Identification of Potential Metabolic Markers for the Selection of a High-Yield Clone of Quercus acutissima in Clonal Seed Orchard

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Sample Preparation
2.2. Metabolic Profiling and Hormone Analysis
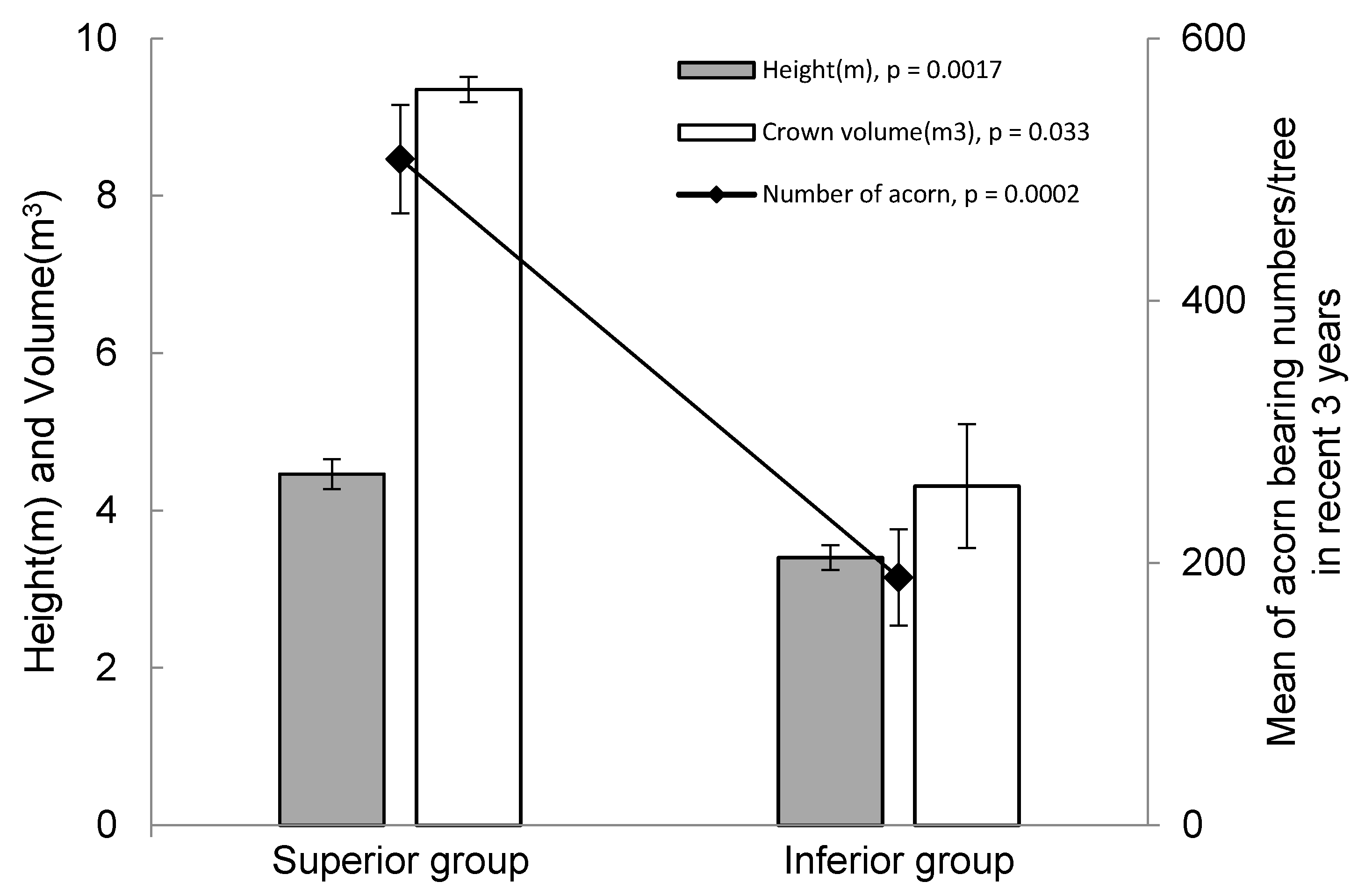
3. Results and Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wu, T.; Wang, G.G.; Wu, Q.; Cheng, X.; Yu, M.; Wang, W.; Yu, X. Patterns of leaf nitrogen and phosphorus stoichiometry among Quercus acutissima provenances across China. Ecol. Complex. 2014, 17, 32–39. [Google Scholar] [CrossRef]
- Kang, K.S.; Kim, C.S.; El-Kassaby, Y.A. Clonal variation in acorn production and its effect on the effective population size in a Quercus acutissima seed orchard. Silvae Genet. 2010, 59, 170–175. [Google Scholar] [CrossRef]
- Hwang, J.T.; Choi, H.K.; Kim, S.H.; Chung, S.; Hur, H.J.; Park, J.H.; Chung, M.Y. Hypolipidemic activity of Quercus acutissima fruit ethanol extract is mediated by inhibition of acetylation. J. Med. Food 2017, 20, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Lee, T. Coloured Flora of Korea; Hayangmunsa: Seoul, Korea, 2003. [Google Scholar]
- Darley-Hill, S.; Johnson, W.C. Acorn dispersal by blue jay (Cyanocitta cristata). Oecologia 1981, 50, 231–232. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.E.; Koo, C.D.; Kim, J.S.; Park, J.I.; Shin, W.S.; Shin, C.S. Ecological characteristics of below-ground ectomycorrhizal colony of Sarcodon aspratus in oak tree stands. J. Korean For. Soc. 2002, 91, 457–464. [Google Scholar]
- Yim, K.B.; Min, Y.T.; Kim, Y.M.; Han, S.D.; Kwon, H.M. Oak Trees; Institute of Forest Genetics: Suwon, Korea, 1995; pp. 35–52. [Google Scholar]
- Han, Y.C.; Chung, H.G.; Min, Y.T.; Kang, K.S. Plus Tree Handbook; Forest Genetics Research Institute: Suwon, Korea, 1996; pp. 180–195. [Google Scholar]
- Sork, V.L.; Bramble, E. Prediction of acorn crops in three species of North American oaks: Quercus alba, Q. rubra and Q. velutina. Ann. Sci. For. 1993, 50, 128s–136s. [Google Scholar] [CrossRef]
- Gilland, K.E.; Keiffer, C.H.; McCarthy, B.C. Seed production of mature forest-grown American chestnut (Castanea dentata (Marsh.) Borkh). J. Torrey Bot. Soc. 2012, 139, 283–289. [Google Scholar] [CrossRef]
- Pourhashemi, M.; Panahi, P.; Zandebasiri, M. Application of visual surveys to estimate acorn production of Brant's oak (Quercus brantii Lindl.) in northern Zagros Forests of Iran. Caspi. J. Environ. Sci. 2013, 11, 85–95. [Google Scholar]
- Kim, H.T.; Kang, J.W.; Lee, W.Y.; Han, S.U.; Park, E.J. Estimation of acorn production capacity using growth characteristics of Quercus acutissima in a clonal seed orchard. For. Sci. Technol. 2016, 12, 51–54. [Google Scholar]
- Fernandez, O.; Urrutia, M.; Bernillon, S.; Giauffret, C.; Tardieu, F.; Le Gouis, J.; Langlade, N.; Charcosset, A.; Moing, A.; Gibon, Y. Fortune telling: Metabolic markers of plant performance. Metabolomics 2016, 12, 158. [Google Scholar] [CrossRef] [PubMed]
- Weckwerth, W. Integration of metabolomics and proteomics in molecular plant physiology-coping with the complexity by data-dimensionality reduction. Physol. Plant. 2008, 132, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Hamzehzarghani, H.; Kushalappa, A.C.; Dion, Y.; Rioux, S.; Comeau, A.; Yaylayan, V.; Marshall, W.D.; Mather, D. Metabolic profiling and factor analysis to discriminate quantitative resistance in wheat cultivars against fusarium head blight. Physiol. Mol. Plant Pathol. 2005, 66, 119–133. [Google Scholar] [CrossRef]
- Witta, S.; Galiciab, L.; Liseca, J.; Cairnsc, J.; Tiessend, A.; Arause, J.L.; Rojasb, N.P.; Ferniea, A.R. Metabolic and phenotypic responses of greenhouse-grown maize hybrids to experimentally controlled drought stress. Mol. Plant. 2012, 5, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Robinson, A.R.; Gheneim, R.; Kozak, R.A.; Ellis, D.D.; Mansfield, S.D. The potential of metabolite profiling as a selection tool for genotype discrimination in Populus. J. Exp. Bot. 2005, 56, 2807–2819. [Google Scholar] [CrossRef] [PubMed]
- Robinson, A.R.; Ukrainetz, N.K.; Kang, K.Y.; Mansfield, S.D. Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trials reveals strong environmental and weak genetic variation. New Phytol. 2007, 174, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Ossipov, V.; Ossipova, S.; Bykov, V.; Oksanen, E.; Koricheva, J.; Haukioja, E. Application of metabolomics to genotype and phenotype discrimination of birch trees grown in a long-term open-field experiment. Metabolomics 2008, 4, 39–51. [Google Scholar] [CrossRef]
- Farrow, S.C.; Emery, R.N. Concurrent profiling of indole-3-acetic acid, abscisic acid, and cytokinins and structurally related purines by high-performance-liquid-chromatography tandem electrospray mass spectrometry. Plant Methods 2012, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Kusaba, S.; Masashi, F.; Chikako, H.; Isomaro, Y.; Tomoaki, S.; Yuriko, K.M. Decreased GA1 Content Caused by the Overexpression ofOSH1 Is Accompanied by Suppression of GA 20-Oxidase Gene Expression. Plant Physiol. 1998, 117, 1179–1184. [Google Scholar] [CrossRef] [PubMed]
- Pearce, D.W.; Rood, S.B.; Wu, R. Phytohormones and shoot growth in a three-generation hybrid poplar family. Tree Physiol. 2004, 24, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Pharis, R.P.; Yeh, F.C.; Bruce, P.D. Superior growth potential in trees: What is its basis, and can it be tested for at an early age? Can. J. For. Res. 1991, 21, 368–374. [Google Scholar] [CrossRef]
- Todorova, D.; Genkov, T.; Vaseva-Gemisheva, I.; Alexieva, V.; Karanov, E.; Smith, A.; Hall, M. Effect of temperature stress on the endogenous cytokinin content in Arabidopsis thaliana (L.) Heynh plants. Acta Physiol. Plant. 2005, 27, 13–18. [Google Scholar] [CrossRef]
- Scott, I.M.; Horgan, R.; McGaw, B.A. Zeatin-9-glucoside, a major endogenous cytokinin of Vinca rosea L. crown gall tissue. Planta 1980, 149, 472–475. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.C.; Mok, M.C.; Mok, D.W.S. A gene encoding the cytokinin enzyme zeatinO-xylosyltransferase of Phaseolus vulgaris. Plant Physiol. 1999, 120, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Horgan, R. A new cytokinin metabolite. Biochem. Biophys. Res. Commun. 1975, 65, 358–363. [Google Scholar] [CrossRef]
- Parker, C.W.; Letham, D.S.; Gollnow, B.I.; Summons, R.E.; Duke, C.C.; MacLeod, J.K. Regulators of cell division in plant tissues XXV. Metabolism of zeatin by lupin seedlings. Planta 1978, 142, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Duke, C.C.; Letham, D.S.; Parker, C.W.; MacLeod, J.K.; Summons, R.E. The complex of 0-glucosylzeatin derivatives formed in Populus species. Phytochemistry 1979, 18, 819–824. [Google Scholar] [CrossRef]
- Toubiana, D.; Fait, A. Metabolomics-assisted crop breeding towards improvement in seed quality and yield. In Seed Development: Omics Technologies Toward Improvement of Seed Quality and Crop Yield; Springer: Dordrecht, The Netherlands, 2012; pp. 453–475. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Metabolites | Relative Contents (Mean) | Fold Changes (Superior/Inferior) | t-Test p-Value | |
|---|---|---|---|---|
| Superior Group | Inferior Group | |||
| <leaves> | ||||
| Ethylene glycol | 74 | 34 | 2.18 | 0.045 |
| Butane-1,3-diol | 509 | 194 | 2.62 | 0.049 |
| 2(3H)-Furanone | 62 | 23 | 2.70 | 0.042 |
| Malic acid | 3514 | 1520 | 2.31 | 0.048 |
| Butane-1,4-diol | 264 | 100 | 2.64 | 0.019 |
| Rythronic acid-2 | 53 | 25 | 2.12 | 0.046 |
| Ononitol | 10,663 | 4997 | 2.13 | 0.048 |
| Phenylpropanolamine | 32,444 | 11,389 | 2.85 | 0.021 |
| Quinic acid | 22,284 | 9341 | 2.39 | 0.026 |
| D-Ribose | 1727 | 968 | 1.78 | 0.002 |
| Galactose | 205 | 75 | 2.73 | 0.044 |
| Myo-inositol | 172 | 90 | 1.91 | 0.037 |
| Muco-inositol | 282 | 97 | 2.91 | 0.017 |
| Sucrose | 39,070 | 14,068 | 2.78 | 0.025 |
| Maltose | 456 | 153 | 2.98 | 0.035 |
| <stems> | ||||
| Phosphoric acid | 146 | 86 | 1.70 | 0.043 |
| Succinic acid | 10 | 5 | 2.02 | 0.000 |
| Rythronic acid | 42 | 30 | 1.40 | 0.021 |
| Xylitol | 15 | 10 | 1.51 | 0.033 |
| Unknown 6 | 9 | 5 | 1.79 | 0.018 |
| Metabolites | Acorn Bearing Numbers/Tree | Crown Volume |
|---|---|---|
| Correlation Coefficient | Correlation Coefficient | |
| Phosphoric acid | 0.4301 * | 0.2380 |
| Succinic acid | 0.5317 * | 0.2252 |
| Malic acid | 0.5430 ** | 0.2821 |
| Butane-1,3-diol | 0.5951 ** | 0.3848 |
| Xylitol | 0.4308 * | 0.3070 |
| Isocitric acid | 0.4738 * | 0.6522 ** |
| Glucitol | 0.4591 * | 0.4317 * |
| Maltose | 0.4110 * | 0.3657 |
| Rythronic acid | 0.2458 | 0.4498 * |
| Characteristic of Acorn Production | Hormone Contents (ng/g Dry Weight ) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| (9G)Z | (9R)Z | (7G)iP | iP | (9R) DZ | BA | cis-Z | IAA | ABA | IAA/ ABA | |
| Superior group | 11.4 ± 1.6 | 108.8 ± 34.7 | 57.2 ± 25.5 | 150.4 ± 102.0 | 19.5 ± 6.1 | 6.8 ± 2.6 | 1.9 ± 0.6 | 280.5 ± 166.0 | 1,802.9 ± 666.3 | 0.19 ± 0.15 |
| Inferior group | 14.5 ± 2.0 | 107.2 ± 30.4 | 47.1 ± 22.5 | 94.3 ± 40.7 | 18.4 ± 5.6 | 6.2 ± 4.0 | 1.5 ± 0.6 | 299.2 ± 72.1 | 2,197.6 ± 576.2 | 0.15 ± 0.05 |
| Sig. | ** | - | - | - | - | - | - | - | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, J.W.; Lee, H.; Lim, H.; Lee, W.Y. Identification of Potential Metabolic Markers for the Selection of a High-Yield Clone of Quercus acutissima in Clonal Seed Orchard. Forests 2018, 9, 116. https://doi.org/10.3390/f9030116
Kang JW, Lee H, Lim H, Lee WY. Identification of Potential Metabolic Markers for the Selection of a High-Yield Clone of Quercus acutissima in Clonal Seed Orchard. Forests. 2018; 9(3):116. https://doi.org/10.3390/f9030116
Chicago/Turabian StyleKang, Jun Won, Hyunseok Lee, Hyemin Lim, and Wi Young Lee. 2018. "Identification of Potential Metabolic Markers for the Selection of a High-Yield Clone of Quercus acutissima in Clonal Seed Orchard" Forests 9, no. 3: 116. https://doi.org/10.3390/f9030116
APA StyleKang, J. W., Lee, H., Lim, H., & Lee, W. Y. (2018). Identification of Potential Metabolic Markers for the Selection of a High-Yield Clone of Quercus acutissima in Clonal Seed Orchard. Forests, 9(3), 116. https://doi.org/10.3390/f9030116