Tropical Tree Species Effects on Soil pH and Biotic Factors and the Consequences for Macroaggregate Dynamics

Abstract
:1. Introduction
- Macroaggregate size distribution and chemistry differ among the six vegetation types. The four species in plantations and the Control plots would have started with similar soil structure at the beginning of this experiment, 25 years prior to our study. Thus, observed differences among vegetation types in the chemistry and quantity of organic matter (OM) inputs [18,20] would influence soil aggregation, depending on the relative importance of their binding material in this soil. A lack of difference in macroaggregate structure would indicate over-riding control by mineralogical interactions.
- Macroaggregate structure is correlated with fine-root growth, litterfall, fungal, and microbial effects on aggregation. Fine-root and litterfall additions are expected to bind aggregates and thus contribute to their stabilization via multiple chemical and physical mechanisms [11]. Fungal hyphae can reorient clay particles, bind particles with extracellular polysaccharides, and enmesh particles, thereby influencing soil aggregation [21] (Ternan et al., 1996). Mycorrhizal fungi can act at multiple scales, from the plant community to the soil mycelium, to influence soil aggregation [22]. In addition, microbial biomass C, rather than fungal hyphae alone, is often associated with factors that stabilize soil organic C (SOC), and could thus affect soil structure [23]. Microbial activity, however, differs temporally, and with aggregate size, soil type, cropping system and management [24,25].
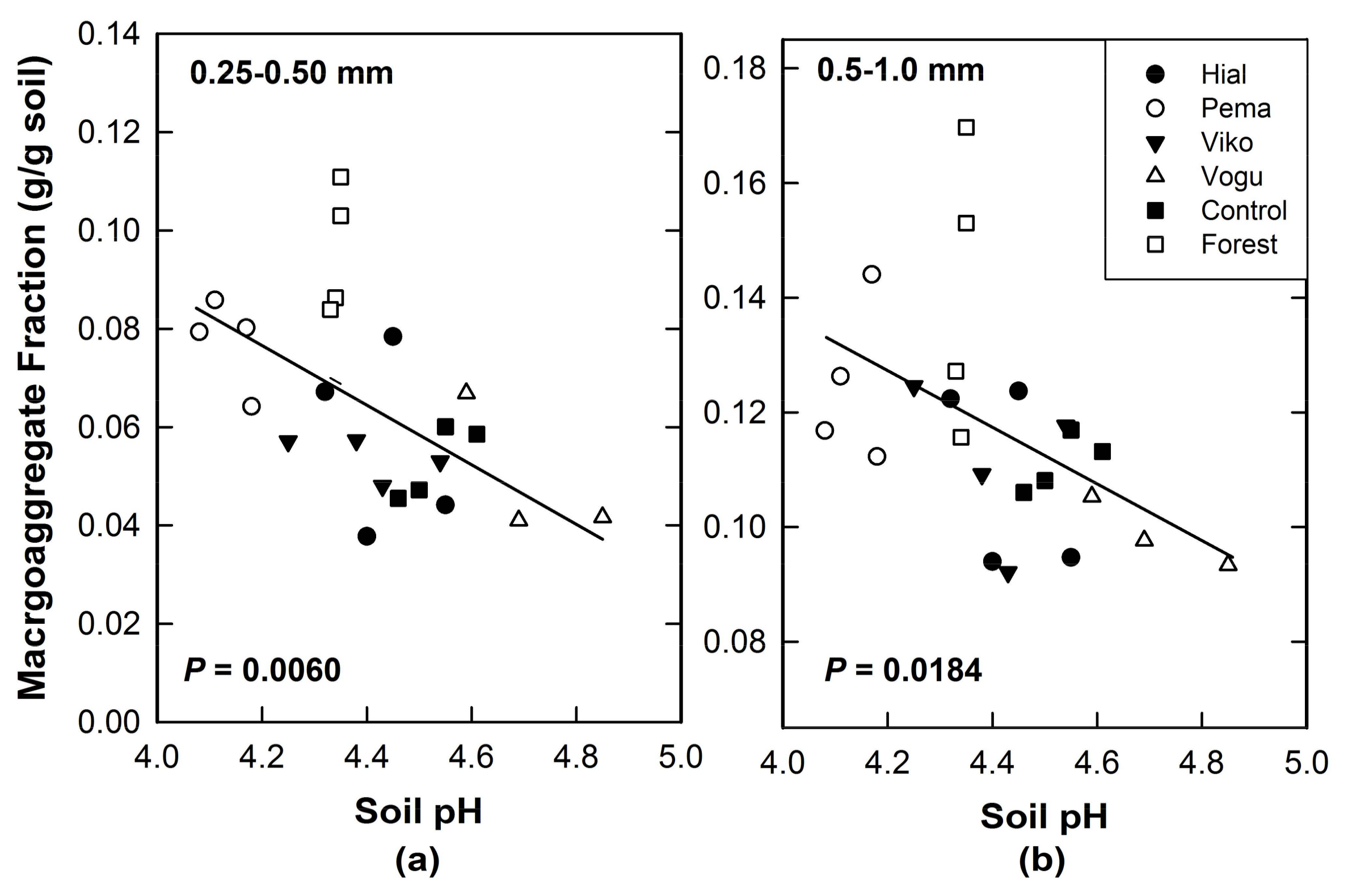
- Macroaggregate structure is correlated with soil pH. The tree species in this experiment have altered soil pH [16,26] and this could alter aggregation processes. Previously, it was hypothesized that in these variable-charge soils, changes in soil pH above or below the point of zero charge (PZC) would disperse colloids [16]. This effect on colloidal stability could thus influence macroaggregate stability.
2. Materials and Methods
2.1. Study Site and Experimental Design
2.2. Field Methods
2.3. Laboratory Analyses
2.4. Data Analysis
3. Results
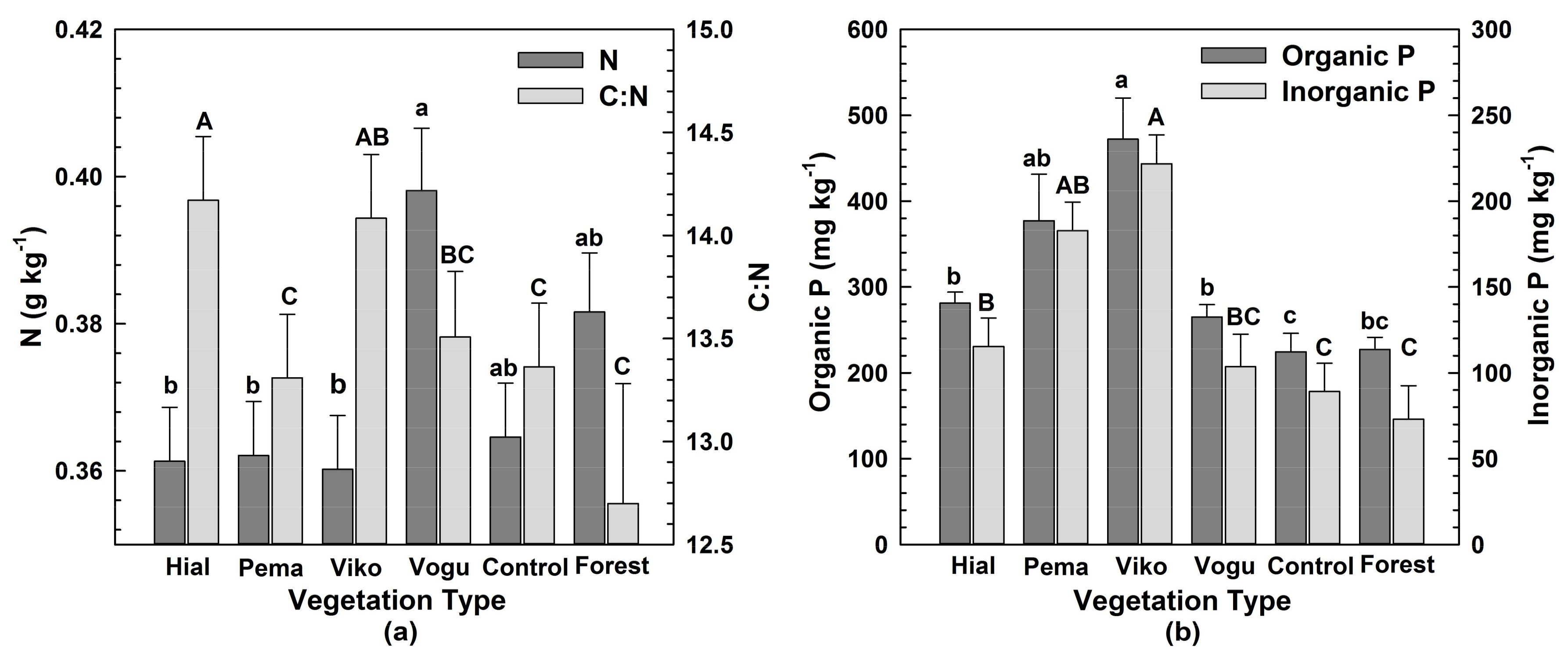
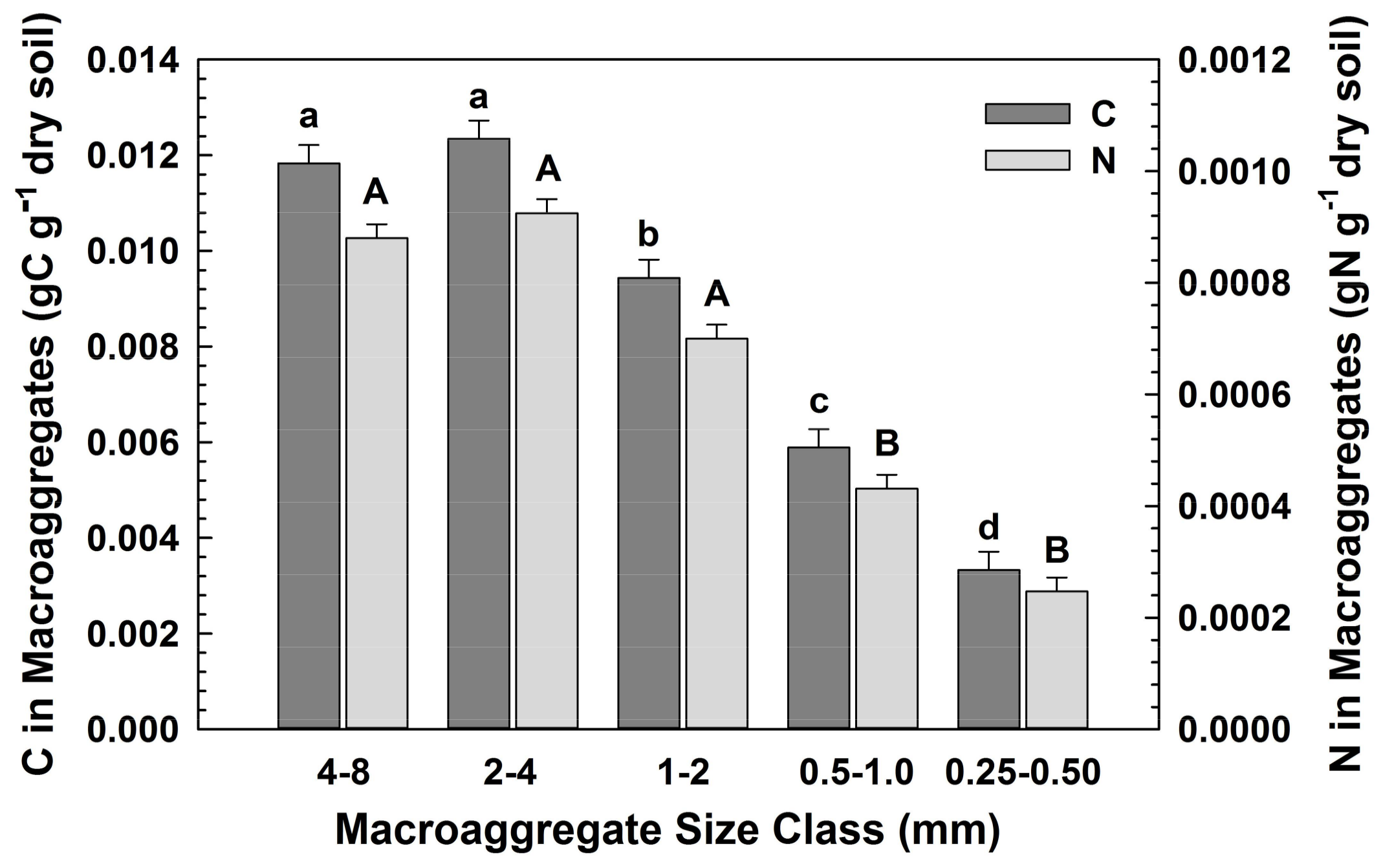
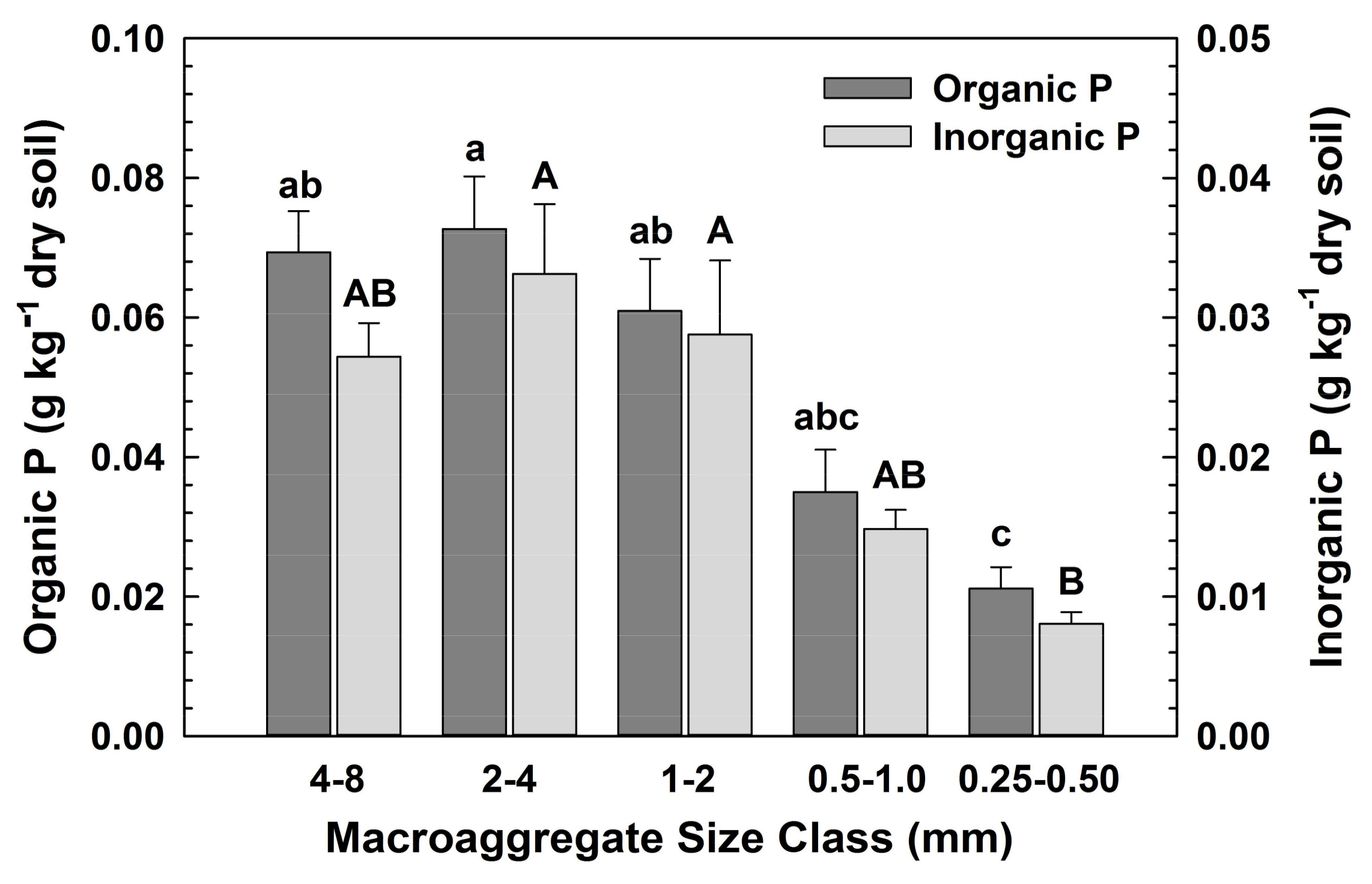
3.1. Macroaggregate Size Distribution and Element Concentrations under Different Vegetation (H1)
3.2. Biotic Effects on Macroaggregate Size Distributions (H2, H3)
4. Discussion
4.1. Cementing Effect of Soil Mineralogy
4.2. Binding Effects of the Biota
4.3. Dispersivity (pH) Effects of Tree Species
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation | Size Class | Fraction of Dry Mass | C | N | C:N | Organic P | Inorganic P |
|---|---|---|---|---|---|---|---|
| mm | g/g | g/kg | g/kg | mg/kg | mg/kg | ||
| Hieronyma | 4–8 | 0.227 ± 0.012 | 48.85 ± 1.87 | 3.57 ± 0.09 | 13.71 ± 0.45 | 247.81 ± 36.39 | 110.97 ± 13.19 |
| 2–4 | 0.246 ± 0.014 | 48.85 ± 2.22 | 3.53 ± 0.13 | 13.85 ± 0.43 | 283.93 ± 19.23 | 113.16 ± 15.27 | |
| 1–2 | 0.196 ± 0.009 | 50.78 ± 2.37 | 3.58 ± 0.13 | 14.17 ± 0.45 | 292.23 ± 10.48 | 119.34 ± 12.73 | |
| 0.5–1 | 0.109 ± 0.008 | 52.51 ± 2.57 | 3.65 ± 0.16 | 14.44 ± 0.68 | 323.97 | 125.28 | |
| 0.25–0.5 | 0.057 ± 0.009 | 54.93 ± 2.36 | 3.75 ± 0.12 | 14.68 ± 0.53 | 318.39 | 108.47 | |
| Pentaclethra | 4–8 | 0.221 ± 0.029 | 48.20 ± 4.95 | 3.61 ± 0.37 | 13.35 ± 0.09 | 400.50 ± 103.38 | 175.37 ± 66.12 |
| 2–4 | 0.251 ± 0.010 | 46.08 ± 3.53 | 3.57 ± 0.24 | 12.87 ± 0.24 | 390.63 ± 101.75 | 160.37 ± 55.96 | |
| 1–2 | 0.191 ± 0.010 | 46.94 ± 3.33 | 3.57 ± 0.27 | 13.17 ± 0.21 | 304.86 ± 142.77 | 258.00 ± 101.07 | |
| 0.5–1 | 0.125 ± 0.007 | 46.94 ± 3.64 | 3.60 ± 0.26 | 13.64 ± 0.20 | 488.09 | 156.27 | |
| 0.25–0.5 | 0.077 ± 0.005 | 49.16 ± 3.74 | 3.75 ± 0.24 | 13.51 ± 0.36 | 408.80 | 164.50 | |
| Virola | 4–8 | 0.243 ± 0.015 | 49.76 ± 1.81 | 3.56 ± 0.09 | 13.98 ± 0.36 | 428.60 ± 12.71 | 139.40 ± 10.64 |
| 2–4 | 0.256 ± 0.008 | 50.14 ± 2.29 | 3.60 ± 0.08 | 13.90 ± 0.35 | 412.34 ± 162.27 | 240.51 ± 99.42 | |
| 1–2 | 0.199 ± 0.007 | 50.31 ± 3.02 | 3.55 ± 0.14 | 14.16 ± 0.41 | 524.92 ± 96.56 | 232.16 ± 92.47 | |
| 0.5–1 | 0.111 ± 0.007 | 51.79 ± 3.14 | 3.63 ± 0.15 | 14.24 ± 0.51 | 535.67 | 234.02 | |
| 0.25–0.5 | 0.054 ± 0.002 | 52.00 ± 2.43 | 3.67 ± 0.08 | 14.15 ± 0.41 | 509.14 | 240.11 | |
| Vochysia | 4–8 | 0.285 ± 0.021 | 52.50 ± 2.61 | 3.92 ± 0.22 | 13.41 ± 0.31 | 233.30 ± 8.44 | 100.00 ± 5.52 |
| 2–4 | 0.270 ± 0.010 | 53.10 ± 2.02 | 3.82 ± 0.18 | 13.91 ± 0.22 | 232.07 ± 15.32 | 102.48 ± 5.44 | |
| 1–2 | 0.171 ± 0.009 | 53.19 ± 2.16 | 3.87 ± 0.18 | 13.77 ± 0.27 | 290.10 ± 24.19 | 99.88 ± 6.75 | |
| 0.5–1 | 0.099 ± 0.003 | 57.76 ± 2.01 | 4.17 ± 0.19 | 13.85 ± 0.15 | 293.99 | 102.09 | |
| 0.25–0.5 | 0.050 ±0.009 | 57.10 ± 1.98 | 4.10 ± 0.26 | 13.98 ± 0.41 | 356.02 | 100.18 | |
| Control | 4–8 | 0.244 ± 0.008 | 47.45 ± 2.51 | 3.58 ± 0.13 | 13.23 ± 0.33 | 242.59 ± 48.55 | 103.52 ± 18.15 |
| 2–4 | 0.262 ± 0.014 | 47.22 ± 1.69 | 3.62 ± 0.14 | 13.05 ± 0.21 | 228.57 ± 47.77 | 104.40 ± 23.58 | |
| 1–2 | 0.202 ± 0.006 | 48.63 ± 2.60 | 3.65 ± 0.16 | 13.30 ± 0.22 | 207.83 ± 43.05 | 95.42 ± 22.29 | |
| 0.5–1 | 0.111 ± 0.002 | 49.56 ± 2.95 | 3.66 ± 0.17 | 13.53 ± 0.34 | 166.38 | 81.69 | |
| 0.25–0.5 | 0.053 ± 0.004 | 50.96 ± 2.60 | 3.72 ± 0.14 | 13.70 ± 0.28 | 260.81 | 60.83 | |
| Mature Forest | 4–8 | 0.219 ± 0.021 | 50.85 ± 2.97 | 3.94 ± 0.17 | 12.88 ± 0.29 | 221.43 ± 26.57 | 72.51 ± 5.47 |
| 2–4 | 0.224 ± 0.014 | 50.43 ± 2.76 | 4.00 ± 0.13 | 12.59 ± 0.31 | 214.55 ± 34.30 | 76.75 ± 9.96 | |
| 1–2 | 0.190 ± 0.007 | 47.54 ± 2.79 | 3.74 ± 0.16 | 12.71 ± | 257.37 ± 18.74 | 69.18 ± 8.14 | |
| 0.5–1 | 0.142 ± 0.012 | 46.87 ± 3.18 | 3.67 ± 0.14 | 12.74 ± | 158.26 | 68.30 | |
| 0.25–0.5 | 0.096 ± 0.007 | 47.04 ± 2.76 | 3.74 ±0.12 | 12.57 ± | 251.20 | 78.68 |
| Vegetation | Hyphae in Aggregates | Microbial Biomass C | Fine-Root Growth | Al in Litterfall | Soil pH |
|---|---|---|---|---|---|
| mm g−1 Dry Soil | μg C g−1 Dry Soil | cm cm−2 yr−1 | kg Al ha−1 yr−1 | ||
| Hieronyma | 16,588 ± 1150 | 1022 ± 108 | 99 ± 8 | 10 ± 2 | 4.43 ± 0.05 |
| Pentaclethra | 19,201 ± 818 | 933 ± 49 | 70 ± 9 | 8 ± 1 | 4.14 ± 0.02 |
| Virola | 17,438 ± 1853 | 781 ± 180 | 91 ± 9 | 14 ± 1 | 4.40 ± 0.06 |
| Vochysia | 12,021 ± 1818 | 1171 ± 33 | 181 ± 15 | 180 ± 17 | 4.71 ± 0.08 |
| Control | 15,420 ± 1543 | 949 ± 139 | 106 ± 10 | 83 ± 14 | 4.53 ± 0.03 |
| Mature Forest | 16,548 ± 721 | 1033 ± 89 | 94 ± 11 | 2 ± 1 | 4.34 ± 0.01 |
References
- Jastrow, J.D.; Amonette, J.E.; Bailey, V.L. Mechanisms controlling soil carbon turnover and their potential application for enhancing carbon sequestration. Clim. Chang. 2007, 80, 5–23. [Google Scholar] [CrossRef]
- Tisdall, J.M.; Oades, J.M. Organic matter and water-stable aggregates in soils. Eur. J. Soil Sci. 1982, 33, 141–163. [Google Scholar] [CrossRef]
- Elliott, E. Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils. Soil Sci. Soc. Am. J. 1986, 50, 627–633. [Google Scholar] [CrossRef]
- Payne, D. Soil structure, tilth and mechanical behaviour. Russells Soil Cond. Plant Growth 1988, 11, 378–411. [Google Scholar]
- El-Swaify, S. Physical and mechanical properties of oxisols. In Soils with Variable Charge; Theng, B.K.G., Ed.; Offset Publ.: Palmerston North, New Zealand, 1980; pp. 303–324. [Google Scholar]
- Warkentin, B.; Maeda, T. Physical and mechanical characteristics of Andisols. N. Z. Soc. Soil Sci. 1980, 12, 281–302. [Google Scholar]
- Igwe, C.A.; Zarei, M.; Stahr, K. Fe and Al oxides distribution in some ultisols and inceptisols of southeastern Nigeria in relation to soil total phosphorus. Environ. Earth Sci. 2010, 60, 1103–1111. [Google Scholar] [CrossRef]
- Spycher, G.; Rose, S.L.; Sollins, P.; Norgren, J.; Young, J.L.; Kermit Cromack, J. Evolution of Structure in a chronosequence of andesitic forest soils. Soil Sci. 1986, 142, 164–178. [Google Scholar] [CrossRef]
- Tiessen, H.; Stewart, J.W. Light and electron microscopy of stained microaggregates: The role of organic matter and microbes in soil aggregation. Biogeochemistry 1988, 5, 312–322. [Google Scholar] [CrossRef]
- Oades, J.; Waters, A. Aggregate hierarchy in soils. Soil Res. 1991, 29, 815–828. [Google Scholar] [CrossRef]
- Bronick, C.J.; Lal, R. Soil structure and management: A review. Geoderma 2005, 124, 3–22. [Google Scholar] [CrossRef]
- Sollins, P.; Homann, P.; Caldwell, B.A. Stabilization and destabilization of soil organic matter: Mechanisms and controls. Geoderma 1996, 74, 65–105. [Google Scholar] [CrossRef]
- Six, J.; Feller, C.; Denef, K.; Ogle, S.; de Moraes Sa, J.C.; Albrecht, A. Soil organic matter, biota and aggregation in temperate and tropical soils-Effects of no-tillage. Agronomie 2002, 22, 755–775. [Google Scholar] [CrossRef]
- Finzi, A.C.; Canham, C.D.; Van Breemen, N. Canopy tree–soil interactions within temperate forests: Species effects on pH and cations. Ecol. Appl. 1998, 8, 447–454. [Google Scholar]
- Reich, P.B.; Oleksyn, J.; Modrzynski, J.; Mrozinski, P.; Hobbie, S.E.; Eissenstat, D.M.; Chorover, J.; Chadwick, O.A.; Hale, C.M.; Tjoelker, M.G. Linking litter calcium, earthworms and soil properties: An common garden test with 14 tree species. Ecol. Lett. 2005, 8, 811–818. [Google Scholar] [CrossRef]
- Russell, A.E.; Hall, S.J.; Raich, J.W. Tropical tree species traits drive soil cation dynamics via effects on pH: A proposed conceptual framework. Ecol. Monogr. 2017, 87, 685–701. [Google Scholar] [CrossRef]
- Fisher, R.F. Amelioration of degraded rain forest soils by plantations of native trees. Soil Sci. Soc. Am. J. 1995, 59, 544–549. [Google Scholar] [CrossRef]
- Raich, J.W.; Russell, A.E.; Bedoya-Arrieta, R. Lignin and enhanced litter turnover in tree plantations of lowland Costa Rica. For. Ecol. Manag. 2007, 239, 128–135. [Google Scholar] [CrossRef]
- Raich, J.W.; Russell, A.E.; Valverde-Barrantes, O. Fine root decay rates vary widely among lowland tropical tree species. Oecologia 2009, 161, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.E.; Raich, J.W.; Arrieta, R.B.; Valverde-Barrantes, O.; González, E. Impacts of individual tree species on carbon dynamics in a moist tropical forest environment. Ecol. Appl. 2010, 20, 1087–1100. [Google Scholar] [CrossRef] [PubMed]
- Ternan, J.; Elmes, A.; Williams, A.; Hartley, R. Aggregate stability of soils in central Spain and the role of land management. Earth Surf. Process. Landf. 1996, 21, 181–193. [Google Scholar] [CrossRef]
- Rillig, M.C.; Mummey, D.L. Mycorrhizas and soil structure. New Phytol. 2006, 171, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Schutter, M.E.; Dick, R.P. Microbial community profiles and activities among aggregates of winter fallow and cover-cropped soil. Soil Sci. Soc. Am. J. 2002, 66, 142–153. [Google Scholar] [CrossRef]
- Mendes, I.; Bandick, A.; Dick, R.; Bottomley, P. Microbial biomass and activities in soil aggregates affected by winter cover crops. Soil Sci. Soc. Am. J. 1999, 63, 873–881. [Google Scholar] [CrossRef]
- Russell, A.E.; Raich, J.W.; Valverde-Barrantes, O.J.; Fisher, R.F. Tree species effects on soil properties in experimental plantations in tropical moist forest. Soil Sci. Soc. Am. J. 2007, 71, 1389–1397. [Google Scholar] [CrossRef]
- Sanford, R.L., Jr.; Paaby, P.; Luvall, J.C.; Phillips, E. Climate, geomorphology, and aquatic systems. In La Selva: Ecology and Natural History of a Neotropical Rain Forest; University of Chicago Press: Chicago, IL, USA, 1994; pp. 19–33. [Google Scholar]
- Sollins, P.; Sancho, F.; Ch, R.M.; Sanford, R.L., Jr. So//s and Soil Process Research. In La Selva: Ecology and Natural History of a Neotropical Rain Forest; University of Chicago Press: Chicago, IL, USA, 1994; p. 34. [Google Scholar]
- Porder, S.; Clark, D.A.; Vitousek, P.M. Persistence of rock-derived nutrients in the wet tropical forests of La Selva, Costa Rica. Ecology 2006, 87, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Kleber, M.; Schwendenmann, L.; Veldkamp, E.; Rößner, J.; Jahn, R. Halloysite versus gibbsite: Silicon cycling as a pedogenetic process in two lowland neotropical rain forest soils of La Selva, Costa Rica. Geoderma 2007, 138, 1–11. [Google Scholar] [CrossRef]
- Chorover, J.; Sposito, G. Surface charge characteristics of kaolinitic tropical soils. Geochim. Cosmochim. Acta 1995, 59, 875–884. [Google Scholar] [CrossRef]
- Anda, M.; Shamshuddin, J.; Fauziah, C.; Omar, S.S. Mineralogy and factors controlling charge development of three Oxisols developed from different parent materials. Geoderma 2008, 143, 153–167. [Google Scholar] [CrossRef]
- Clark, D.B.; Clark, D.A. Landscape-scale variation in forest structure and biomass in a tropical rain forest. For. Ecol. Manag. 2000, 137, 185–198. [Google Scholar] [CrossRef]
- Kivlin, S.N.; Hawkes, C.V. Temporal and spatial variation of soil bacteria richness, composition, and function in a neotropical rainforest. PLoS ONE 2016, 11, e0159131. [Google Scholar] [CrossRef] [PubMed]
- Kivlin, S.N.; Hawkes, C.V. Tree species, spatial heterogeneity, and seasonality drive soil fungal abundance, richness, and composition in Neotropical rainforests. Environ. Microbiol. 2016, 18, 4662–4673. [Google Scholar] [CrossRef] [PubMed]
- Valverde-Barrantes, O.J.; Raich, J.W.; Russell, A.E. Fine-root mass, growth and nitrogen content for six tropical tree species. Plant Soil 2007, 290, 357–370. [Google Scholar] [CrossRef]
- Crosland, A.; Zhao, F.; McGrath, S.; Lane, P. Comparison of aqua regia digestion with sodium carbonate fusion for the determination of total phosphorus in soils by inductively coupled plasma atomic emission spectroscopy (ICP). Commun. Soil Sci. Plant Anal. 1995, 26, 1357–1368. [Google Scholar] [CrossRef]
- Kuo, S. Phosphorus. In Methods of Soil Analysis Part 3—Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Eds.; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1996; pp. 869–919. [Google Scholar]
- Kingston, H.M.; Haswell, S.J. Microwave-Enhanced Chemistry; American Chemical Society: Washington, DC, USA, 1997. [Google Scholar]
- Díaz-Romeu, R.; Hunter, A. Metodologías de Muestreo de Suelos, Análisis Químico de Suelos y Tejido Vegetal y de Investigaciones en Invernadero; Centro Agronómico Tropical de Investigación y Enseñanza (CATIE): Cartago, Costa Rica, 1978. [Google Scholar]
- Thomas, G. Soil pH and soil acidity. In Methods of Soil Analysis Part 3—Chemical Methods; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1996; pp. 475–490. [Google Scholar]
- Brundrett, M.; Melville, L.; Peterson, L. Practical Methods in Mycorrhiza Research: Based on a Workshop Organized in Conjuction with the Ninth North American Conference on Mycorrhizae, University of Guelph, Guelph, Ontario, Canada; Mycologue Publications: Devon, UK, 1994. [Google Scholar]
- Sylvia, D.M. 3 Quantification of External Hyphae of Vesicular-arbuscular Mycorrhizal Fungi. In Methods in Microbiology; Elsevier: Amsterdam, The Netherlands, 1992; Volume 24, pp. 53–65. [Google Scholar]
- Scott-Denton, L.E.; Rosenstiel, T.N.; Monson, R.K. Differential controls by climate and substrate over the heterotrophic and rhizospheric components of soil respiration. Glob. Chang. Biol. 2006, 12, 205–216. [Google Scholar] [CrossRef]
- Brookes, P.; Landman, A.; Pruden, G.; Jenkinson, D. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Littell, R.; Milliken, G.; Stroup, W.; Wolfinger, R. SAS Systems for Mixed Models; SAS Inst. Inc.: Cary, NC, USA, 1996. [Google Scholar]
- Neter, J.; Kutner, M.H.; Nachtsheim, C.J.; Wasserman, W. Applied Linear Statistical Models; Irwin Chicago: Chicago, IL, USA, 1996; Volume 4. [Google Scholar]
- Duiker, S.W.; Rhoton, F.E.; Torrent, J.; Smeck, N.E.; Lal, R. Iron (hydr) oxide crystallinity effects on soil aggregation. Soil Sci. Soc. Am. J. 2003, 67, 606–611. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Nat. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Prober, S.M.; Leff, J.W.; Bates, S.T.; Borer, E.T.; Firn, J.; Harpole, W.S.; Lind, E.M.; Seabloom, E.W.; Adler, P.B.; Bakker, J.D. Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide. Ecol. Lett. 2015, 18, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Barberán, A.; McGuire, K.L.; Wolf, J.A.; Jones, F.A.; Wright, S.J.; Turner, B.L.; Essene, A.; Hubbell, S.P.; Faircloth, B.C.; Fierer, N. Relating belowground microbial composition to the taxonomic, phylogenetic, and functional trait distributions of trees in a tropical forest. Ecol. Lett. 2015, 18, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Van Miegroet, H.; Cole, D. The Impact of Nitrification on Soil Acidification and Cation Leaching in a Red Alder Ecosystem 1. J. Environ. Qual. 1984, 13, 586–590. [Google Scholar] [CrossRef]
- Essington, M. Soil and Water Chemistry: An Integrative Approach; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]





| Response Variable | Explanatory Variable | ||
|---|---|---|---|
| Size (S) | Vegetation (V) | S × V | |
| Fraction (g g−1 dry soil) 1 | <0.0001 | 0.8952 | 0.0012 |
| C (g kg−1) | 0.3155 | 0.0781 | 0.9935 |
| N (g kg−1) | 0.7791 | 0.0478 | 0.9984 |
| C:N | 0.0420 | 0.0024 | 0.9367 |
| Inorganic P (mg kg−1) 2 | 0.7682 | 0.0008 | 0.9750 |
| Organic P (mg kg−1) | 0.9019 | 0.0006 | 0.9998 |
| C Fraction (gC g−1 dry soil) | <0.0001 | 0.5782 | 0.2863 |
| N Fraction (gN g−1 dry soil) | <0.0001 | 0.2100 | 0.3230 |
| Inorganic P Fraction (gP kg−1 dry soil) | <0.0001 | 0.0039 | 0.8511 |
| Organic P Fraction (gP kg−1 dry soil) | <0.0001 | 0.0003 | 0.7911 |
| Explanatory Variables 1 | Macroaggregate Size Class (mm) | ||||
|---|---|---|---|---|---|
| 4–8 | 2–4 | 1–2 | 0.5–1.0 | 0.25–0.50 | |
| p-value 2 | |||||
| Hyphae in Aggregates | 0.2185 | 0.9067 | 0.4062 | 0.6491 | 0.9383 |
| Microbial Biomass C | 0.7711 | 0.7080 | 0.2663 | 0.7559 | 0.2316 |
| Fine Root Growth | 0.0013 | 0.8827 | 0.0077 | 0.7732 | 0.5268 |
| Al in Litterfall | 0.7450 | 0.1382 | 0.4991 | 0.1079 | 0.0201 |
| Test Statistic | Macroaggregate Size Class (mm) | ||||
|---|---|---|---|---|---|
| 4–8 | 2–4 | 1–2 | 0.5–1.0 | 0.25–0.50 | |
| p | 0.5240 | 0.4122 | 0.5489 | 0.0563 | 0.0055 |
| r | 0.1400 | 0.1794 | 0.1319 | 0.4032 | 0.5595 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russell, A.E.; Kivlin, S.N.; Hawkes, C.V. Tropical Tree Species Effects on Soil pH and Biotic Factors and the Consequences for Macroaggregate Dynamics. Forests 2018, 9, 184. https://doi.org/10.3390/f9040184
Russell AE, Kivlin SN, Hawkes CV. Tropical Tree Species Effects on Soil pH and Biotic Factors and the Consequences for Macroaggregate Dynamics. Forests. 2018; 9(4):184. https://doi.org/10.3390/f9040184
Chicago/Turabian StyleRussell, Ann E., Stephanie N. Kivlin, and Christine V. Hawkes. 2018. "Tropical Tree Species Effects on Soil pH and Biotic Factors and the Consequences for Macroaggregate Dynamics" Forests 9, no. 4: 184. https://doi.org/10.3390/f9040184
APA StyleRussell, A. E., Kivlin, S. N., & Hawkes, C. V. (2018). Tropical Tree Species Effects on Soil pH and Biotic Factors and the Consequences for Macroaggregate Dynamics. Forests, 9(4), 184. https://doi.org/10.3390/f9040184