Drought Effects on Photosynthesis and Implications of Photoassimilate Distribution in 11C-Labeled Leaves in the African Tropical Tree Species Maesopsis eminii Engl.


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Set-Up
2.3. Photosynthesis Measurements
2.4. Light Response Curves
2.5. 11CO2 Leaf Labeling and Positron Autoradiography
2.6. Stable δ 13C Isotope Analysis
2.7. Vein Anatomy
2.8. Data Analysis and Statistics
3. Results
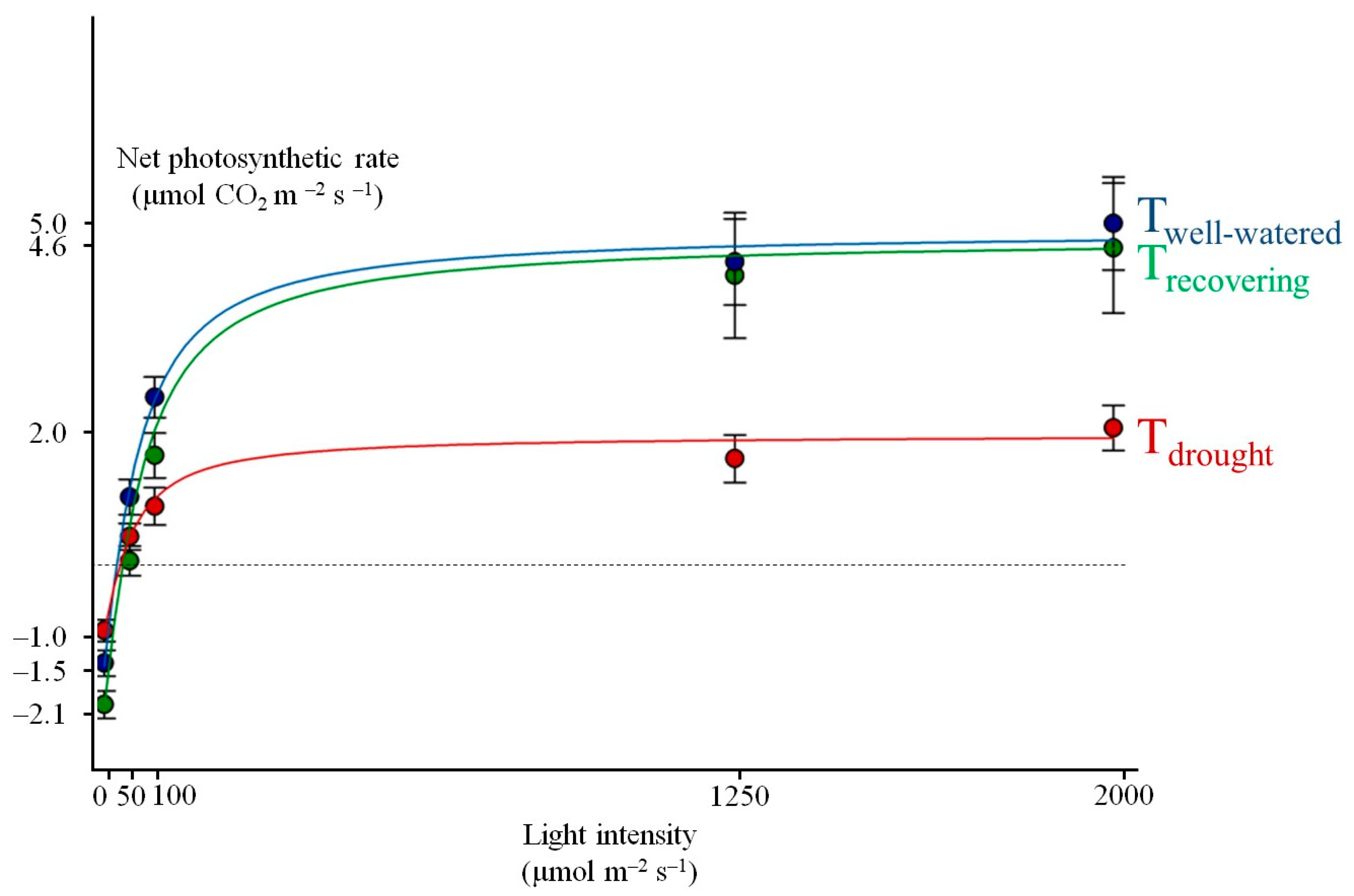
3.1. M. eminii’s Photosynthetic Performance
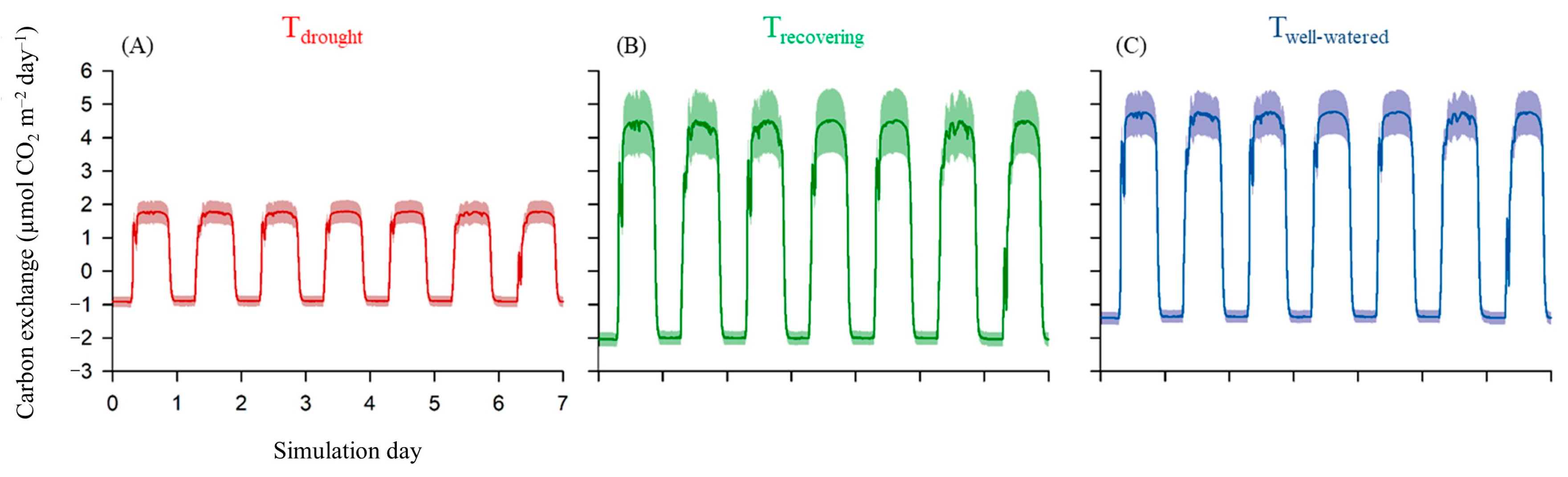
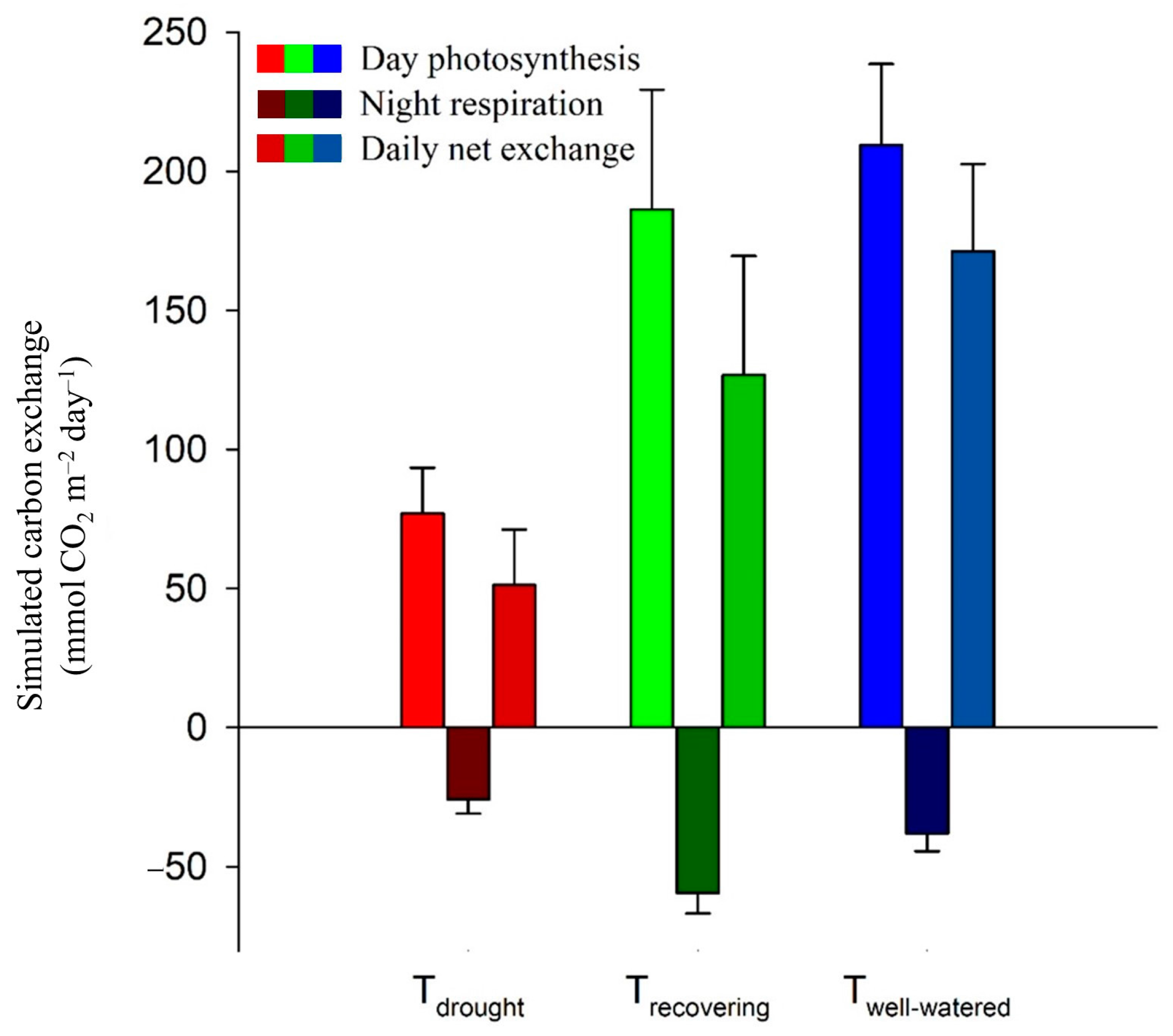
3.2. Estimated Diel Carbon Fluxes and Leaf Carbon Balances
3.3. 11C Autoradiographs and 13C Stable Isotope
3.4. Anatomy of the Main, Secondary and Tertiary Veins
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- IPCC. Summary for Policymakers. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 3–29. [Google Scholar]
- Fauset, S.; Baker, T.R.; Lewis, S.L.; Feldpausch, T.R.; Affum-Baffoe, K.; Foli, E.G.; Hamer, K.C.; Swaine, M.D. Drought-induced shifts in the floristic and functional composition of tropical forests in Ghana. Ecol. Lett. 2012, 15, 1120–1129. [Google Scholar] [CrossRef] [PubMed]
- Asefi-Najafabady, S.; Saatchi, S. Response of African humid tropical forests to recent rainfall anomalies. Philos. Trans. R. Soc. B 2013, 368, 20120306. [Google Scholar] [CrossRef] [PubMed]
- James, R.; Washington, R.; Rowell, D.P. Implications of global warming for the climate of african rainforests. Philos. Trans. R. Soc. B 2013, 368, 20120298. [Google Scholar] [CrossRef] [PubMed]
- Malhi, Y.; Adu-Bredu, S.; Asare, R.A.; Lewis, S.L.; Mayaux, P. African rainforests: Past, present and future. Philos. Trans. R. Soc. B 2013, 368, 20120312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oslisly, R.; White, L.; Bentaleb, I.; Favier, C.; Fontugne, M.; Gillet, J.-F.; Sebag, D. Climatic and cultural changes in the west congo basin forests over the past 5000 years. Philos. Trans. R. Soc. B 2013, 368, 20120304. [Google Scholar] [CrossRef] [PubMed]
- Otto, F.E.; Jones, R.G.; Halladay, K.; Allen, M.R. Attribution of changes in precipitation patterns in African rainforests. Philos. Trans. R. Soc. B 2013, 368, 20120299. [Google Scholar] [CrossRef] [PubMed]
- Ouédraogo, D.Y.; Mortier, F.; Gourlet-Fleury, S.; Freycon, V.; Picard, N. Slow-growing species cope best with drought: Evidence from long-term measurements in a tropical semi-deciduous moist forest of Central Africa. J. Ecol. 2013, 101, 1459–1470. [Google Scholar] [CrossRef]
- Willis, K.; Bennett, K.D.; Burrough, S.; Macias-Fauria, M.; Tovar, C. Determining the response of African biota to climate change: Using the past to model the future. Philos. Trans. R. Soc. B 2013, 368, 20120491. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Tian, Y.; Myneni, R.B.; Ciais, P.; Saatchi, S.; Liu, Y.Y.; Piao, S.; Chen, H.; Vermote, E.F.; Song, C. Widespread decline of congo rainforest greenness in the past decade. Nature 2014, 509, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.A.; Hanan, N.P.; Neff, J.C.; Scholes, R.J.; Berry, J.A.; Denning, A.S.; Baker, D.F. Africa and the global carbon cycle. Carbon Balance Manag. 2007, 2, 3. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.B.; Sikka, M.; Sitch, S.; Ciais, P.; Poulter, B.; Galbraith, D.; Lee, J.E.; Huntingford, C.; Viovy, N.; Zeng, N. African tropical rainforest net carbon dioxide fluxes in the twentieth century. Philos. Trans. R. Soc. B 2013, 368, 20120376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, M.; Zhang, Q.; Shugart, H.H. Dynamic responses of African ecosystem carbon cycling to climate change. Clim. Res. 2001, 17, 183–193. [Google Scholar] [CrossRef]
- McGuire, A.; Sitch, S.; Clein, J.; Dargaville, R.; Esser, G.; Foley, J.; Heimann, M.; Joos, F.; Kaplan, J.; Kicklighter, D.W.; et al. Carbon balance of the terrestrial biosphere in the twentieth century: Analyses of CO2, climate and land use effects with four process-based ecosystem models. Glob. Biogeochem. Cycles 2001, 15, 183–206. [Google Scholar] [CrossRef]
- Ciais, P.; Piao, S.-L.; Cadule, P.; Friedlingstein, P.; Chédin, A. Variability and recent trends in the African terrestrial carbon balance. Biogeosciences 2009, 6, 1935–1948. [Google Scholar] [CrossRef]
- Hiernaux, P.; Mougin, E.; Diarra, L.; Soumaguel, N.; Lavenu, F.; Tracol, Y.; Diawara, M. Sahelian rangeland response to changes in rainfall over two decades in the gourma region, Mali. J. Hydrol. 2009, 375, 114–127. [Google Scholar] [CrossRef]
- Lewis, S.L.; Lopez-Gonzalez, G.; Sonké, B.; Affum-Baffoe, K.; Baker, T.R.; Ojo, L.O.; Phillips, O.L.; Reitsma, J.M.; White, L.; Comiskey, J.A. Increasing carbon storage in intact African tropical forests. Nature 2009, 457, 1003–1006. [Google Scholar] [CrossRef] [PubMed]
- Ciais, P.; Bombelli, A.; Williams, M.; Piao, S.; Chave, J.; Ryan, C.; Henry, M.; Brender, P.; Valentini, R. The carbon balance of Africa: Synthesis of recent research studies. Philos. Trans. R. Soc. B 2011, 369, 2038–2057. [Google Scholar] [CrossRef] [PubMed]
- Washington, R.; James, R.; Pearce, H.; Pokam, W.M.; Moufouma-Okia, W. Congo basin rainfall climatology: Can we believe the climate models? Philos. Trans. R. Soc. B 2013, 368, 20120296. [Google Scholar] [CrossRef] [PubMed]
- DosAnjos, L.; Oliva, M.A.; Kuki, K.N.; Mielke, M.S.; Ventrella, M.C.; Galvão, M.F.; Pinto, L.R. Key leaf traits indicative of photosynthetic plasticity in tropical tree species. Trees 2015, 29, 247–258. [Google Scholar] [CrossRef]
- Atkin, O.K.; Bloomfield, K.J.; Reich, P.B.; Tjoelker, M.G.; Asner, G.P.; Bonal, D.; Bönisch, G.; Bradford, M.G.; Cernusak, L.A.; Cosio, E.G. Global variability in leaf respiration in relation to climate, plant functional types and leaf traits. New Phytol. 2015, 206, 614–636. [Google Scholar] [CrossRef] [PubMed]
- Fatichi, S.; Leuzinger, S.; Körner, C. Moving beyond photosynthesis: From carbon source to sink-driven vegetation modeling. New Phytol. 2014, 201, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Doughty, C.E.; Metcalfe, D.; Girardin, C.; Amézquita, F.F.; Cabrera, D.G.; Huasco, W.H.; Silva-Espejo, J.; Araujo-Murakami, A.; da Costa, M.; Rocha, W. Drought impact on forest carbon dynamics and fluxes in Amazonia. Nature 2015, 519, 78–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhi, Y.; Doughty, C.E.; Goldsmith, G.R.; Metcalfe, D.B.; Girardin, C.A.; Marthews, T.R.; Aguila-Pasquel, J.; Aragão, L.E.; Araujo-Murakami, A.; Brando, P. The linkages between photosynthesis, productivity, growth and biomass in lowland Amazonian forests. Glob. Chang. Biol. 2015, 21, 2283–2295. [Google Scholar] [CrossRef] [PubMed]
- Kearsley, E.; de Haulleville, T.; Hufkens, K.; Kidimbu, A.; Toirambe, B.; Baert, G.; Huygens, D.; Kebede, Y.; Defourny, P.; Bogaert, J. Conventional tree height-diameter relationships significantly overestimate aboveground carbon stocks in the central congo basin. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef] [PubMed]
- Picard, N.; Henry, M.; Fonton, N.H.; Kondaoulé, J.; Fayolle, A.; Birigazzi, L.; Sola, G.; Poultouchidou, A.; Trotta, C.; Maïdou, H. Error in the estimation of emission factors for forest degradation in central Africa. J. For. Res. 2016, 21, 23–30. [Google Scholar] [CrossRef]
- Saatchi, S.; Mascaro, J.; Xu, L.; Keller, M.; Yang, Y.; Duffy, P.; Espírito-Santo, F.; Baccini, A.; Chambers, J.; Schimel, D. Seeing the forest beyond the trees. Glob. Ecol. Biogeogr. 2015, 24, 606–610. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Galmes, J.; Medrano, H.; Ribas-Carbó, M. Keeping a positive carbon balance under adverse conditions: Responses of photosynthesis and respiration to water stress. Physiol. Plant. 2006, 127, 343–352. [Google Scholar] [CrossRef]
- Flexas, J.; Galmes, J.; Ribas-Carbo, M.; Medrano, H. The effects of water stress on plant respiration. In Plant Respiration; Lambers, H., Ribas-Carbo, M., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 85–94. [Google Scholar]
- Varone, L.; Gratani, L. Leaf respiration responsiveness to induced water stress in Mediterranean species. Environ. Exp. Bot. 2015, 109, 141–150. [Google Scholar] [CrossRef]
- De Schepper, V.; Bühler, J.; Thorpe, M.; Roeb, G.; Huber, G.; van Dusschoten, D.; Jahnke, S.; Steppe, K. 11C-pet imaging reveals transport dynamics and sectorial plasticity of oak phloem after girdling. Front. Plant Sci. 2013, 4, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mencuccini, M. Temporal scales for the coordination of tree carbon and water economies during droughts. Tree Physiol. 2014, 34, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Sevanto, S. Phloem transport and drought. J. Exp. Bot. 2014, 65, 1751–1759. [Google Scholar] [CrossRef] [PubMed]
- Hubeau, M.; Steppe, K. Plant-PET scans: In vivo mapping of xylem and phloem functioning. Trends Plant Sci. 2015, 20, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Steppe, K.; Sterck, F.; Deslauriers, A. Diel growth dynamics in tree stems: Linking anatomy and ecophysiology. Trends Plant Sci. 2015, 20, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.G.; Hubbard, R.M.; Pongracic, S.; Raison, R.; McMurtrie, R.E. Foliage, fine-root, woody-tissue and stand respiration in Pinus radiata in relation to nitrogen status. Tree Physiol. 1996, 16, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Van der Molen, M.K.; Dolman, A.J.; Ciais, P.; Eglin, T.; Gobron, N.; Law, B.E.; Meir, P.; Peters, W.; Phillips, O.L.; Reichstein, M.; et al. Drought and ecosystem carbon cycling. Agric. For. Meteorol. 2011, 151, 765–773. [Google Scholar] [CrossRef]
- Gustafson, E.J.; De Bruijn, A.M.; Pangle, R.E.; Limousin, J.M.; McDowell, N.G.; Pockman, W.T.; Sturtevant, B.R.; Muss, J.D.; Kubiske, M.E. Integrating ecophysiology and forest landscape models to improve projections of drought effects under climate change. Glob. Chang. Biol. 2015, 21, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Turgeon, R. The role of phloem loading reconsidered. Plant Physiol. 2010, 152, 1817–1823. [Google Scholar] [CrossRef] [PubMed]
- Vico, G.; Dralle, D.; Feng, X.; Thompson, S.; Manzoni, S. How competitive is drought deciduousness in tropical forests? A combined eco-hydrological and eco-evolutionary approach. Environ. Res. Lett. 2017, 12, 065006. [Google Scholar] [CrossRef]
- Epila, J.; Maes, W.H.; Verbeeck, H.; Van Camp, J.; Okullo, J.B.L.; Steppe, K. Plant measurements on African tropical Maesopsis eminii seedlings contradict pioneering water use behaviour. Environ. Exp. Bot. 2017, 135, 27–37. [Google Scholar] [CrossRef]
- Van Camp, J.; Hubeau, M.; Van den Bulcke, J.; Van Acker, J.; Steppe, K. Cambial pinning relates wood anatomy to ecophysiology in the African tropical tree Maesopsis eminii. Tree Physiol. 2018, 38, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Caird, M.A.; Richards, J.H.; Donovan, L.A. Nighttime stomatal conductance and transpiration in C3 and C4 plants. Plant Physiol. 2007, 143, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Gao, J.; Zhao, P.; McCarthy, H.R.; Zhu, L.; Ni, G.; Ouyang, L. Tree species with photosynthetic stems have greater nighttime sap flux. Front. Plant Sci. 2018, 9, 30. [Google Scholar] [CrossRef] [PubMed]
- Epila, J.; De Baerdemaeker, N.J.F.; Vergeynst, L.L.; Maes, W.H.; Beeckman, H.; Steppe, K. Capacitive water release and internal leaf water relocation delay drought-induced cavitation in African Maesopsis eminii. Tree Pysiol. 2017, 37, 481–490. [Google Scholar]
- Tyree, M.T.; Engelbrecht, B.M.; Vargas, G.; Kursar, T.A. Desiccation tolerance of five tropical seedlings in Panama. Relationship to a field assessment of drought performance. Plant Physiol. 2003, 132, 1439–1447. [Google Scholar] [PubMed]
- Engelbrecht, B.M.; Kursar, T.A.; Tyree, M.T. Drought effects on seedling survival in a tropical moist forest. Trees 2005, 19, 312–321. [Google Scholar] [CrossRef]
- Wang, K.; Kellomäki, S.; Laitinen, K. Effects of needle age, long-term temperature and CO2 treatments on the photosynthesis of scots pine. Tree Physiol. 1995, 15, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Cannell, M.; Thornley, J. Temperature and CO2 responses of leaf and canopy photosynthesis: A clarification using the non-rectangular hyperbola model of photosynthesis. Ann. Bot. 1998, 82, 883–892. [Google Scholar] [CrossRef]
- Thornley, J. Dynamic model of leaf photosynthesis with acclimation to light and nitrogen. Ann. Bot. 1998, 81, 421–430. [Google Scholar] [CrossRef]
- Epron, D.; Bahn, M.; Derrien, D.; Lattanzi, F.A.; Pumpanen, J.; Gessler, A.; Hogberg, P.; Maillard, P.; Dannoura, M.; Gerant, D.; et al. Pulse-labelling trees to study carbon allocation dynamics: A review of methods, current knowledge and future prospects. Tree Physiol. 2012, 32, 776–798. [Google Scholar] [CrossRef] [PubMed]
- Zuidema, P.A.; Baker, P.J.; Groenendijk, P.; Schippers, P.; van der Sleen, P.; Vlam, M.; Sterck, F. Tropical forests and global change: Filling knowledge gaps. Trends Plant Sci. 2013, 18, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Kiser, M.R.; Reid, C.D.; Crowell, A.S.; Phillips, R.P.; Howell, C.R. Exploring the transport of plant metabolites using positron emitting radiotracers. HFSP J. 2008, 2, 189–204. [Google Scholar] [CrossRef] [PubMed]
- Minchin, P.E.H.; Thorpe, M.R. Using the short-lived isotope 11C in mechanistic studies of photosynthate transport. Funct. Plant Biol. 2003, 30, 831–841. [Google Scholar] [CrossRef]
- Bloemen, J.; Bauweraerts, I.; De Vos, F.; Vanhove, C.; Vandenberghe, S.; Boeckx, P.; Steppe, K. Fate of xylem-transported 11C- and 13C-labeled CO2 in leaves of poplar. Physiol. Plant. 2015, 153, 555–564. [Google Scholar] [CrossRef] [PubMed]
- LI-CO. Using the LI-6400/LI-6400XT Portable Photosynthesis System; LI-COR: Lincoln, NE, USA, 2008. [Google Scholar]
- Fang, L.; Zhang, S.; Zhang, G.; Liu, X.; Xia, X.; Zhang, S.; Xing, W.; Fang, X. Application of five light-response models in the photosynthesis of populus × euramericana cv. ‘Zhonglin46’ leaves. Appl. Biochem. Biotechnol. 2015, 176, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Calama, R.; Puértolas, J.; Madrigal, G.; Pardos, M. Modeling the environmental response of leaf net photosynthesis in Pinus pinea L. Natural regeneration. Ecol. Model. 2013, 251, 9–21. [Google Scholar] [CrossRef]
- Levin, C.S.; Hoffman, E.J. Calculation of positron range and its effect on the fundamental limit of positron emission tomography system spatial resolution. Phys. Med. Biol. 1999, 44, 781–799. [Google Scholar] [CrossRef] [PubMed]
- Jødal, L.; Le Loirec, C.; Champion, C. Positron range in PET imaging: An alternative approach for assessing and correcting the blurring. Phys. Med. Biol. 2012, 57, 3931. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Ehleringer, J.R. 13C/12C fractionation and its utility in terrestrial plant studies. In Carbon Isotope Techniques; Coleman, D.C., Fry, B., Eds.; Academic Press: San Diego, CA, USA, 1991; pp. 187–200. [Google Scholar]
- O’Leary, M.H. Carbon isotopes in photosynthesis. Bioscience 1988, 38, 328–336. [Google Scholar] [CrossRef]
- Koller, S.; Holland, V.; Brüggemann, W. Effects of drought stress on the evergreen Quercus ilex L., the deciduous Q. robur L. And their hybrid Q. × turneri willd. Photosynthetica 2013, 51, 574–582. [Google Scholar] [CrossRef]
- Varone, L.; Ribas-Carbo, M.; Cardona, C.; Gallé, A.; Medrano, H.; Gratani, L.; Flexas, J. Stomatal and non-stomatal limitations to photosynthesis in seedlings and saplings of Mediterranean species pre-conditioned and aged in nurseries: Different response to water stress. Environ. Exp. Bot. 2012, 75, 235–247. [Google Scholar] [CrossRef]
- Loustalot, A.J. Influence of soil moisture conditions on apparent photosynthesis and transpiration of pecan leaves. J. Agric. Res. 1945, 71, 519–532. [Google Scholar]
- Upchurch, R.P.; Peterson, M.L.; Hagan, R.M. Effect of soil-moisture content on the rate of photosynthesis and respiration in ladino clover (Trifolium repens L.). Plant Physiol. 1995, 30, 297–303. [Google Scholar] [CrossRef]
- De Swaef, T.; Steppe, K.; Lemeur, R. Determining reference values for stem water potential and maximum daily trunk shrinkage in young apple trees based on plant responses to water deficit. Agric. Water Manag. 2009, 96, 541–550. [Google Scholar] [CrossRef]
- Pantin, F.; Simonneau, T.; Muller, B. Coming of leaf age: Control of growth by hydraulics and metabolics during leaf ontogeny. New Phytol. 2012, 196, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Peguero-Pina, J.J.; Sancho-Knapik, D.; Morales, F.; Flexas, J.; Gil-Pelegrín, E. Differential photosynthetic performance and photoprotection mechanisms of three Mediterranean evergreen oaks under severe drought stress. Funct. Plant Biol. 2009, 36, 453–462. [Google Scholar] [CrossRef]
- Ackerly, D.D.; Bazzaz, F.A. Leaf dynamics, self-shading and carbon gain in seedlings of a tropical pioneer tree. Oecologia 1995, 101, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Tinoco-Ojanguren, C.; Pearcy, R.W. A comparison of light quality and quantity effects on the growth and steady-state and dynamic photosynthetic characteristics of three tropical tree species. Funct. Ecol. 1995, 9, 222–230. [Google Scholar] [CrossRef]
- Fetene, M.; Feleke, Y. Growth and photosynthesis of seedlings of four tree species from a dry tropical afromontane forest. J. Trop. Ecol. 2001, 17, 269–283. [Google Scholar] [CrossRef]
- Meir, P.; Levy, P.E.; Grace, J.; Jarvis, P.G. Photosynthetic parameters from two contrasting woody vegetation types in west Africa. Plant Ecol. 2007, 192, 277–287. [Google Scholar] [CrossRef]
- Riddoch, I.; Grace, J.; Fasehun, F.E.; Riddoch, B.; Ladipo, D.O. Photosynthesis and Successional Status of Seedlings in a Tropical Semi-Deciduous Rain Forest in Nigeria. J. Ecol. 1990, 79, 491–503. [Google Scholar] [CrossRef]
- Ellis, A.R.; Hubbell, S.P.; Potvin, C. In situ field measurements of photosynthetic rates of tropical tree species: A test of the functional group hypothesis. Can. J. Bot. 2000, 78, 1336–1347. [Google Scholar]
- Domingues, T.F.; Meir, P.; Feldpausch, T.R.; Saiz, G.; Veenendaal, E.M.; Schrodt, F.; Bird, M.; Djagbletey, G.; Hien, F.; Compaore, H.; et al. Co-limitation of photosynthetic capacity by nitrogen and phosphorus in West Africa woodlands. Plant Cell Environ. 2010, 33, 959–980. [Google Scholar] [CrossRef] [PubMed]
- Bürkle, L.; Hibberd, J.M.; Quick, W.P.; Kühn, C.; Hirner, B.; Frommer, W.B. The H(+)-sucrose cotransporter NtSUT1 is essential for sugar export from Tobacco leaves. Plant Physiol. 1998, 118, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Jahnke, S.; Schlesinger, U.; Feige, G.B.; Knust, E.J. Transport of photoassimilates in young trees of Fraxinus and Sorbus: Measurement of translocation in vivo. Bot. Acta 1998, 111, 307–315. [Google Scholar] [CrossRef]
- Wong, J.H.H.; Randall, D.D. Translocation of photoassimilate from leaves of two popyploid genotypes of tall fescue differing in photosynthetic rates. Physiol. Plant. 1985, 63, 445–450. [Google Scholar] [CrossRef]
- Rennie, E.A.; Turgeon, R. A comprehensive picture of phloem loading strategies. Proc. Natl. Acad. Sci. USA 2009, 106, 14162–14167. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Cheng, L.; Guo, Y.; Turgeon, R. Phloem loading strategies and water relations in trees and herbaceous plants. Plant Physiol. 2011, 157, 1518–1527. [Google Scholar] [CrossRef] [PubMed]
- Zavaliev, R.; Sagi, G.; Gera, A.; Epel, B.L. The constitutive expression of Arabidopsis plasmodesmal-associated class 1 reversibly glycosylated polypeptide impairs plant development and virus spread. J. Exp. Bot. 2010, 61, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Slewinski, T.L.; Braun, D.M. Current perspectives on the regulation of whole-plant carbohydrate partitioning. Plant Sci. 2010, 178, 341–349. [Google Scholar] [CrossRef]
- Epila, J.; Verbeeck, H.; Otim-Epila, T.; Okullo, P.; Kearsley, E.; Steppe, K. The ecology of Maesopsis eminii Engl. in tropical Africa. J. Afr. Ecol. 2017. [Google Scholar] [CrossRef]
- Jensen, K.H.; Savage, J.A.; Holbrook, N.M. Optimal concentration for sugar transport in plants. J. R. Soc. Interface 2013, 10, 20130055. [Google Scholar] [CrossRef] [PubMed]
- Scheiter, S.; Langan, L.; Higgins, S.I. Next-generation dynamic global vegetation models: Learning from community ecology. New Phytol. 2013, 198, 957–969. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measured Values | Non-Rectangular Hyperbola Model Predicted Values | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Measurement Day | N | P2000 | Rd | P2000 | Rd | Pn,max | α | Ls at 70% Pn,max | Ic | R2 |
| μmol CO2 m−2 s−1 | μmol CO2 m−2 s−1 | μmol CO2 (μmol PAR)−1 | µmol PAR m−2 s−1 | |||||||
| Tdrought | 10 | 1.9 ± 0.4 a | 1.0 ± 0.1 a | 1.8 ± 0.3 a | 1.0 ± 0.2 a | 1.9 ± 0.3 a | 0.042 ± 0.008 a | 80 ± 20 | 50 ± 13 | 0.964 |
| Trecovering | 12 | 4.0 ± 0.9 b | 2.0 ± 0.2 b | 4.5 ± 1.0 b | 2.1 ± 0.2 b | 4.8 ± 1.0 b | 0.069 ± 0.005 b | 71 ± 14 | 33 ± 6 | 0.990 |
| Twell-watered | 10 | 4.9 ± 0.6 b | 1.4 ± 0.2 a,b | 4.8 ± 0.7 b | 1.5 ± 0.2 a,b | 4.9 ± 0.7 b | 0.074 ± 0.009 b | 27 ± 7 | 27 ± 7 | 0.992 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Epila, J.; Hubeau, M.; Steppe, K. Drought Effects on Photosynthesis and Implications of Photoassimilate Distribution in 11C-Labeled Leaves in the African Tropical Tree Species Maesopsis eminii Engl. Forests 2018, 9, 109. https://doi.org/10.3390/f9030109
Epila J, Hubeau M, Steppe K. Drought Effects on Photosynthesis and Implications of Photoassimilate Distribution in 11C-Labeled Leaves in the African Tropical Tree Species Maesopsis eminii Engl. Forests. 2018; 9(3):109. https://doi.org/10.3390/f9030109
Chicago/Turabian StyleEpila, Jackie, Michiel Hubeau, and Kathy Steppe. 2018. "Drought Effects on Photosynthesis and Implications of Photoassimilate Distribution in 11C-Labeled Leaves in the African Tropical Tree Species Maesopsis eminii Engl." Forests 9, no. 3: 109. https://doi.org/10.3390/f9030109
APA StyleEpila, J., Hubeau, M., & Steppe, K. (2018). Drought Effects on Photosynthesis and Implications of Photoassimilate Distribution in 11C-Labeled Leaves in the African Tropical Tree Species Maesopsis eminii Engl. Forests, 9(3), 109. https://doi.org/10.3390/f9030109