1. Introduction
Eastern white pine (
Pinus strobus L.) is a coniferous species with a natural range extending from southeastern Canada, through the Midwest and Northeast regions of the United States and southward through the Southern Appalachian Mountains and into adjacent upper Piedmont regions [
1]. Commercially, the wood is used for lumber, cabinetry, millwork, and toys. The species is used as an ornamental and for reforestation, where it is often planted on eroded or degraded sites [
1,
2]. In North Carolina, 39,790 ha of Eastern white pine are in timber production [
3] and 95% of the volume harvested is grown in the mountainous western region of the state [
4].
Eastern white pine is also a worthy Christmas tree species with soft blue-green foliage, good post-harvest needle retention, relatively strong branches but only a slight aroma. [
1,
5] In North Carolina, it has traditionally been grown in the mountains for cut Christmas trees and greenery, however, growers have extended the range eastward into the Piedmont and Coastal Plain regions. There, it is grown on ‘choose and cut’ farms and often cultivated with a suite of other species including Virginia pine (
Pinus virginiana Mill.), Eastern redcedar (
Juniperus virginiana L.), and various cultivars of Leyland cypress (
Cuprocyparis leylandii (A.B. Jacks. & Dallim.) Farjon), pure cypress (
Cupressus), and arborvitae (
Thuja). Eastern white pine is popular in this region, where it is the most widely planted species. A 2008 survey revealed that 78% of the farms in the region grow Eastern white pine and it accounts for 42% of trees sold [
6].
Today, Fraser fir (
Abies fraseri (Pursh) Poir.) is the most popular Christmas tree species grown in the southern Appalachians, where revenue from annual sales exceed
$US 100 million in North Carolina alone [
7]. However, Fraser fir is highly susceptible to root rot disease caused primarily by
Phytophthora cinnamomi Rands, an introduced soil-borne pathogen with an extensive host range [
8,
9,
10]. Resistance to this pathogen has not been found in Fraser fir and chemical methods of control are stop-gap at best [
11,
12]. Because
P. cinnamomi can persist in soil for decades, infested sites are often removed from Fraser fir production. Faced with this reality, growers commonly plant Eastern white pine on infested sites where Fraser fir cannot survive. This strategy is mostly successful, the exceptions being extremely high disease hazard areas (i.e., sites with extremely poor drainage and high inoculum load) [
13]. Kirby and Grand 1975 [
14] demonstrated that
P. cinnamomi is pathogenic to Eastern white pine. Infected seedlings showed stunting of new growth, chlorosis, necrosis, needle loss, and severe root rot. On lower Piedmont and Coastal Plain sites, especially where Eastern white pine is planted outside its natural range, it is more sensitive to Phytophthora root rot and recently an increase in disease has occurred on Christmas tree farms in this region. Annual losses of
$US 15 million to the littleleaf disease complex of southern pines [
15] and
$US 9 million to root rot disease in Fraser fir [
16], both also caused by
P. cinnamomi, underscore the concern over potential losses in Eastern white pine Christmas tree and forest plantations.
Genetic resistance is widely used to ameliorate impacts of diseases caused by
Phytophthora spp. in agriculture and horticulture [
8], however, nothing is known about genetic resistance in Eastern white pine. As such, this study sought to ascertain the degree of genetic control, if any, of resistance to
P. cinnamomi in Eastern white pine and secondarily, to compare the aggressiveness of two
P. cinnamomi cultures isolated from different host species (Fraser fir and Eastern white pine). The long-term goal is to reduce Phytophthora root rot damage by identifying and deploying families with higher resistance.
2. Materials and Methods
2.1. Plant Production
The North Carolina Forest Service (NCFS) provided white pine seedlings from their Nursery and Tree Improvement Program for use in this study. The parent trees originated from the southern portion of the natural white pine range, primarily North Carolina. They were selected from natural stands (first generation) or progeny test plantings (second generation) for forest production with emphasis on total stem volume but also considering straightness, stem taper, self-pruning, crown density, and branch angle. It is unknown if any of the selection sites were infested with Phytophthora, but if so, only symptomless trees would have been selected. In 2015, one-year-old seedlings of 83 open-pollinated families grown in a greenhouse at the NCFS Linville River Nursery in Avery County, N.C., were transferred into a greenhouse at the Horticulture Field Lab, North Carolina State University, Raleigh. Seedlings of the same family had been grown in the same flats and so were rearranged into the experimental design described below while in the greenhouse and then transferred into an outdoor shade house (40% shade). All seedlings were fertilized bi-weekly with Peters 15–16–17 Peat Lite Special (150–200 ppm N) (The Scotts Co., Marysville, OH, USA) in the spring. While in the shade house, the seedlings were automatically irrigated two times daily for 20 min (about 10 mm/day) until it was discontinued for winter and the seedlings were moved back into a greenhouse. During spring 2016, seedlings were again moved outside into the shade house and top-dressed with slow release fertilizer (Multicote 6, 18-6-12 + micronutrients, Haifa Chemicals Ltd., Haifa, Israel) for another set of inoculations and disease assessments under a similar irrigation regime.
2.2. Inoculum Production and Inoculation
A large main study was conducted using a standard culture of
P. cinnamomi, 23ss04, isolated from Fraser fir and employed in several published studies involving fir (
Abies) and chestnut (
Castanea) species [
11,
12,
17,
18]. For comparison, a smaller supplemental study was conducted using isolate 2334 from Eastern white pine. Inoculum of both isolates were prepared by autoclaving Erlenmeyer flasks containing 25 g of long grain rice and 18 mL deionized water two consecutive times prior to adding 3–4 corn agar plugs of the isolates [
19]. The flasks were shaken daily to avoid clumping as mycelia colonized the grains. Seedlings were inoculated with colonized rice grains by making one 2 cm deep hole about 1 cm from each side of the stem, inserting a grain, and then pushing the medium back to cover the inoculum [
20] (
Figure 1). The first inoculation occurred in June 2015 and surviving seedlings were inoculated a second time eight weeks later in August. In June 2016, one year and one week after the first inoculation, all living seedlings were re-inoculated and again eight weeks later in the same manner with inoculum from the appropriate isolate. Depending on availability, up to three non-inoculated control seedlings of each family were grown in the shade house throughout the study in a block beside the inoculated seedlings.
2.3. Experimental Design and Data Collection
Both studies were set up with a randomized block design including five blocks (
Table 1). The main study included seedlings from all 83 families, while the supplemental study only included 20 families. Up to 15 seedlings per family were included in each block occupying one row of a tray (Forest Tray 135, Stuewe and Sons, Tangent, OR, USA). For families with insufficient numbers, seedlings were equally distributed across blocks so that the family size in the main study ranged from 40 to 75. All families selected for the supplemental study had enough seedlings to allocate 15 per block.
Biweekly disease symptom assessments began two weeks after the first inoculation and continued through week 16. In the second year, to account for winter mortality, an assessment was made immediately before the first inoculation followed by eight additional biweekly assessments. At each assessment, seedling health was assessed by estimating percent shoot (needle and stem) necrosis on a 0 to 100 scale in increments of 10 but also including 1%, 2%, and 95% levels.
In 2016, after the final disease assessment, roots from inoculated and control seedlings were tested with Agdia ImmunoStrips® (Elkhart, IN, USA) for Phytophthora. Roots from 10 randomly selected inoculated seedlings (5 dead and 5 alive) from the main study were tested, as well as 10 randomly selected control seedlings (5 dead and 5 alive).
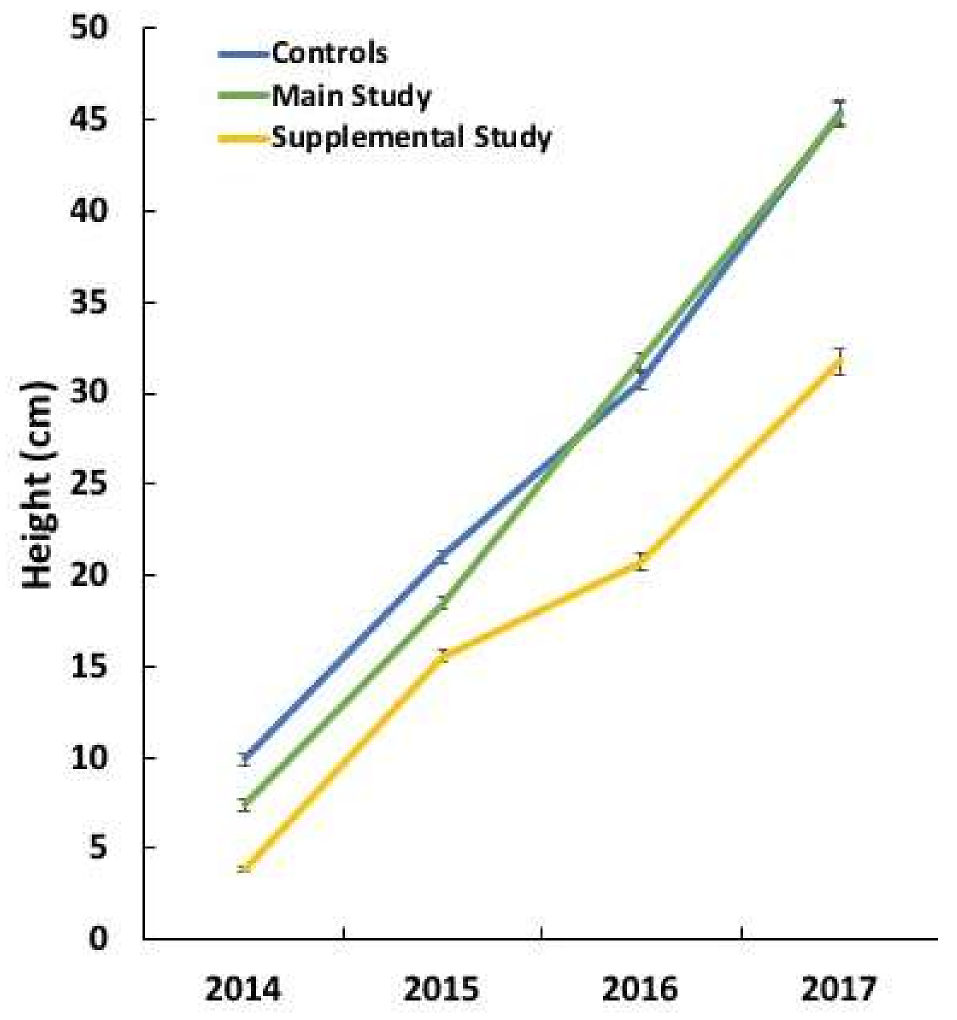
After the study, seedlings spent the 2016–2017 winter in the outdoor shade house. Subsequent to completing their 2017 apical growth, height measurements were taken on a subset of healthy seedlings with no disease symptoms. For six families, the heights of the previous four years (2014–2017) were measured for seedlings from the main study (n = 132 total, 4–38/family), the supplemental study (n = 101 total, 9–23/family), and the control group (n = 17 total, 2–3/family).
2.4. Statistical Analyses and Heritability Estimates
Prior to statistical analyses, the disease scale was simplified to a binomial mortality rating, where seedlings were considered dead (1) when they displayed 100% shoot necrosis and alive (0) otherwise. During the first four weeks of the study, some seedlings displayed obvious signs of transplant shock as a result of rearranging them into the experimental design. This confused symptom assessment so that this information was recorded and these seedlings were excluded from subsequent analyses.
Family means for mortality were calculated for each assessment period of the main and supplemental studies. Individual analyses of variance were carried out for each assessment period of the main and supplemental studies using the GLIMMIX procedure of SAS Enterprise Guide 7.11 HF3 (7.100.1.2805) (64-bit) (SAS Institute Inc., Cary, NC, USA) with block (fixed effect), family (random effect), and their interaction (random effect) as sources of variation. Because mortality was the response variable, a binomial distribution with a logistic link function was employed. This same model was used in another analysis of the main study data using only the 20 families in common with the supplemental study. Similarly, combined analyses of variance were conducted for the 20 families in common to both studies with study (fixed effect), block (study) (fixed effect), family (random effect), and their interactions as sources of variation.
Individual-tree (
h2i) and family mean (
h2f) heritabilities for mortality were estimated using variance components obtained from the GLIMMIX procedure in SAS as follows:
where:
The error variance for mortality was set to π
2/3 = 3.29 in calculation of phenotypic variances, as suggested by Gilmour et al. (1985) [
21] for binary traits. We assumed that the family variance component was about one-quarter of additive genetic variance [
22]. Standard errors of heritability were estimated using the delta method [
23].
Height data from the non-symptomatic seedlings of six families were subjected to analyses of variance for each of the four measurement years (2014–2017) using study (fixed effect), family (random effect), and their interaction (random effect) as sources of variation. Differences among the least squares means of the main study, supplemental study, and control group were compared using Tukey-Kramer tests at the α = 0.05 level.
4. Discussion
Eastern white pine proved to be relatively resistant to root rot caused by
P. cinnamomi. The relatively low mortality in these studies is contrary to the 87% mortality reported by Kirby and Grand (1975). The cause of this discrepancy is unknown but may be due to their use of potentially more infectious inoculation techniques, wetter conditions, and, perhaps, more susceptible host material and/or a more aggressive
P. cinnamomi isolate. The studies reported here were initially planned for one year but because of the relatively low mortality (18.6%) in the main study at the end of the first year, they were extended for another year with additional inoculations. In contrast to the first-year results for Eastern white pine, Fraser fir and American chestnut (
Castanea dentata (March.) Borkh.) approached 100 and 90 percent mortality, respectively, during the same time period when challenged with the same inoculum (23ss04) using the same inoculation methods, and under similar environmental conditions [
12,
17,
18]. Furthermore, in a similar resistance screening of 32
Abies species, only the single most resistant species demonstrated less mortality than Eastern white pine [
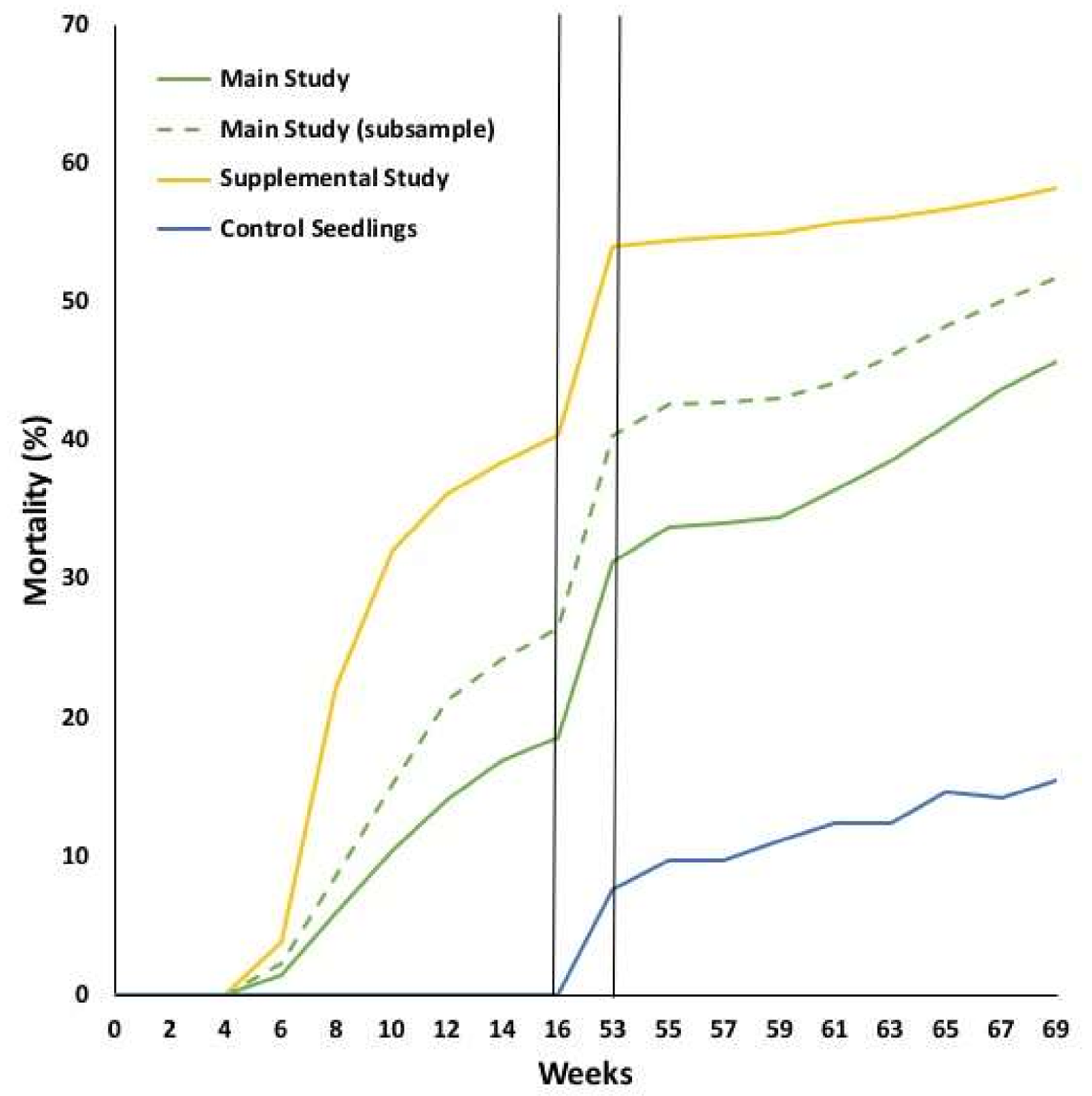
11]. Continuing the studies reported here into the second year did not provide additional information about genetic control of resistance. Instead, the results were confounded by the overwintering environment and the effect of the increasingly crowded conditions on both the root systems and crowns of the seedlings. These effects are manifest in the second-year mortality of the non-inoculated control seedlings (
Figure 2), a sub-sample of which all tested negative for
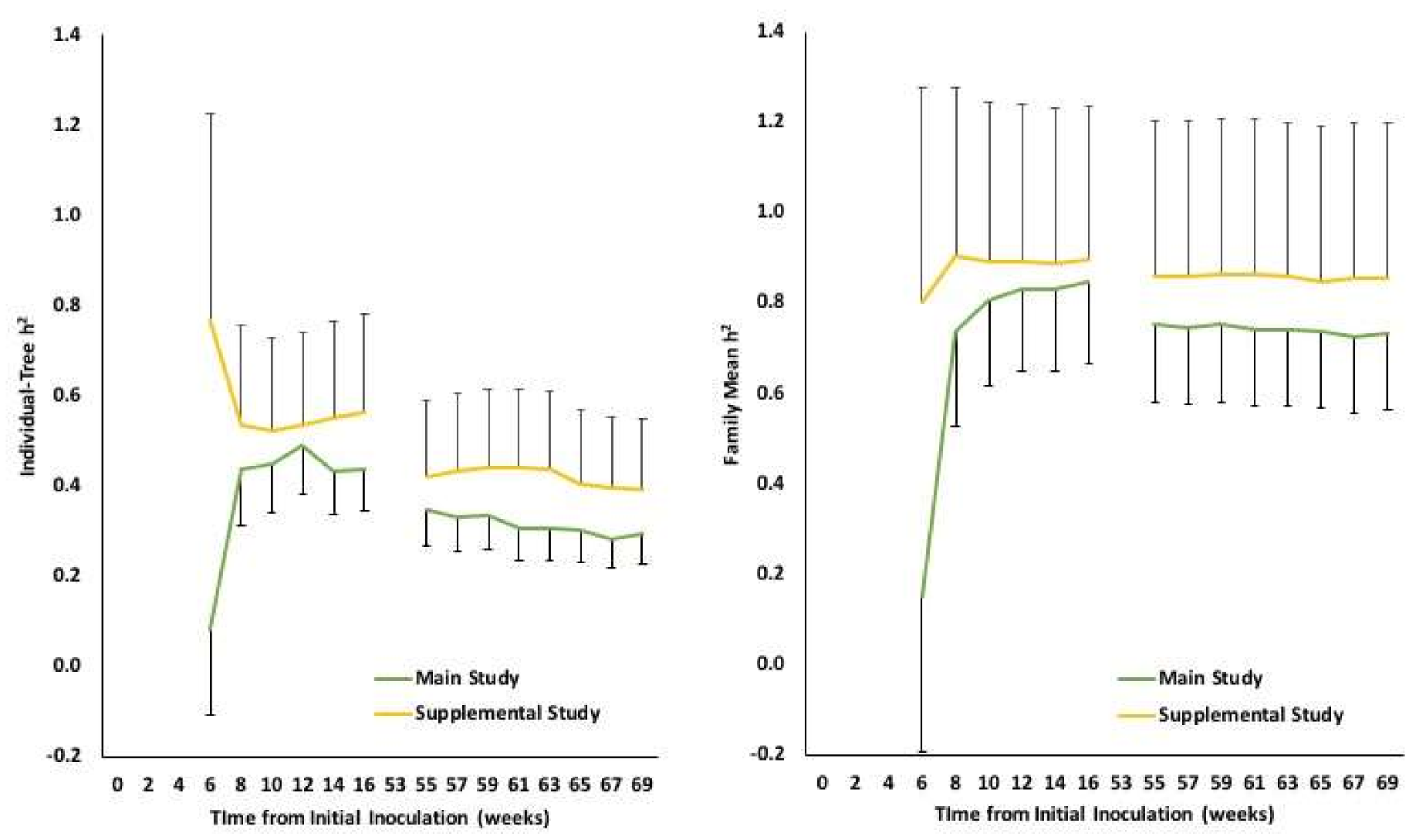
Phytophthora. Due to the increased environmental effects on mortality, the heritability estimates generally decreased from year one to year two, although they remained relatively stable throughout the second year (
Figure 3). Future evaluations of Phytophthora root rot resistance in Eastern white pine seedlings can be relatively rapid with a final assessment between 12 and 16 weeks after the initial inoculation.
Although Eastern white pine is relatively resistant to
P. cinnamomi, there is considerable variation among families within the species. It should be noted that the families evaluated in this study originated from selections made primarily in North Carolina, a relatively small portion of the expansive range of Eastern white pine. The amount of variation in resistance to an introduced pathogen is rather remarkable. Possibly, a pattern of geographic variation for resistance exists, as has been shown for Trojan (
Abies equi-trojani Aschers. et Sint) and Turkish (
A. bornmuelleriana Mattf.) fir in Turkey [
12]. Unfortunately, family deployment recommendations are complicated by the large interaction observed between the pine families and the two
P. cinnamomi isolates studied. The isolate derived from Eastern white pine (supplemental study) was clearly more aggressive (40.4% mortality) than the isolate derived from Fraser fir (26.4% mortality) on the same 20 families. Interestingly, after the first year (before environmental effects considerably confounded mortality), the mortality heritability estimates for the same 20 families were higher when challenged by the isolate from white pine relative to being challenged with the isolate from Fraser fir (
Table 1). However, the isolate × family interaction variance component was high and the correlation of family mortality means between studies (isolates) was not significant. This was surprising because these two isolates (among others) had been used in a previous
Abies study [
17]. In that study, the magnitude of mortality among four fir species inoculated with these isolates was similar with identical species mortality rankings. Further, the correlation of family mortality means between isolates 2334 and 23ss04 was high (
r = 0.87,
p ≤ 0.0001). Apparently, isolate 2334 has some attribute allowing it to be specifically aggressive to Eastern white pine. Three of the twenty families averaged 20% mortality or less across both isolates and will be recommended for deployment. Planting these families versus the most susceptible or even an average mix of families could substantially improve survival and decrease economic losses associated with Christmas tree and forestry regeneration efforts. Clearly, however, resistance evaluations of more families using isolate 2334 and other isolates derived from Eastern white pine are needed. In retrospect, it would have been preferable to have inoculated all 83 families with isolate 2334.
In addition to causing more mortality, the P. cinnamomi isolate from Eastern white pine caused a reduction in height growth in inoculated but non-symptomatic seedlings relative to the non-inoculated control seedlings, while the isolate from Fraser fir did not. It is not known whether these seedlings were infected or continually engaged in resisting infection of Phytophthora in the medium, but in either case, it appears that more resources were required to maintain defense responses to the isolate from white pine. This has important practical implications for planting stock producers, especially bare-root nurseries, that encounter Phytophthora problems. In addition to the risk of spreading the pathogen, infected non-symptomatic seedlings will likely reduce productivity when established in Christmas tree and forest plantations.
Warm soil temperatures (>50 F or >10 °C) are one of several epidemiological factors (including average soil pH (pH 4.5–6) and saturated soils) that increase damage caused by
P. cinnamomi via favoring sporangium formation, zoospore release, and infection. A recent study demonstrated a dramatic increase in root rot severity and mortality of seven
Abies species caused by
P. cinnamomi and three other
Phytophthora species under a warm (27–32 °C) versus cool (15–21 °C) environment [
24]. Differences in soil temperature provide at least a partial explanation for the contrasting experiences of North Carolina Christmas tree growers, where in the mountains, Eastern white pine is deliberately planted on sites known to be infested with
Phytophthora, often without obvious deleterious effects, versus in the Piedmont and Coastal Plain regions, where infected trees commonly die. Warmer soil temperatures associated with climate change will not only increase disease severity and mortality rate but are also expected to expand the range of
P. cinnamomi [
25], thus further increasing the threat to both Christmas tree and forest plantations.
In addition to climate change, the introduction of new
Phytophthora species is another serious threat to Eastern white pine. Recently, the number of
Phytophthora species contributing to losses in Fraser fir Christmas trees in the Southern Appalachians has increased [
26,
27]. In a 2014 survey of Fraser fir Christmas tree plantations in the region, six
Phytophthora species were identified, three of which were isolated for the first time in the region [
27].
P. cryptogea Pethybr. & Laff., in particular, appears to have rapidly spread. Once introduced and dispersed in a new area,
Phytophthora species have proven to be nearly impossible to control and these new species are very likely to eventually threaten Eastern white pine. As such, the challenge to finding resistance may become substantially more complex and include the need to account for variation within
P. cinnamomi, as demonstrated by the studies reported here, as well as the need to evaluate resistance to other pathogen species.
Phytophthora species are certain to continue to cause losses to Eastern white pine Christmas tree and forest plantations. Nurseries, growers, and foresters all need to be knowledgeable about, and diligent in implementing, practices that reduce its spread, especially ensuring that planting stock is disease free [
13,
27]. Selection and use of genetically resistant material can play a key role in the overall strategy in order to ameliorate future catastrophic impacts of these intransigent pathogens.






{kind=link}
{kind=link}
{kind=link}
{kind=link}