Encroachment Dynamics of Juniperus virginiana L. and Mesic Hardwood Species into Cross Timbers Forests of North-Central Oklahoma, USA




Abstract
:1. Introduction
2. Materials and Methods
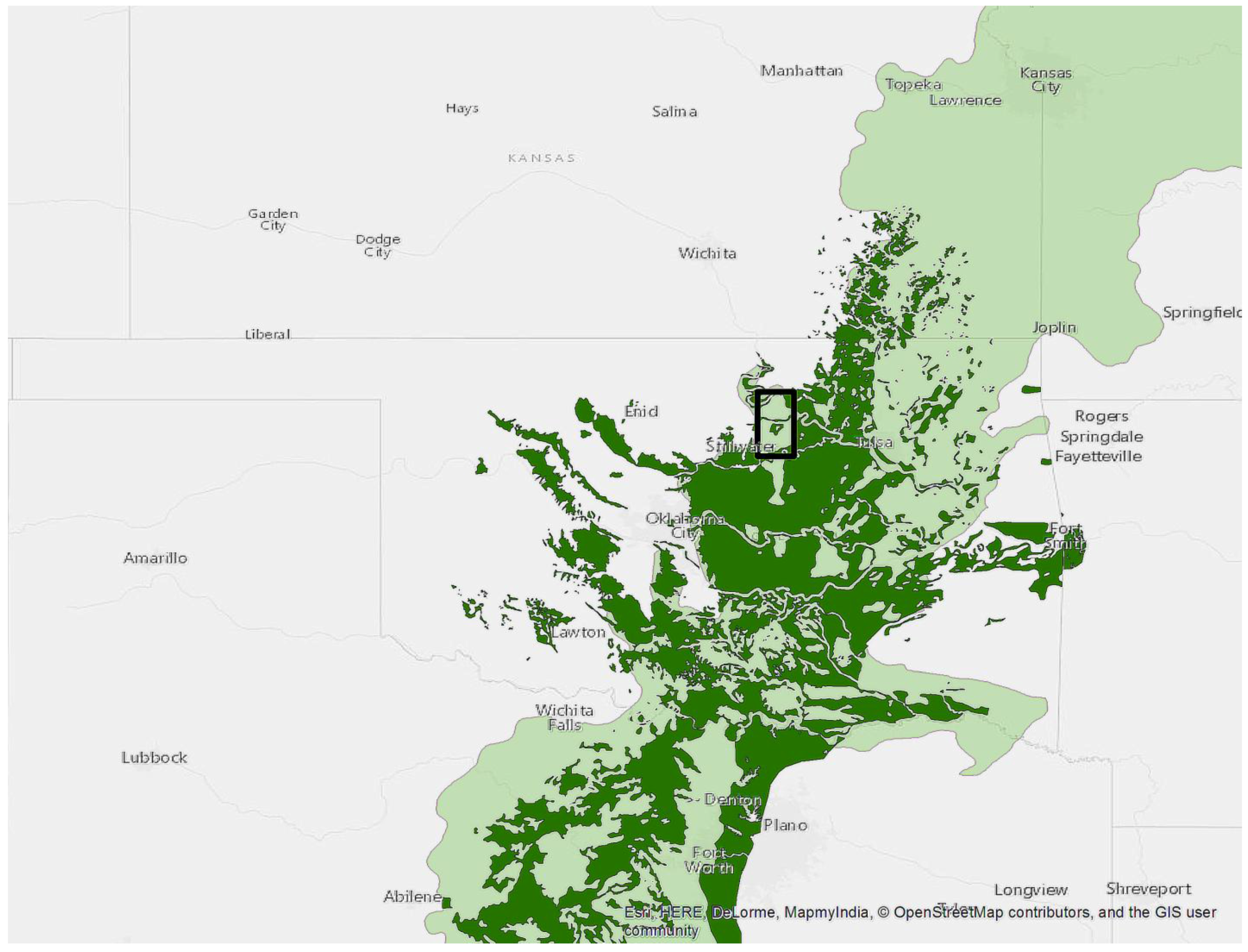
2.1. Study Area
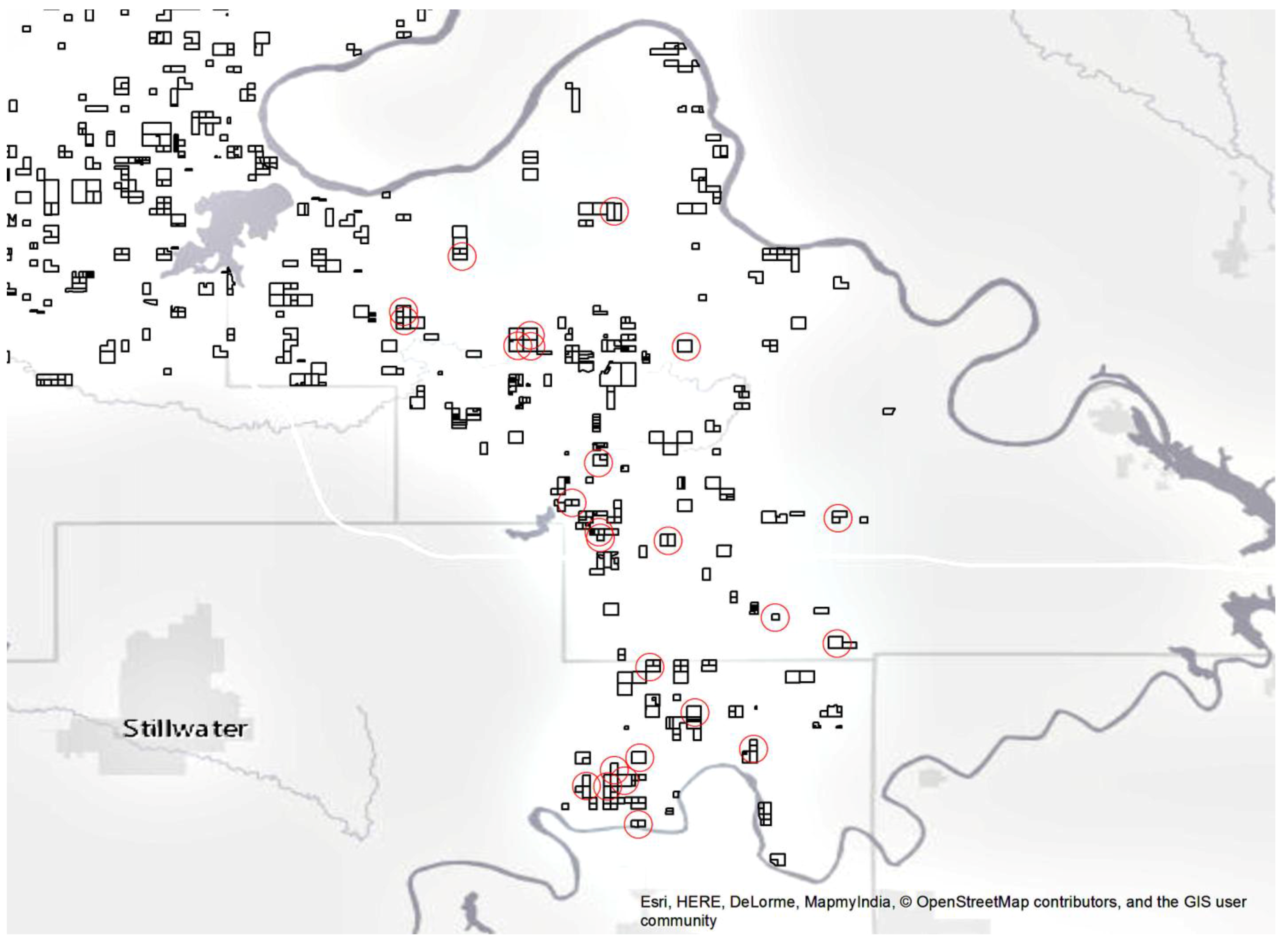
2.2. Data Collection
2.3. Age
2.4. Analyses
3. Results
3.1. Forest Composition
3.2. Relative Dominance and Density
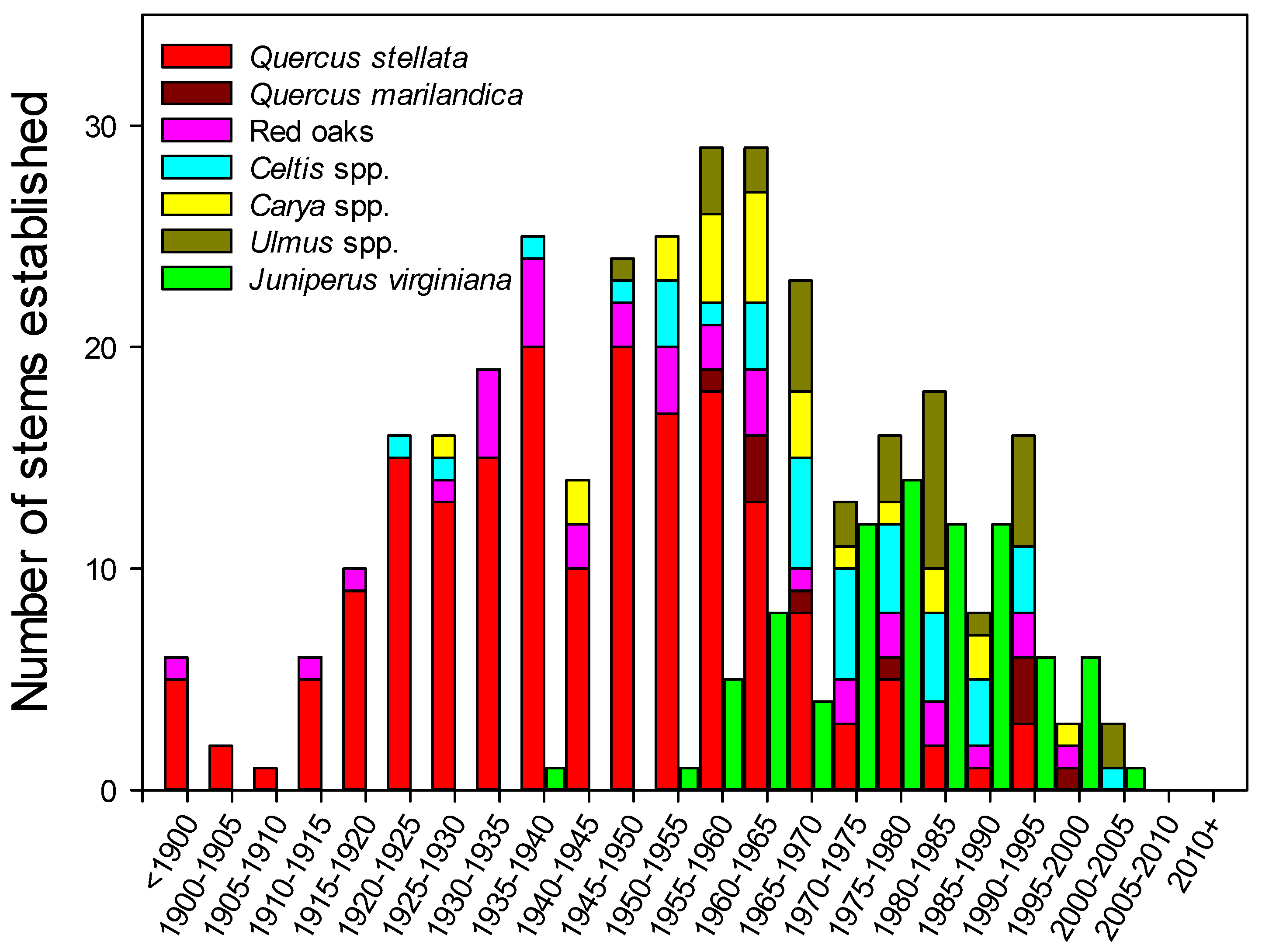
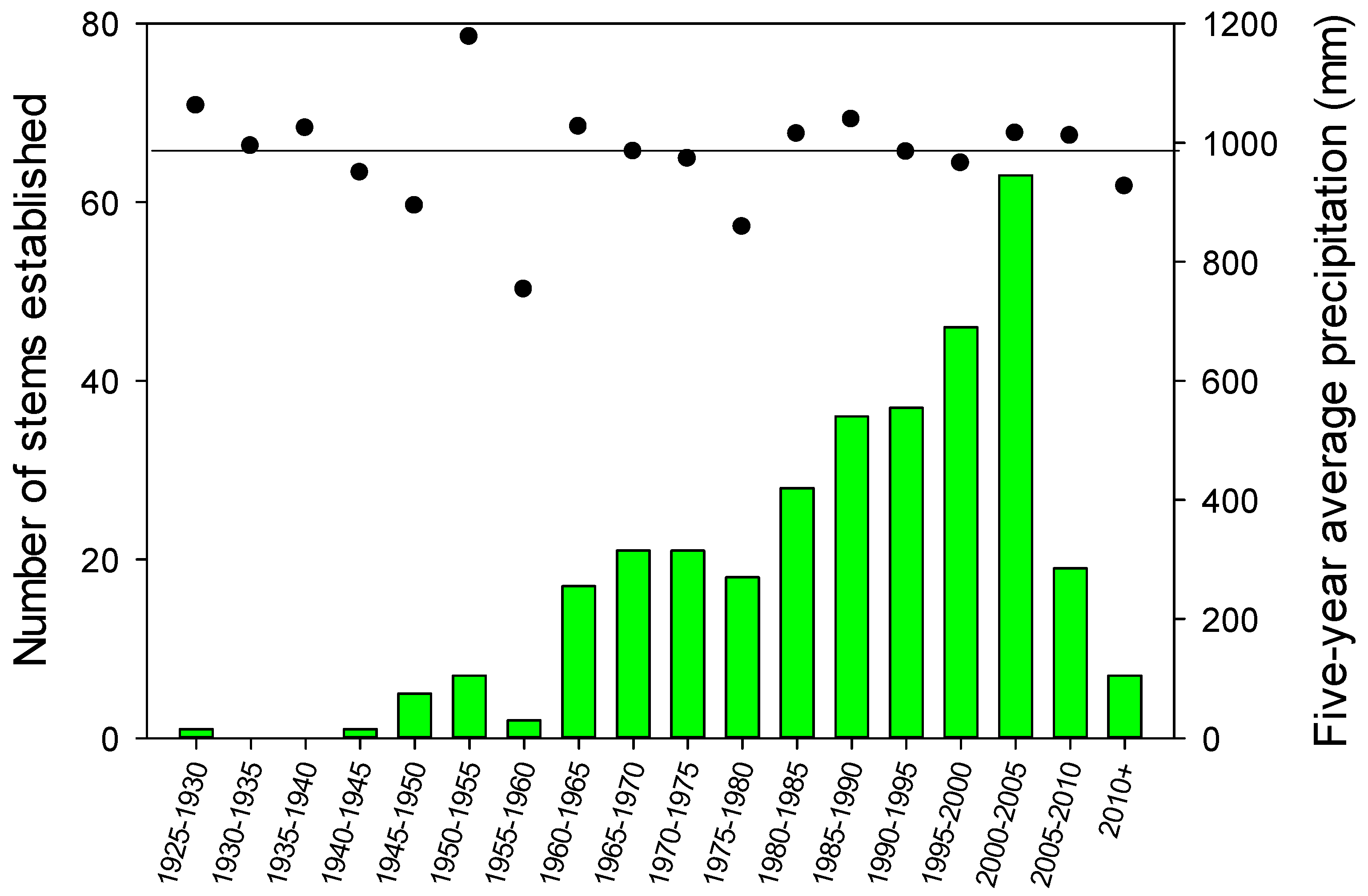
3.3. Age Distribution
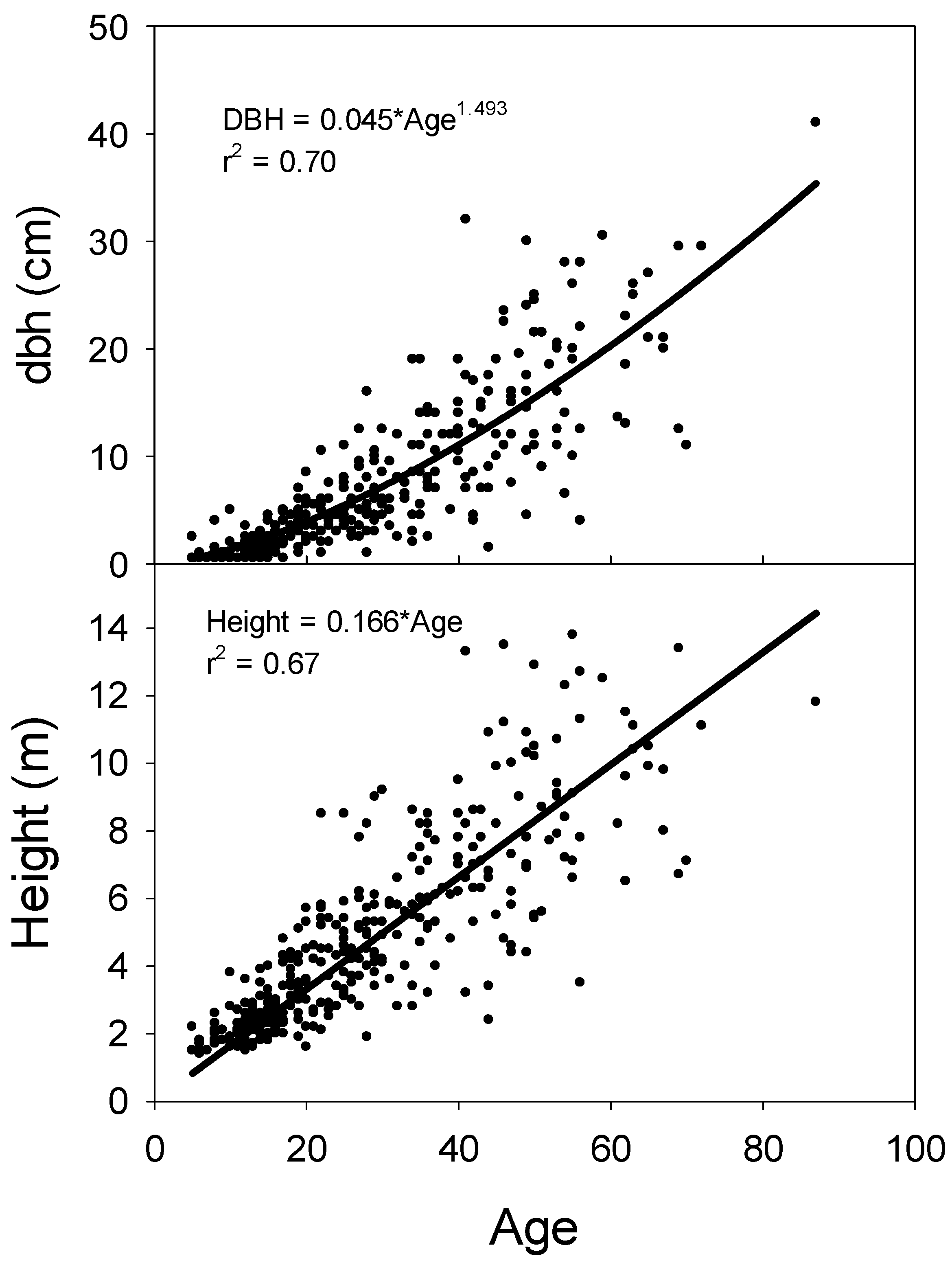
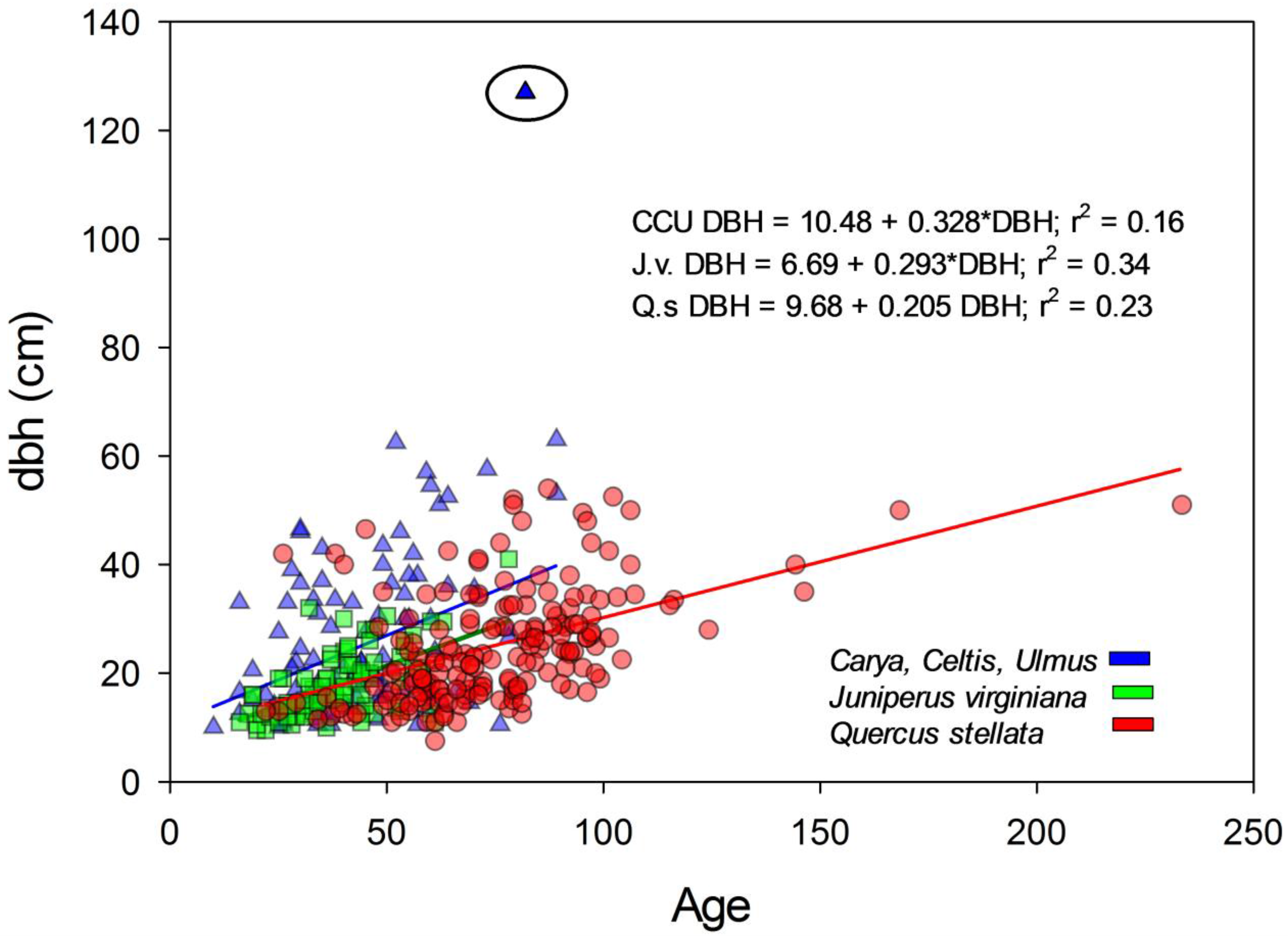
3.4. Age Size Relationships between Groups
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kuchler, A.W. Potential Natural Vegetation of the Conterminous United States; Special Publication No. 36; American Geographical Society: New York, NY, USA, 1964. [Google Scholar]
- Therrell, M.D.; Stahle, D.W. A predictive model to locate ancient forests in the Cross Timbers of Osage County, Oklahoma. J. Biogeogr. 1998, 25, 847–854. [Google Scholar] [CrossRef]
- Hallgren, S.W.; DeSantis, R.D.; Burton, J.A. Fire and vegetation dynamics in the Cross Timbers forests of South-Central North America. In Proceedings of the 4th Fire in Eastern Oak Forests Conference, Springfield, MO, USA, 17–19 May 2011; Dey, D.C., Stambaugh, M.C., Clark, S.L., Schweitzer, C.J., Eds.; USDA Forest Service, Northern Research Station: Newtown Square, PA, USA, 2012. [Google Scholar]
- Clark, S.L.; Hallgren, S.W.; Stahle, D.W.; Lynch, T.B. Characteristics of the Keystone Ancient Forest Preserve, an old-growth forest in the Cross Timbers of Oklahoma. Nat. Area J. 2005, 25, 165–175. [Google Scholar]
- Stambaugh, M.C.; Guyette, R.P.; Godfrey, R.; McMurry, E.R.; Marschall, J.M. Fire, drought, and human history near the Western terminus of the Cross Timbers, Wichita Mountains, Oklahoma, USA. Fire Ecol. 2009, 5, 51–65. [Google Scholar] [CrossRef]
- DeSantis, R.D.; Hallgren, S.W.; Stahle, D.W. Historic fire regime of an upland oak forest in south-central North America. Fire Ecol. 2010, 6, 45–61. [Google Scholar] [CrossRef]
- Allen, M.S.; Palmer, M.W. Fire history of a prairie/forest boundary: More than 250 years of frequent fire in a North American tallgrass prairie. J. Veg. Sci. 2011, 22, 436–444. [Google Scholar] [CrossRef]
- Stambaugh, M.C.; Sparks, J.C.; Guyette, R.P.; Willson, G. Fire history of a relict oak woodland in northeast Texas. Rangel. Ecol. Manag. 2011, 64, 419–423. [Google Scholar] [CrossRef]
- DeSantis, R.D.; Hallgren, S.W.; Stahle, D.W. Drought and fire suppression lead to rapid forest composition change in a forest-prairie ecotone. For. Ecol. Manag. 2011, 261, 1183–1840. [Google Scholar] [CrossRef]
- Nowacki, G.J.; Abrams, M.D. The demise of fire and “mesophication” of forests in the Eastern United States. BioScience 2008, 58, 123–138. [Google Scholar] [CrossRef]
- Rice, E.L.; Penfound, W.T. The upland forests of Oklahoma. Ecology 1959, 40, 593–608. [Google Scholar] [CrossRef]
- DeSantis, R.D.; Hallgren, S.W.; Lynch, T.B.; Burton, J.A.; Palmer, M.W. Long-term directional changes in upland Quercus forests throughout Oklahoma, USA. J. Veg. Sci. 2010, 21, 606–615. [Google Scholar] [CrossRef]
- Caterina, G.L.; Will, R.E.; Turton, D.J.; Wilson, D.S.; Zou, C.B. Water use of Juniperus virginiana trees encroached into mesic prairies in Oklahoma, USA. Ecohydrology 2014, 7, 1124–1134. [Google Scholar]
- Van Els, P.; Will, R.E.; Palmer, M.W.; Hickman, K.R. Changes in forest understory associated with Juniperus encroachment in Oklahoma, USA. Appl. Veg. Sci. 2010, 13, 356–368. [Google Scholar]
- Nunes-Biral, V.C. Impacts of Tree Encroachment into a Grassland on Soil Carbon and Inputs in Northcentral Oklahoma. Master’s Thesis, Oklahoma State University, Stillwater, OK, USA, 2017. [Google Scholar]
- Williams, R.J.; Hallgren, S.W.; Wilson, G.W.T.; Palmer, M.W. Juniperus virginiana encroachment into upland oak forests alters arbuscular mycorrhizal abundance and litter chemistry. Appl. Soil Ecol. 2013, 65, 23–30. [Google Scholar] [CrossRef]
- Ansley, R.J.; Wiedemann, H.T. Reversing the woodland steady state: Vegetation responses during restoration of Juniperus-dominated grasslands with chaining and fire. In Western North American Juniperus Communities: A Dynamic Vegetation Type; Van Auken, O.W., Ed.; Springer: New York, NY, USA, 2008; pp. 272–290. [Google Scholar]
- Owensby, C.E.; Blan, K.R.; Eaton, B.J.; Russ, O.G. Evaluation of eastern redcedar infestations in the Northern Kansas Flint Hills. J. Range Manag. 1973, 26, 256–260. [Google Scholar] [CrossRef]
- Twidwell, D.; Fuhlendorf, S.D.; Taylor, C.A., Jr.; Rogers, W.E. Refining thresholds in coupled fire-vegetation models to improve management of encroaching woody plants in grasslands. J. Appl. Ecol. 2013, 50, 603–613. [Google Scholar] [CrossRef]
- Hoff, D.; Will, R.E.; Zou, C.B.; Weir, J.; Gregory, M.; Lillie, N. Estimating increased fuel loading within the Cross Timbers forest matrix of Oklahoma, USA due to an encroaching conifer, Juniperus virginiana, using leaf-off imagery. For. Ecol. Manag. 2018, 409, 215–224. [Google Scholar] [CrossRef]
- Stambaugh, M.C.; Marschall, J.M.; Guyette, R.P. Linking fire history to successional changes of xeric oak woodlands. For. Ecol. Manag. 2014, 320, 83–95. [Google Scholar] [CrossRef]
- Bragg, D.C.; Stahle, D.W.; Cerny, K.C. Structural attributes of two old-growth Cross Timbers stands in western Arkansas. Am. Midl. Nat. 2012, 167, 40–55. [Google Scholar] [CrossRef]
- Scott, G.F.; Boyett, J.B.; Dennis, J.; White, J.D. Supplement to the Soil Survey of Pawnee County, Oklahoma; USDA Natural Resource Conservation Service: Washington, DC, USA, 2006. [Google Scholar]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’Amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the world: A new map of life on earth. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Lutes, D.C.; Keane, R.E.; Caratti, J.F.; Key, C.H.; Benson, N.C.; Sutherland, S.; Gangi, L.J. FIREMON: Fire Effects Monitoring and Inventory System; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2006. [Google Scholar]
- Southern Climate Impacts Planning Program. Climate Trends for Oklahoma North-Central Region. Available online: http://charts.srcc.lsu.edu/trends/ (accessed on 2 October 2017).
- Clark, S.L.; Hallgren, S.W. Dynamics of oak (Quercus marilandica and Q. stellata) reproduction in an old-growth Cross Timbers forest. Southeast. Nat. 2003, 2, 559–574. [Google Scholar]
- Lykin, R.W. Estimation of Aboveground Eastern Redcedar Biomass. Master’s Thesis, Oklahoma State University, Stillwater, OK, USA, 1995. [Google Scholar]
- Despres-Loustau, M.L.; Marcais, B.; Nageleisen, L.M.; Piou, D.; Vannini, A. Interactive effects of drought and pathogens in forest trees. Ann. For. Sci. 2006, 63, 597–612. [Google Scholar] [CrossRef]
- Oklahoma Climatological Survey. The Climate of Pawnee County. Available online: http://climate.mesonet.org/county_climate/Products/County_Climatologies/county_climate_pawnee.pdf (accessed on 26 May 2017).
- Masters, R.E.; Waymire, J.R. Oak savanna restoration: Oak response to fire and thinning through 28 years. In Proceedings of the 4th Fire in Eastern Oak Forests Conference, Springfield, MO, USA, 17–19 May 2011; Dey, D.C., Stambaugh, M.C., Clark, S.L., Schweitzer, C.J., Eds.; USDA Forest Service, Northern Research Station: Newtown Square, PA, USA, 2012. [Google Scholar]
- Sperry, J.S.; Tyree, M.T. Water-stress-induced xylem embolism in three species of conifers. Plant Cell Environ. 1990, 13, 427–436. [Google Scholar] [CrossRef]
- Willson, C.J.; Manos, P.S.; Jackson, R.B. Hydraulic traits are influenced by phylogenetic history in the drought-resistant, invasive genus Juniperus (Cupressaceae). Am. J. Bot. 2008, 95, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Volder, A.; Tjoelker, M.G.; Briske, D.D. Contrasting physiological responsiveness of establishing trees and a C4 grass to rainfall events, intensified summer drought, and warming in oak savanna. Glob. Chang. Biol. 2010, 16, 3349–3362. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Overstory Trees | Saplings | Seedlings | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| dbh (cm) | Height (m) | dbh (cm) | Height (m) | Height (m) | |||||||||
| Genus Species | N | Mean | Std. | Mean | Std. | N | Mean | Std. | Mean | Std. | N | Mean | Std. |
| Acer negundo L. | 2 | 21.0 | 14.1 | - | - | 5 | 4.6 | 2.6 | 4.9 | 1.7 | 0 | - | - |
| Carya illinoinensis (Wangenh.) K.Koch | 37 | 27.8 | 14.1 | 13.3 | 5.5 | 4 | 4.5 | 3.0 | 5.2 | 2.8 | 8 | 0.4 | 0.2 |
| Carya texana Buckley | 90 | 18.3 | 8.5 | 11.9 | 4.4 | 36 | 4.3 | 2.8 | 4.2 | 2.3 | 62 | 0.4 | 0.3 |
| Celtis laevigata Willdenow | 50 | 19.1 | 9.7 | 11.1 | 4.5 | 21 | 2.8 | 2.0 | 3.6 | 1.9 | 0 | - | - |
| Celtis occidentalis L. | 164 | 20.8 | 14.1 | 11.3 | 4.7 | 141 | 2.4 | 2.1 | 3.1 | 1.6 | 338 | 0.4 | 0.3 |
| Cercis canadensis L. | 4 | 10.9 | 1.0 | - | - | 107 | 2.4 | 2.1 | 2.9 | 1.2 | 69 | 0.5 | 0.3 |
| Cornus drummondii C.A.Mey. | 0 | - | - | - | - | 149 | 1.1 | 0.7 | 2.0 | 0.9 | 158 | 0.8 | 0.4 |
| Diospyros virginiana L. | 10 | 18.0 | 7.7 | 10.5 | 5.1 | 8 | 4.0 | 2.4 | 4.2 | 1.5 | 8 | 0.6 | 0.3 |
| Fraxinus spp. L. | 36 | 28.6 | 13.8 | 15.7 | 4.6 | 6 | 3.0 | 2.5 | 3.5 | 2.1 | 5 | 0.8 | 0.3 |
| Gleditsia triacanthos L. | 1 | 12.5 | - | - | - | 1 | 3.0 | - | 4.4 | - | 3 | 0.5 | 0.1 |
| Gymnocladus dioicus (L.) K. Koch | 6 | 21.8 | 22.2 | 12.9 | 5.5 | 0 | - | - | - | - | 0 | - | - |
| Juglans nigra L. | 23 | 23.8 | 16.0 | 10.3 | 4.9 | 3 | 6.3 | 3.1 | 4.7 | 1.7 | 0 | - | - |
| Juniperus virginiana L. | 296 | 16.4 | 6.2 | 8.6 | 2.1 | 1210 | 3.0 | 2.4 | 3.2 | 1.5 | 891 | 0.7 | 0.4 |
| Morus rubra L. | 29 | 19.5 | 9.4 | 12.0 | 3.7 | 0 | - | - | - | - | 5 | 0.5 | 0.2 |
| Platanus occidentalis L. | 11 | 36.7 | 13.9 | 18.7 | 0.1 | 0 | - | - | - | - | 0 | - | - |
| Prunus angustifolia Marsh. | 2 | 11.0 | 1.4 | - | - | 0 | - | - | - | - | 0 | - | - |
| Prunus mexicana S.Watson | 8 | 12.9 | 2.4 | 6.4 | 2.5 | 26 | 1.9 | 1.7 | 4.6 | 1.9 | 54 | 0.8 | 0.4 |
| Prunus serotina Ehrh. | 0 | - | - | - | - | 0 | - | - | - | - | 2 | 0.4 | 0.0 |
| Quercus macrocarpa Michx. | 10 | 24.2 | 15.1 | 10.7 | 2.1 | 0 | - | - | - | - | 2 | 0.4 | 0.0 |
| Quercus marilandica Muenchh. | 82 | 21.1 | 9.3 | 10.7 | 2.7 | 9 | 5.2 | 3.1 | 2.4 | 0.9 | 30 | 0.2 | 0.2 |
| Quercus muehlenbergii Engelm. | 18 | 27.0 | 13.8 | 10.3 | 2.8 | 11 | 2.5 | 2.4 | 4.4 | 1.8 | 57 | 0.3 | 0.3 |
| Quercus shumardii Buckland | 2 | 31.0 | 0.7 | 13.1 | - | 0 | - | - | - | - | 0 | - | - |
| Quercus stellata Wangenh. | 888 | 22.9 | 9.3 | 12.0 | 3.2 | 23 | 4.7 | 3.3 | 3.3 | 2.3 | 157 | 0.3 | 0.2 |
| Quercus velutina Lam. | 137 | 31.7 | 16.2 | 16.3 | 4.3 | 16 | 2.9 | 2.6 | 1.8 | 0.5 | 161 | 0.3 | 0.3 |
| Rhus copallina L. | 0 | - | - | - | - | 4 | 1.0 | 0.0 | 2.1 | 0.4 | 18 | 0.4 | 0.3 |
| Robinia pseudoacacia L. | 0 | - | - | - | - | 4 | 1.5 | 1.0 | 3.2 | 1.5 | 2 | 1.0 | 0.2 |
| Salix nigra Marshall | 3 | 37.5 | 5.8 | 12.4 | - | 0 | - | - | - | - | 0 | - | - |
| Sapindus drummondii L. | 14 | 15.9 | 4.1 | 9.3 | 1.9 | 35 | 2.3 | 1.8 | 3.6 | 1.6 | 31 | 0.3 | 0.3 |
| Sideroxylon languinosum Michx. | 41 | 17.8 | 6.0 | 9.5 | 2.5 | 9 | 2.1 | 2.3 | 2.8 | 1.1 | 82 | 0.4 | 0.3 |
| Ulmus americana L. | 150 | 18.8 | 10.7 | 11.0 | 3.5 | 27 | 3.5 | 2.9 | 3.8 | 1.4 | 433 | 0.3 | 0.2 |
| Ulmus rubra Muhl. | 3 | 34.2 | 26.5 | 20.1 | - | 4 | 3.0 | 2.3 | 2.8 | 0.2 | 6 | 0.1 | 0.0 |
| Viburnum rufidulum Raf. | 0 | - | - | - | - | 3 | 1.7 | 1.2 | 1.6 | 0.1 | 3 | 0.3 | 0.1 |
| Zanthoxylum americanum Mill. | 0 | - | - | - | - | 10 | 1.0 | 0.0 | 2.9 | 1.6 | 0 | - | - |
| Overstory Trees | Saplings | Seedlings | ||||||
|---|---|---|---|---|---|---|---|---|
| BA (m2 ha−1) | Stems (ha−1) | Stems (ha−1) | Stems (ha−1) | |||||
| Genus Species | Mean | Std. | Mean | Std. | Mean | Std. | Mean | Std. |
| Acer negundo | 0.0 | 0.2 | 0.4 | 3.1 | 9.6 | 8.1 | 0.0 | 0.0 |
| Carya illinoinensis | 0.5 | 2.2 | 7.1 | 25.0 | 7.7 | 6.9 | 61.5 | 7.5 |
| Carya texana | 0.6 | 1.9 | 17.3 | 41.6 | 67.3 | 23.4 | 476.9 | 33.9 |
| Celtis laevigata | 0.3 | 1.1 | 9.6 | 27.0 | 40.4 | 18.5 | 0.0 | 0.0 |
| Celtis occidentalis | 1.6 | 4.1 | 31.5 | 68.8 | 271.2 | 69.9 | 2600.0 | 111.6 |
| Cercis canadensis | 0.0 | 0.1 | 0.8 | 5.3 | 205.8 | 109.4 | 530.8 | 40.1 |
| Cornus drummondii | 0.0 | 0.0 | 0.0 | 0.0 | 325.0 | 140.2 | 1215.4 | 164.7 |
| Diospyros virginiana | 0.1 | 0.5 | 1.9 | 14.1 | 15.4 | 13.3 | 61.5 | 7.5 |
| Fraxinus spp. | 0.6 | 2.2 | 6.9 | 25.3 | 11.5 | 6.8 | 38.5 | 6.5 |
| Gleditsia triacanthos | 0.0 | 0.0 | 0.2 | 2.2 | 1.9 | 2.4 | 23.1 | 6.6 |
| Gymnocladus dioicus | 0.1 | 0.8 | 1.2 | 8.2 | 0.0 | 0.0 | 0.0 | 0.0 |
| Juglans nigra | 0.3 | 1.6 | 4.4 | 15.7 | 5.8 | 4.2 | 0.0 | 0.0 |
| Juniperus virginiana | 1.3 | 3.3 | 56.5 | 117.2 | 232.7 | 429.1 | 171.4 | 484.0 |
| Morus rubra | 0.2 | 0.7 | 5.6 | 16.0 | 0.0 | 0.0 | 38.5 | 5.7 |
| Platanus occidentalis | 0.3 | 2.0 | 2.1 | 19.9 | 0.0 | 0.0 | 0.0 | 0.0 |
| Prunus angustifolia | 0.0 | 0.0 | 0.4 | 4.4 | 0.0 | 0.0 | 0.0 | 0.0 |
| Prunus mexicana | 0.0 | 0.1 | 1.5 | 6.0 | 50.0 | 19.2 | 415.4 | 39.4 |
| Prunus serotina | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 15.4 | 3.1 |
| Quercus macrocarpa | 0.1 | 0.7 | 1.9 | 9.7 | 0.0 | 0.0 | 15.4 | 4.4 |
| Quercus marilandica | 0.7 | 2.4 | 15.8 | 48.3 | 17.3 | 11.0 | 230.8 | 17.5 |
| Quercus muehlenbergii | 0.3 | 1.0 | 3.5 | 10.2 | 21.2 | 12.9 | 438.5 | 36.4 |
| Quercus shumardii | 0.0 | 0.2 | 0.4 | 3.1 | 0.0 | 0.0 | 0.0 | 0.0 |
| Quercus stellata | 8.2 | 8.1 | 170.8 | 174.4 | 44.2 | 21.4 | 1207.7 | 71.3 |
| Quercus velutina | 2.6 | 5.3 | 26.4 | 45.3 | 30.8 | 11.9 | 1238.5 | 69.2 |
| Rhus copallina | 0.0 | 0.0 | 0.0 | 0.0 | 7.7 | 9.8 | 138.5 | 17.5 |
| Robinia pseudoacacia | 0.0 | 0.0 | 0.0 | 0.0 | 7.7 | 7.7 | 15.4 | 3.1 |
| Salix nigra | 0.1 | 0.7 | 0.6 | 6.6 | 0.0 | 0.0 | 0.0 | 0.0 |
| Sapindus drummondii | 0.1 | 0.5 | 2.7 | 21.6 | 67.3 | 45.0 | 238.5 | 41.6 |
| Sideroxylon lanuginosum | 0.2 | 0.7 | 7.9 | 21.8 | 17.3 | 7.0 | 630.8 | 29.1 |
| Ulmus americana | 1.1 | 2.7 | 28.9 | 64.7 | 51.9 | 20.2 | 3330.8 | 198.4 |
| Ulmus rubra | 0.1 | 0.7 | 0.6 | 3.8 | 7.7 | 5.9 | 46.2 | 9.3 |
| Viburnum rufidulum | 0.0 | 0.0 | 0.0 | 0.0 | 5.8 | 4.2 | 23.1 | 3.8 |
| Zanthoxylum americanum | 0.0 | 0.0 | 0.0 | 0.0 | 19.2 | 24.4 | 0.0 | 0.0 |
| Total | 19.0 | - | 407.0 | - | 1542.0 | - | 13,202.0 | - |
| Overstory | Sapling | Seedling | ||||
|---|---|---|---|---|---|---|
| Genus Species | Prop | Plot | Prop | Plot | Prop | Plot |
| Acer negundo | 8 | 2 | 8 | 2 | 0 | 0 |
| Carya illinoinensis | 24 | 10 | 8 | 2 | 24 | 5 |
| Carya texana | 52 | 22 | 40 | 16 | 60 | 21 |
| Celtis laevigata | 60 | 22 | 28 | 8 | 0 | 0 |
| Celtis occidentalis | 68 | 35 | 56 | 30 | 92 | 61 |
| Cercis canadensis | 8 | 2 | 56 | 27 | 48 | 17 |
| Cornus drummondii | 0 | 0 | 48 | 18 | 60 | 18 |
| Diospyros virginiana | 12 | 3 | 8 | 2 | 24 | 5 |
| Fraxinus spp. | 40 | 12 | 8 | 4 | 4 | 2 |
| Gleditsia triacanthos | 4 | 1 | 4 | 1 | 4 | 1 |
| Gymnocladus dioicus | 12 | 2 | 0 | 0 | 0 | 0 |
| Juglans nigra | 28 | 9 | 8 | 2 | 0 | 0 |
| Juniperus virginiana | 96 | 53 | 100 | 84 | 92 | 46 |
| Morus rubra | 52 | 14 | 0 | 0 | 16 | 3 |
| Platanus occidentalis | 12 | 2 | 0 | 0 | 0 | 0 |
| Prunus angustifolia | 4 | 1 | 0 | 0 | 0 | 0 |
| Prunus mexicana | 28 | 6 | 24 | 10 | 48 | 16 |
| Prunus serotina | 0 | 0 | 0 | 0 | 4 | 2 |
| Quercus macrocarpa | 16 | 5 | 0 | 0 | 4 | 1 |
| Quercus marilandica | 52 | 21 | 12 | 4 | 44 | 14 |
| Quercus muehlenbergii | 44 | 12 | 12 | 4 | 44 | 17 |
| Quercus shumardii | 8 | 2 | 0 | 0 | 0 | 0 |
| Quercus stellata | 92 | 78 | 20 | 6 | 88 | 34 |
| Quercus velutina | 72 | 35 | 36 | 10 | 68 | 33 |
| Rhus copallina | 0 | 0 | 4 | 1 | 24 | 6 |
| Robinia pseudoacacia | 0 | 0 | 4 | 2 | 8 | 2 |
| Salix nigra | 4 | 1 | 0 | 0 | 0 | 0 |
| Sapindus drummondii | 8 | 2 | 16 | 4 | 16 | 5 |
| Sideroxylon lanuginosum | 60 | 18 | 28 | 7 | 88 | 34 |
| Ulmus americana | 88 | 38 | 28 | 12 | 100 | 61 |
| Ulmus rubra | 8 | 2 | 8 | 2 | 8 | 2 |
| Viburnum rufidulum | 0 | 0 | 8 | 2 | 12 | 2 |
| Zanthoxylum americanum | 0 | 0 | 4 | 1 | 0 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoff, D.L.; Will, R.E.; Zou, C.B.; Lillie, N.D. Encroachment Dynamics of Juniperus virginiana L. and Mesic Hardwood Species into Cross Timbers Forests of North-Central Oklahoma, USA. Forests 2018, 9, 75. https://doi.org/10.3390/f9020075
Hoff DL, Will RE, Zou CB, Lillie ND. Encroachment Dynamics of Juniperus virginiana L. and Mesic Hardwood Species into Cross Timbers Forests of North-Central Oklahoma, USA. Forests. 2018; 9(2):75. https://doi.org/10.3390/f9020075
Chicago/Turabian StyleHoff, Daniel L., Rodney E. Will, Chris B. Zou, and Nathan D. Lillie. 2018. "Encroachment Dynamics of Juniperus virginiana L. and Mesic Hardwood Species into Cross Timbers Forests of North-Central Oklahoma, USA" Forests 9, no. 2: 75. https://doi.org/10.3390/f9020075
APA StyleHoff, D. L., Will, R. E., Zou, C. B., & Lillie, N. D. (2018). Encroachment Dynamics of Juniperus virginiana L. and Mesic Hardwood Species into Cross Timbers Forests of North-Central Oklahoma, USA. Forests, 9(2), 75. https://doi.org/10.3390/f9020075