doi.org/10.3390/f9120782
Chicago/Turabian StyleSoares, Thaissa P. F., Edson A. Pozza, Adélia A. A. Pozza, Reginaldo Gonçalves Mafia, and Maria A. Ferreira. 2018. "Calcium and Potassium Imbalance Favours Leaf Blight and Defoliation Caused by Calonectria pteridis in Eucalyptus Plants" Forests 9, no. 12: 782. https://doi.org/10.3390/f9120782
APA StyleSoares, T. P. F., Pozza, E. A., Pozza, A. A. A., Mafia, R. G., & Ferreira, M. A. (2018). Calcium and Potassium Imbalance Favours Leaf Blight and Defoliation Caused by Calonectria pteridis in Eucalyptus Plants. Forests, 9(12), 782. https://doi.org/10.3390/f9120782