Ecophysiological Responses of Three Tree Species to a High-Altitude Environment in the Southeastern Tibetan Plateau

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Leaf Gas Exchange
2.3. Leaf Water Potential
2.4. Photosynthetic Light Response Curves
2.5. Diurnal Course of Chlorophyll Fluorescence
2.6. Survival Rate, Height, Biomass
2.7. Data Analysis
3. Results
3.1. Morphological Characteristics
3.2. Water Relations
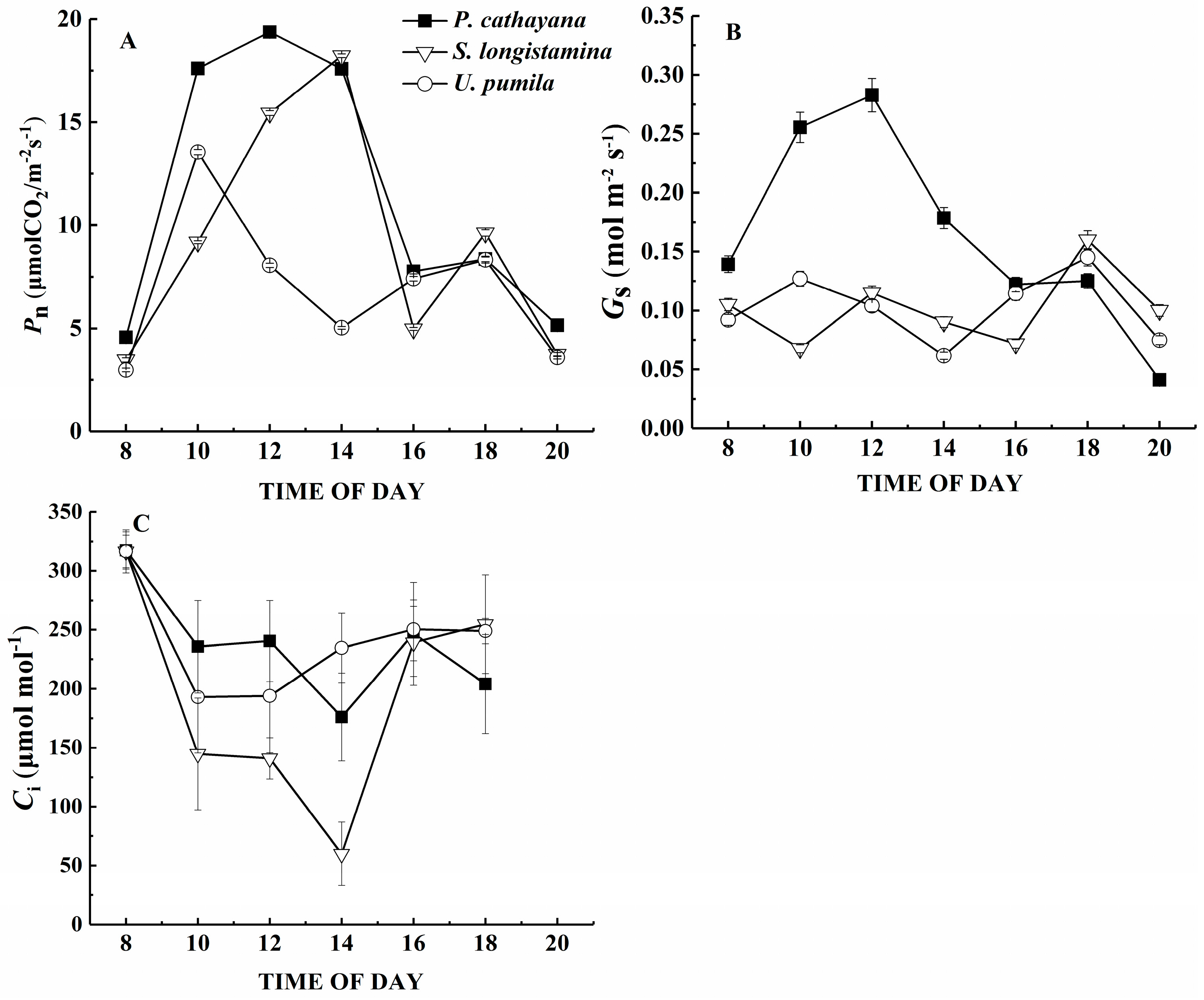
3.3. Gas Exchange
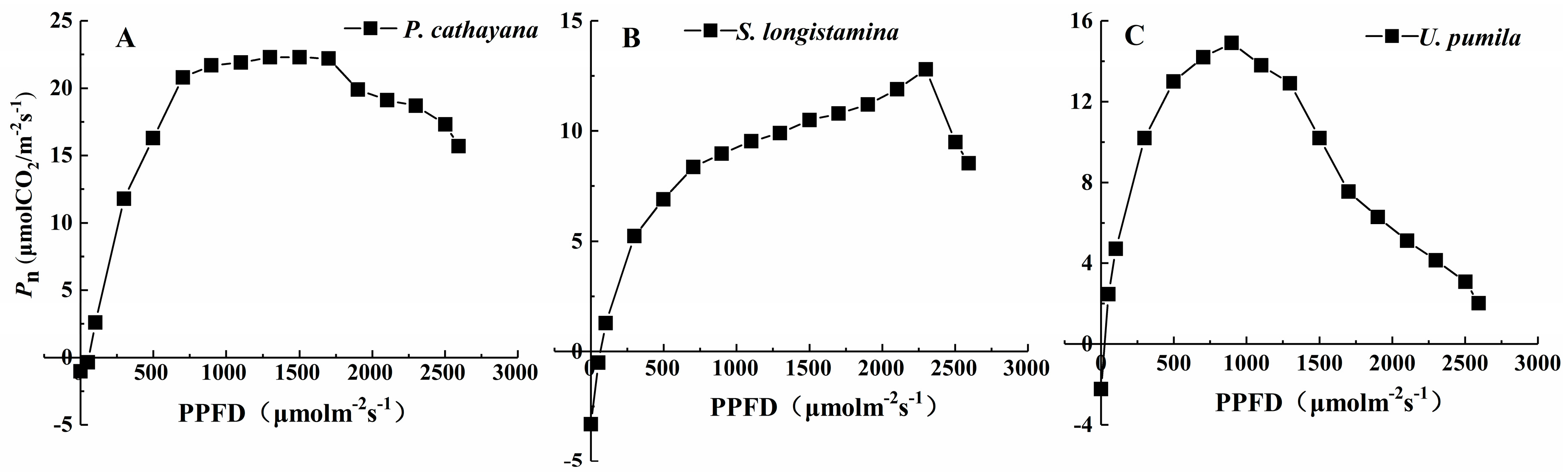
3.4. Pn-Light Response Curves
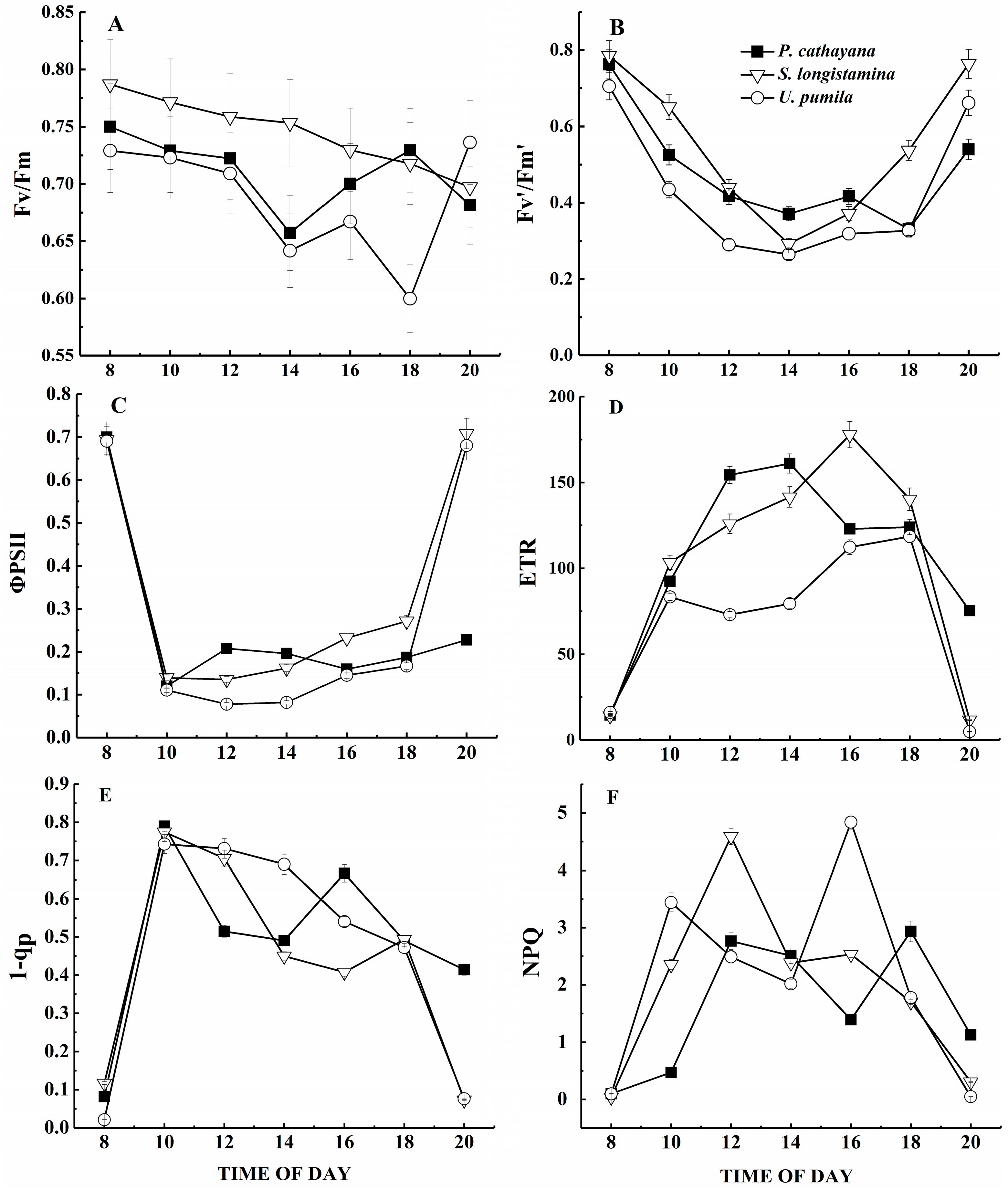
3.5. Chlorophyll Fluorescence Parameters
4. Discussion
4.1. Morphological Characteristics
4.2. Water Relations
4.3. Gas Exchange
4.4. Photosynthesis Curve
4.5. Photochemistry Efficiency
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Devi, N.; Agedorn, F.; Moiseev, P.; Bugmann, H.; Shiyatov, S.; Mazepa, V.; Rigling, A. Expanding forests and changing growth forms of Siberian larch at the Polar Urals tree line during the 20th century. Glob. Chang. Biol. 2008, 14, 1581–1591. [Google Scholar] [CrossRef]
- Qian, C.; Yin, H.; Shi, Y.; Zhao, J.; Yin, C.; Luo, W. Population dynamics of Agriophyllum squarrosum, a pioneer annual plant endemic to mobile sand dunes, in response to global climate change. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Okunlola, G.O.; Olusanya, A.O. Physiological response of the three most cultivated pepper species (capsicum spp.) in Africa to drought stress imposed at three stages of growth and development. Sci. Hortic. 2017, 224, 198–205. [Google Scholar] [CrossRef]
- Liang, E.Y.; Wang, Y.F.; Xu, Y. Growth variation in Abies georgei var. smithii along altitudinal gradients in the Sygera Mountains, Southeastern Tibetan Plateau. Trees 2010, 24, 363–373. [Google Scholar] [CrossRef]
- Najafabadi, M.Y.; Ehsanzadeh, P. Photosynthetic and antioxidative upregulation in drought-stressed sesame (Sesamum indicum L.) subjected to foliar-applied salicylic acid. Photosynthetica 2017, 4, 611–622. [Google Scholar] [CrossRef]
- Wonsick, M.M.; Pinker, R.T. The radiative environment of the Tibetan plateau. Int. J. Clim. 2014, 34, 2153–2162. [Google Scholar] [CrossRef]
- Roupioz, L.; Jia, L.; Nerry, F.; Menenti, M. Estimation of daily solar radiation budget at kilometer resolution over the Tibetan plateau by integrating Modis data products and a DEM. Remote Sens. 2016, 8, 504. [Google Scholar] [CrossRef]
- Chen, C.; Peng, Y.H. AFLP analysis of genetic diversity in Populus cathayana Rehd. originating from Southeastern Qinghai–Tibetan Plateau of China. Pak. J. Bot. 2010, 42, 117–127. [Google Scholar]
- Song, Y.; Ma, K.; Ci, D.; Zhang, Z.; Zhang, D. Biochemical, physiological and gene expression analysis reveals sex-specific differences in Populus tomentosa floral development. Physiol. Plant. 2013, 150, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Li, Z.; Wu, F.; Tang, M. Comparative photochemistry activity and antioxidant responses in male and female Populus cathayana cuttings inoculated with arbuscular mycorrhizal fungi under salt. Sci. Rep. 2016, 6, 37663. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Li, Y.; Liu, W.; Xu, H.; Sun, O.J. Changes in water use with growth in Ulmus pumila, in semiarid sandy land of northern China. Trees 2014, 28, 41–52. [Google Scholar] [CrossRef]
- Park, G.; Lee, D.; Kim, K.; Batkhuu, N.O.; Tsogtbaatar, J.; Zhu, J.J.; Jin, Y.; Park, P.S.; Hyun, O.; Kim, H.S. Morphological characteristics and water-use efficiency of siberian elm trees (Ulmus pumila L.) within arid regions of northeast Asia. Forests 2016, 7, 280. [Google Scholar] [CrossRef]
- Guillermo, N.; Doffo, S.E.; Monteoliva, M.E.; Rodríguez, V.M.; Luquez, C. Physiological responses to alternative flooding and drought stress episodes in two willow (Salix spp.) clones. Can. J. For. Res. 2016, 47, 174–182. [Google Scholar] [CrossRef]
- Karrenberg, S.; Edwards, P.J.; Kollmann, J. The life history of Salicaceae living in the active zone of floodplains. Freshw. Biol. 2002, 47, 733–748. [Google Scholar] [CrossRef]
- Pusz, W.; Urbaniak, J. Foliar diseases of willows (Salix spp.) in selected locations of the Karkonosze mts. (the giant mts). Eur. J. Plant Pathol. 2017, 148, 45–51. [Google Scholar] [CrossRef]
- Yang, H.; Chu, J.; Lu, Q.; Gao, T. Relationships of native trees with grasses in a temperate, semi-arid sandy ecosystem of northern China. Appl. Veg. Sci. 2014, 17, 338–345. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, L.; Zhou, K.; Li, Y.; Zhao, Z. Changes in protein profile of Platycladus orientalis, (L.) roots and leaves in response to drought stress. Tree Genet. Genomes 2017, 13, 76. [Google Scholar] [CrossRef]
- Robakowski, P.; Laitat, E. Effects of an enhanced ultraviolet–B irradiation on photosynthetic apparatus of several forests coniferous tree species from different locations. Acta Physiol. Plant. 1999, 21, 283–296. [Google Scholar] [CrossRef]
- Lukaszek, M.; Poskuta, J. Chlorophyll a fluorescence kinetics and CO2 exchange rates in light and in darkness in leaves of tall fescue as affected by UV radiation. Acta Physiol. Plant. 1996, 18, 345–350. [Google Scholar]
- Liu, X.; Wang, Z.; Bao, W.; Li, X.M. Photosynthetic responses of two pleurocarpous mosses to low-level nitrogen addition: A study in an old-growth fir forest. J. Bryol. 2015, 37, 15–22. [Google Scholar] [CrossRef]
- Bilger, W.G.; Schreiber, U.; Bock, M. Determination of the quantum efficiency of photosystem II and of non–photochemical quenching of chlorophyll fluorescence in the field. Oecologia 1995, 102, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Drake, B.G.; Read, M. Carbon dioxide assimilation, photosynthetic efficiency, and respiration of a Chesapeake Bay salt marsh. J. Ecol. 1981, 69, 405–423. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Van Kooten, O.; Snel, J.F.H. The use of fluorescence nomenclature in plant stress physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Bilger, W.; Björkman, O. Role of the xanthophylls cycle in photoprotection elucidated by measurements of light–induced absorbance changes, fluorescence and photosynthesis in Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Demmig, B.; Björkman, O. Comparison of the effects of excessive light on chlorophyll fluorescence (77K) and photon yield of O2 evolution in leaves of higher plants. Planta 1987, 171, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Ceulemans, R.; Deraedt, W. Production physiology and growth potential of poplars under short-rotation forestry culture. For. Ecol. Manag. 1999, 121, 9–23. [Google Scholar] [CrossRef]
- Gong, J.R.; Zhang, X.S.; Huang, Y.M. Comparison of the performance of several hybrid poplar clones and their potential suitability for use in northern China. Biomass Bioenergy 2011, 35, 2755–2764. [Google Scholar] [CrossRef]
- Kaczmarek, D.J.; Coyle, D.R.; Coleman, M.D. Survival and growth of a range of Populus clones in central South Carolina USA through age ten: Do early assessments reflect longer-term survival and growth trends. Biomass Bioenergy 2013, 49, 260–272. [Google Scholar] [CrossRef]
- Major, J.E.; Mosseler, A.; Malcolm, J.W.; Heartz, S. Salinity tolerance of three Salix, species: Survival, biomass yield and allocation, and biochemical efficiencies. Biomass Bioenergy 2017, 105, 10–22. [Google Scholar] [CrossRef]
- Dong, Y.; Yan, M.A.; Wang, H.; Zhang, J.; Zhang, G.; Yang, M.S. Assessment of tolerance of willows to saline soils through electrical impedance measurements. For. Ecosyst. 2013, 15, 32–40. [Google Scholar]
- Mamashita, T.; Larocque, G.R.; Desrochers, A.; Beaulieu, J.; Thomas, B.R.; Mosseler, A.; Major, J.; Sidders, D. Short-term growth and morphological responses to nitrogen availability and plant density in hybrid poplars and willows. Biomass Bioenergy 2015, 81, 88–97. [Google Scholar] [CrossRef]
- Galmés, J.; Flexas, J.; Savé, R.; Medrano, H. Water relations and stomatal characteristics of Mediterranean plants with different growth forms and leaf habits: Responses to water stress and recovery. Plant Soil 2007, 290, 139–155. [Google Scholar] [CrossRef]
- Chen, G.C.; Liu, Z.; Zhang, J.; Owens, G. Phytoaccumulation of copper in willow seedlings under different hydrological regimes. Ecol. Eng. 2012, 44, 285–289. [Google Scholar] [CrossRef]
- Cao, Y.; Ma, C.; Chen, G.; Zhang, J.; Xing, B. Physiological and biochemical responses of Salix integrathunb. under copper stress as affected by soil flooding. Environ. Pollut. 2017, 225, 644–653. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Samarakoon, M.B.; Yagisawa, J. Effects of root architecture, physical tree characteristics, and soil shear strength on maximum resistive bending moment for overturning Salix babylonica, and Juglans ailanthifolia. Landsc. Ecol. Eng. 2012, 8, 69–79. [Google Scholar] [CrossRef]
- Davies, W.J.; Wilkinson, S.; Loveys, B. Stomatal control by chemical signalling and the exploitation of this mechanism to increase water use efficiency in agriculture. New Phytol. 2002, 153, 449–460. [Google Scholar] [CrossRef]
- Maud, V.; Smith, H.K.; David, C.; Jennifer, D.; Harriet, T.; Marijke, S.; Bastien, C.; Taylor, G. Adaptive mechanisms and genomic plasticity for drought tolerance identified in European black poplar (Populus nigral.). Tree Physiol. 2016, 36, 909–928. [Google Scholar]
- Wikberg, J.; Ogren, E. Variation in drought resistance, drought acclimation and water conservation in four willow cultivars used for biomass production. Tree Physiol. 2007, 27, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- Podlaski, S.; Pietkiewicz, S.; Chołuj, D.; Horaczek, T.; Wiśniewski, G.; Gozdowski, D.; Kalaji, H.M. The relationship between the soil water storage and water-use efficiency of seven energy crops. Photosynthetica 2017, 55, 210–218. [Google Scholar] [CrossRef]
- Landhaüsser, S.M.; Stadt, K.J.; Lieffers, V.J. Photosynthetic strategies of summer green and evergreen understory herbs of the boreal mixed wood forest. Oecologia 1997, 112, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, G.S.; Van Rees, K.C.J. Soil organic carbon sequestration by shelterbelt agroforestry systems in Saskatchewan. Can. J. Soil Sci. 2017, 97, 394–409. [Google Scholar] [CrossRef]
- Kattge, J.; Knorr, W.; Raddatz, T.; Wirth, C. Quantifying photosynthetic capacity and its relationship to leaf nitrogen content for global-scale terrestrial biosphere models. Glob. Chang. Biol. 2009, 15, 976–991. [Google Scholar] [CrossRef]
- Weih, M.; Bonosi, L.; Ghelardini, L. Optimizing nitrogen economy under drought: Increased leaf nitrogen is an acclimation to water stress in willow (Salix spp.). Ann. Bot. 2011, 108, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Feng, D.; Wang, Y.; Lu, T.; Zhang, Z.; Han, X. Proteomics analysis reveals a dynamic diurnal pattern of photosynthesis-related pathways in maize leaves. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Wang, L.; Zhou, Q.; Huang, X. Stomatal and non-stomatal factors regulated the photosynthesis of soybean seedlings in the present of exogenous bisphenol A. Ecotoxicol. Environ. Saf. 2017, 145, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Adams, W.W., III; Diaz, M.; Winter, K. Diurnal changes in photochemical efficiency, the reduction state of Q. radiationless energy dissipation, and non-photochemical fluorescence quenching in cacti exposed to natural sunlight in northern Venezuela. Oecologia 1989, 80, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Meng, Z.; Zhang, G.; Qi, M.; Sun, Z.; Liu, Y.; Li, T. Sub-high temperature and high light intensity induced irreversible inhibition on photosynthesis system of tomato plant (Solanum lycopersicum L.). Front. Plant Sci. 2017, 8, 182. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Pan, X.L.; Hong, L. Responses of photosystem II (PSII) function in leaves and samaras of Ulmus pumila to chilling and freezing temperatures and subsequent recovery. Int. J. Agric. Biol. 2012, 14, 739–744. [Google Scholar]
- Kaipiainen, E.L. Parameters of photosynthesis light curve in Salix dasyclados and their changes during the growth season. Russ. J. Plant Physiol. 2009, 56, 445–453. [Google Scholar] [CrossRef]
- Ögren, E.; Sjöström, M. Estimation of the effect of photoinhibition on the carbon gain in leaves of a willow canopy. Planta 1990, 181, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Zhao, C.Y.; Li, J.; Yan, Y.Y.; Yu, B.; Han, M. Growth and leaf gas exchange in Populus euphratica across soil water and salinity gradients. Photosynthetica 2013, 51, 321–329. [Google Scholar] [CrossRef]
- Cai, Z.Q. Shade delayed flowering and decreased photosynthesis, growth and yield of Sacha Inchi (Plukenetia volubilis) plants. Ind. Crop. Prod. 2011, 34, 1235–1237. [Google Scholar] [CrossRef]
- Ďurkovič, J.; Čaňová, I.; Priwitzer, T.; Biroščíková, M.; Kapraľ, P.; Saniga, M. Field assessment of photosynthetic characteristics in Micropropagated and grafted Wych elm (Ulmus glabra, huds.) trees. Plant Cell Tiss. Organ Cult. 2010, 101, 221–228. [Google Scholar] [CrossRef]
- Gong, J.R.; Zhao, A.F.; Huang, Y.M.; Zhang, X.S.; Zhang, C.L. Water relations, gas exchange, photochemical efficiency, and Peroxidative stress of four plant species in the Heihe drainage basin of Northern China. Photosynthetica 2006, 44, 355–364. [Google Scholar] [CrossRef]
- Xue, W.; Li, X. Moderate shade environment facilitates establishment of desert phreatophytic species Alhagi sparsifolia, seedlings by enlarge fine root biomass. Acta Physiol. Plant 2017, 39, 7. [Google Scholar] [CrossRef]
- Liu, M.Z.; Jiang, G.M.; Li, Y.G.; Niu, S.L.; Cui, H.X. Gas exchange, photochemical efficiency, and leaf water potential in three Salix species. Photosynthetica 2003, 1, 393–398. [Google Scholar] [CrossRef]
- Contin, D.R.; Soriani, H.H.; Hernández, I.; Furriel, R.P.M.; Munné-Bosch, S.; Martinez, C.A. Antioxidant and photoprotective defenses in response to gradual water stress under low and high irradiance in two Malvaceae tree species used for tropical forest restoration. Trees 2014, 28, 1705–1722. [Google Scholar] [CrossRef]
- Liu, J.; Last, R.L. A chloroplast thylakoid lumen protein is required for proper photosynthetic acclimation of plants under fluctuating light environments. Proc. Natl. Acad. Sci. USA 2017, 114, 8110–8117. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Ungerer, P.; Zhang, H.; Ruban, A.V. Direct impact of the sustained decline in the photosystem ii efficiency upon plant productivity at different developmental stages. J. Plant Physiol. 2017, 212, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Wang, L.; Zhang, X.; Korpelainen, H.; Li, C. Sexual differences in photosynthetic activity, ultrastructure and phytoremediation potential of Populus cathayana exposed to lead and drought. Tree Physiol. 2013, 33, 1043–1060. [Google Scholar] [CrossRef] [PubMed]
- Demmig–Adams, B.; Adams, W.W., III; Logan, B.A.; Verhoeven, A.S. Xanthophyll cycle dependent energy dissipation and flexible PSII efficiency in plants acclimated to light stress. Aust. J. Plant Physiol. 1995, 22, 249–261. [Google Scholar]
- Ding, L.; Wang, K.J.; Jiang, G.M.; Li, Y.G.; Jiang, C.D.; Liu, M.Z.; Niu, S.L.; Peng, Y. Diurnal variation of gas exchange, chlorophyll fluorescence, and xanthophyll cycle components of maize hybrids released in different years. Photosynthetica 2006, 44, 26–31. [Google Scholar] [CrossRef]
- Pierangelini, M.; Ryšánek, D.; Lang, I.; Adlassnig, W.; Holzinger, A. Terrestrial adaptation of green algae Klebsormidium and Zygnema (charophyta) involves diversity in photosynthetic traits but not in CO2 acquisition. Planta 2017, 246, 971–986. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.V. Quantifying the efficiency of photoprotection. Phil. Trans. R. Soc. B 2017, 372, 1730. [Google Scholar] [CrossRef] [PubMed]
- Randriamanana, T.R.; Nissinen, K.; Moilanen, J.; Nybakken, L.; Julkunen-Tiitto, R. Long-term UV-B and temperature enhancements suggest that females of Salix myrsinifolia, plants are more tolerant to UV-B than males. Environ. Exp. Bot. 2014, 109, 296–305. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, G.; Guerra, C.; Jesus, C.; Amaral, J.; Santos, C. Effect of irradiance during acclimatization on content of proline and phytohormones in micropropagated Ulmus minor. Biol. Plant. 2013, 57, 769–772. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Depth (cm) | |||
|---|---|---|---|---|
| 0–10 | 10–20 | 20–40 | 40–60 | |
| Populus cathayana | 2.78 | 1.94 | 2.32 | 1.38 |
| Ulmus pumila | 1.28 | 0.55 | 0.79 | 1.52 |
| Salix longistamina | 2.34 | 2.15 | 1.23 | 0.95 |
| Species | Survival Rates (%) | Crown Diameter (cm × cm) | Height (cm) | Total Biomass (g) |
|---|---|---|---|---|
| Populus cathayana | 83.83 ± 2.75 | 45.00 × 45.00 | 182.33 ± 3.18 | 381.41 ± 7.08 |
| Ulmus pumila | 93.80 ± 5.52 | 56.00 × 67.00 | 167.00 ± 5.77 | 209.61 ± 4.73 |
| Salix longistamina | 71.70 ± 3.06 | 78.00 × 63.00 | 122.75 ± 1.73 | 608.97 ± 29.94 |
| Species | Pmax (µmol CO2 m−2 s−1) | LSP (µmol m−2 s−1) | LCP (µmol m−2 s−1) | Rd (µmol m−2 s−1) | AQY (µmol CO2 µmol−1) |
|---|---|---|---|---|---|
| P. cathayana | 22.30 ± 2.14 | 1791.67 ± 33.29 | 39.89 ± 3.69 | 1.02 ± 0.09 | 0.0448 ± 0.0012 |
| S. longistamina | 12.80 ± 3.11 | 2290.00 ± 28.35 | 73.01 ± 4.54 | 3.31 ± 0.15 | 0.0454 ± 0.0014 |
| U. pumila | 14.90 ± 2.32 | 998.33 ± 13.58 | 26.35 ± 2.25 | 2.23 ± 0.12 | 0.0686 ± 0.0011 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, J.; Zhang, Z.; Zhang, C.; Zhang, J.; Ran, A. Ecophysiological Responses of Three Tree Species to a High-Altitude Environment in the Southeastern Tibetan Plateau. Forests 2018, 9, 48. https://doi.org/10.3390/f9020048
Gong J, Zhang Z, Zhang C, Zhang J, Ran A. Ecophysiological Responses of Three Tree Species to a High-Altitude Environment in the Southeastern Tibetan Plateau. Forests. 2018; 9(2):48. https://doi.org/10.3390/f9020048
Chicago/Turabian StyleGong, Jirui, Zihe Zhang, Chunlai Zhang, Jiaqiong Zhang, and An Ran. 2018. "Ecophysiological Responses of Three Tree Species to a High-Altitude Environment in the Southeastern Tibetan Plateau" Forests 9, no. 2: 48. https://doi.org/10.3390/f9020048
APA StyleGong, J., Zhang, Z., Zhang, C., Zhang, J., & Ran, A. (2018). Ecophysiological Responses of Three Tree Species to a High-Altitude Environment in the Southeastern Tibetan Plateau. Forests, 9(2), 48. https://doi.org/10.3390/f9020048