Susceptibility of Trees to Windthrow Storm Damage in Partially Harvested Complex-Structured Multi-Species Forests

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Methods
2.3. Analysis
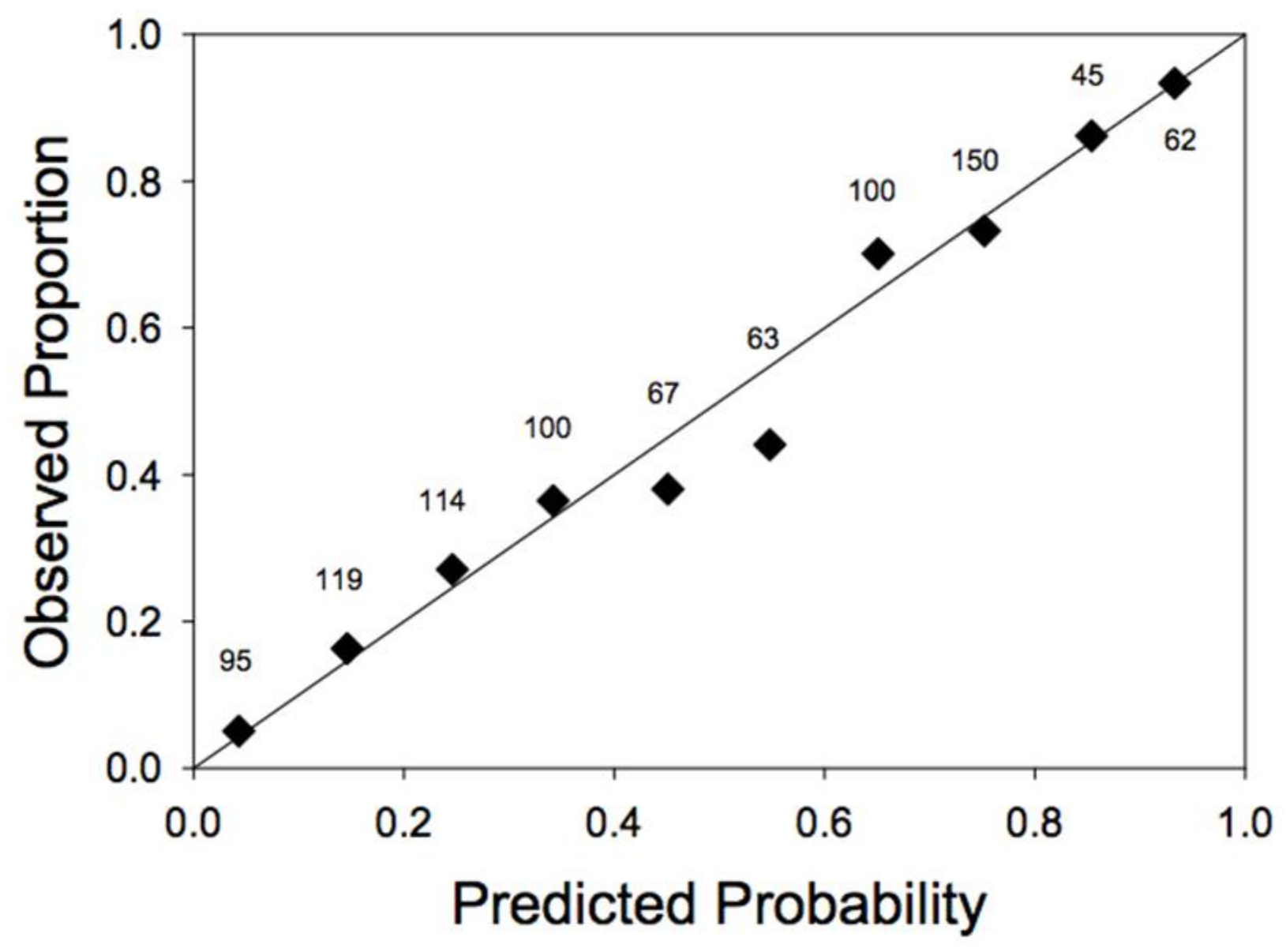
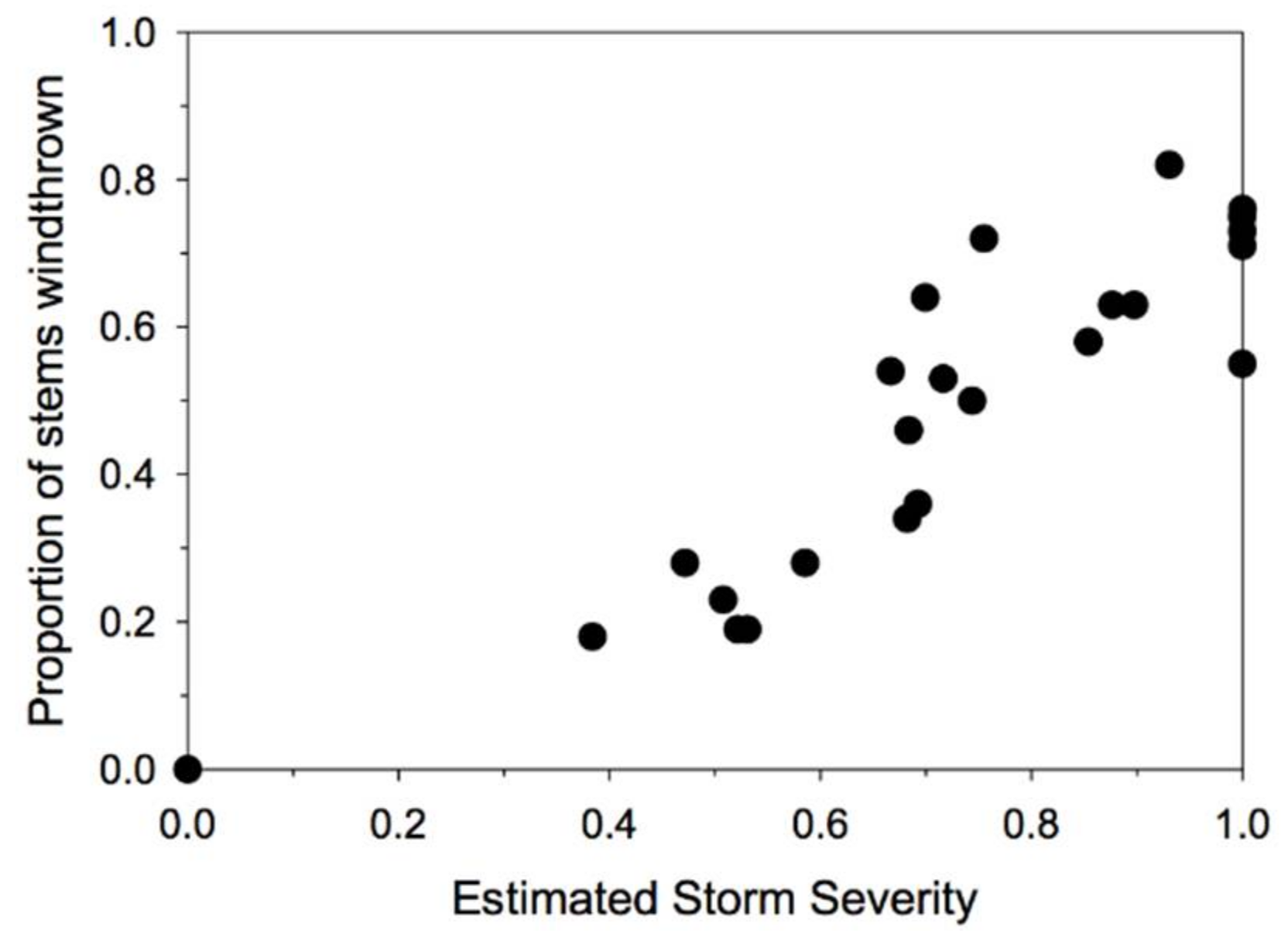
3. Results
3.1. Model Selection
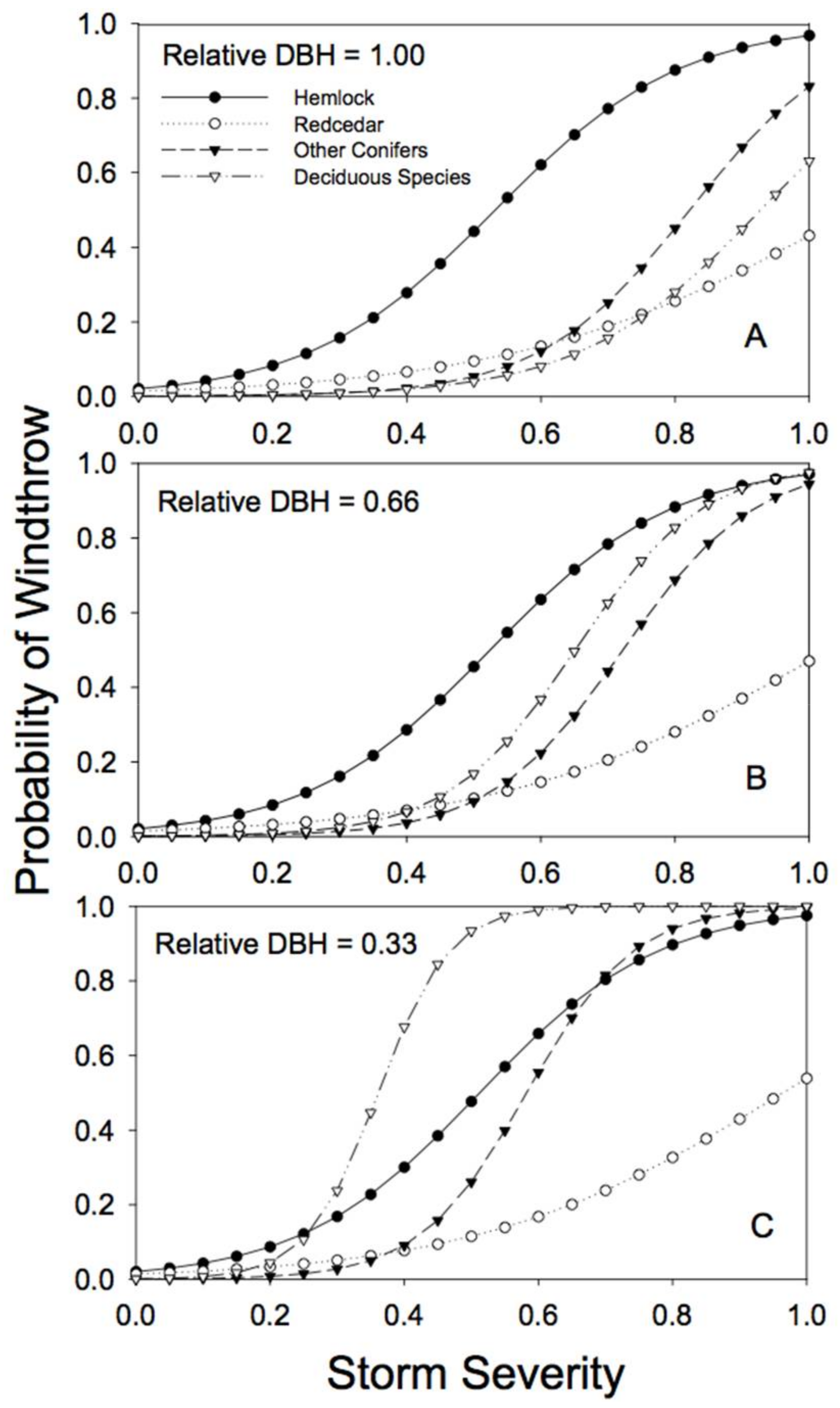
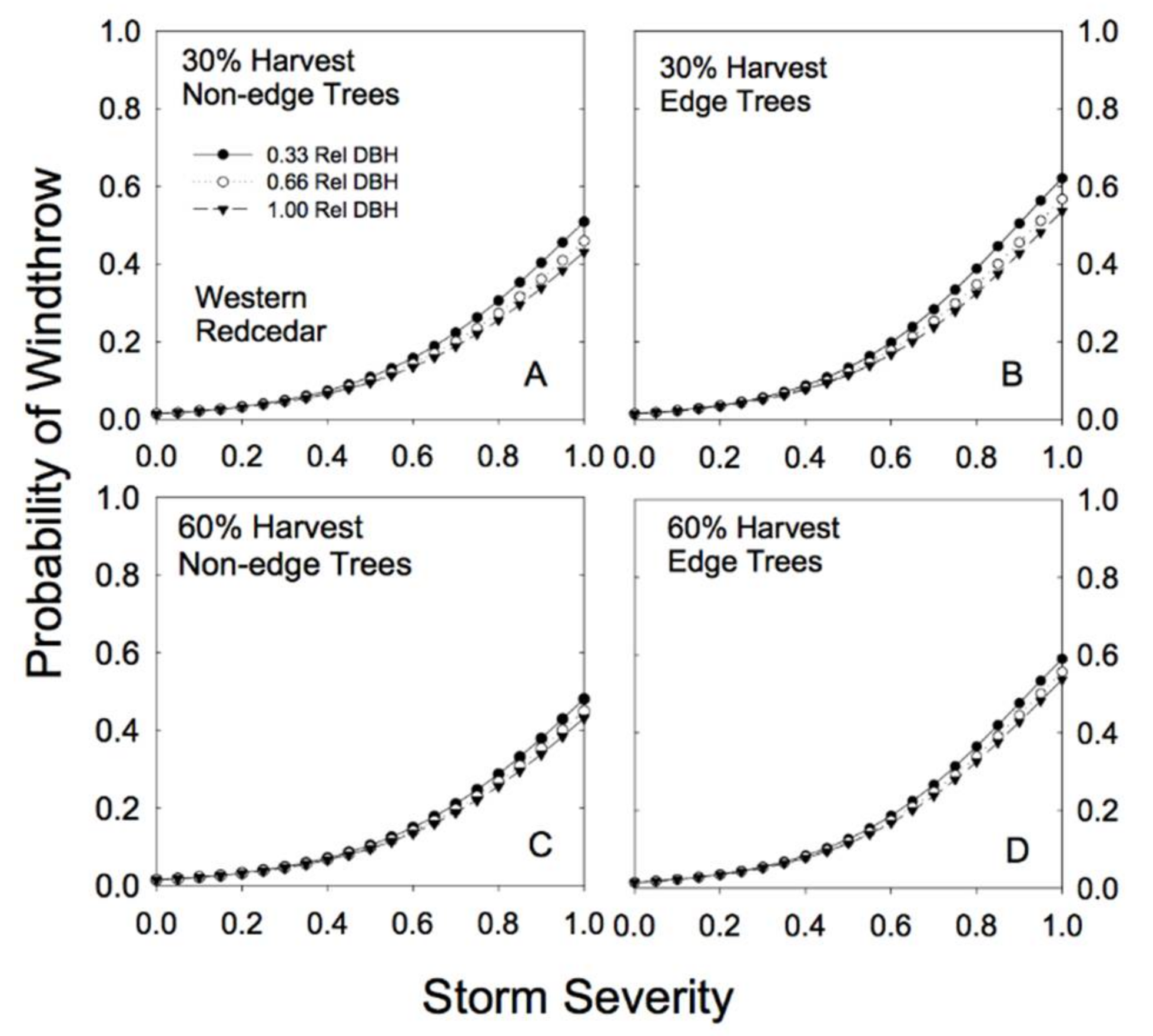
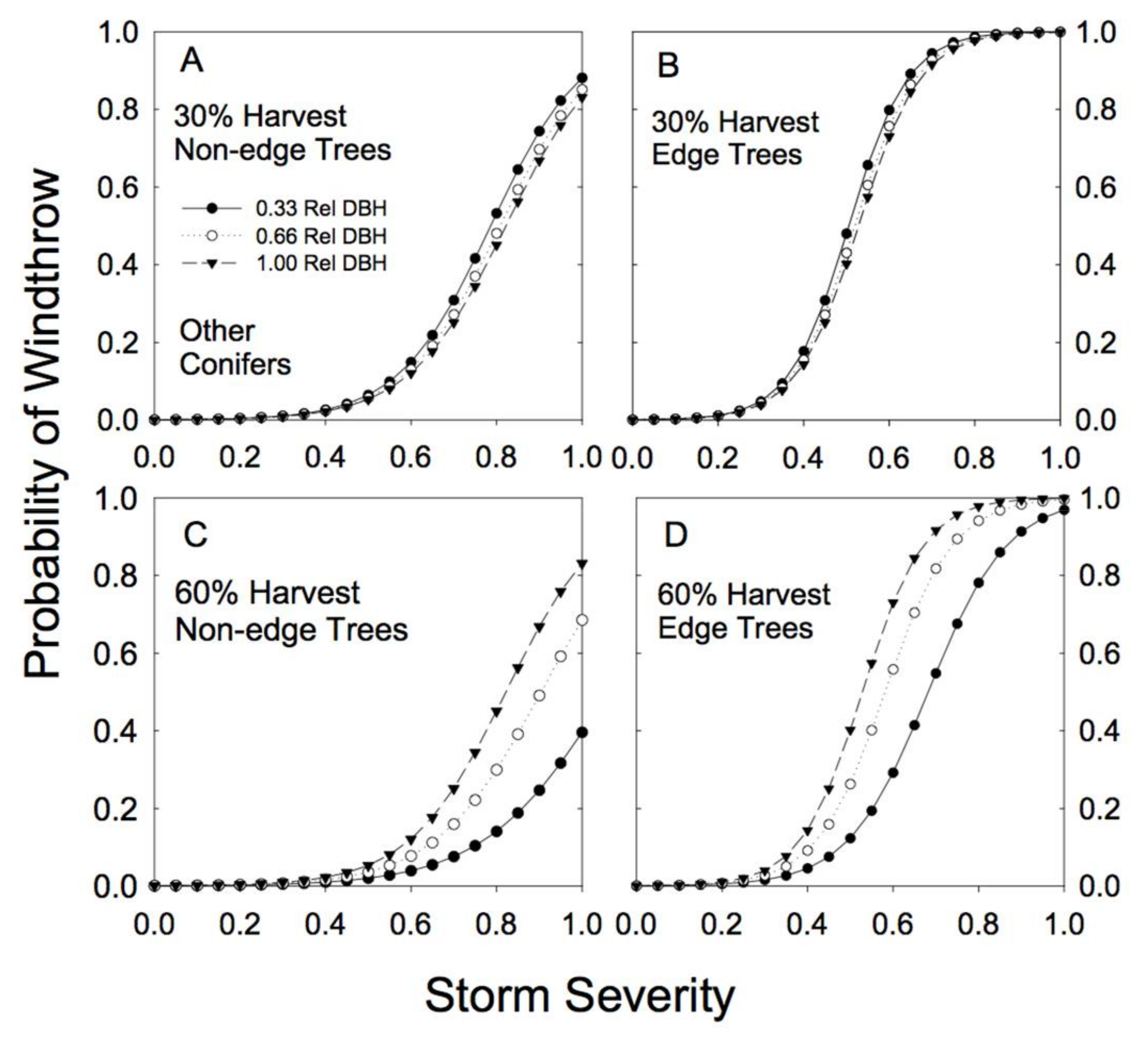
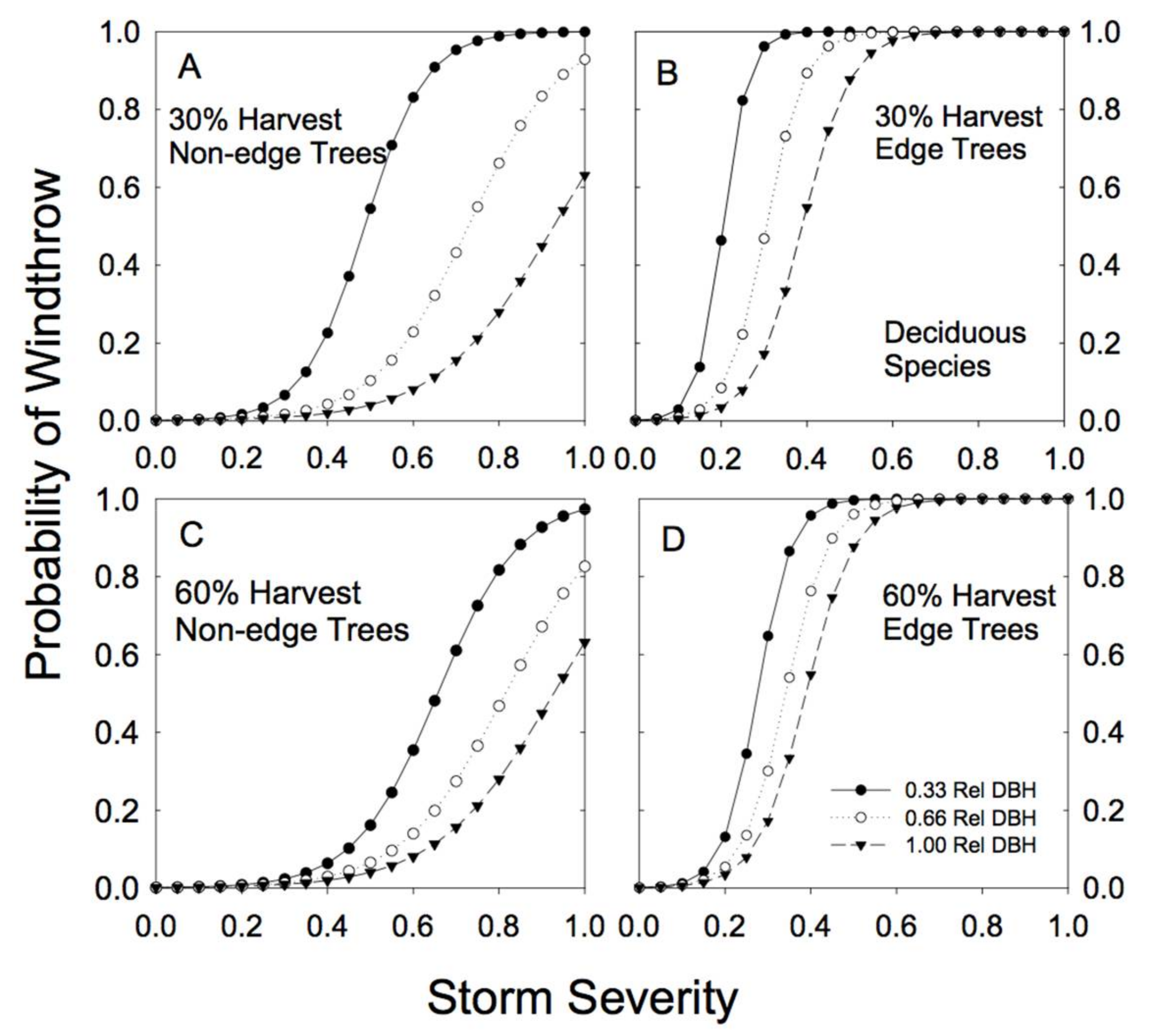
3.2. Windthrow Susceptibility in Unlogged Forests
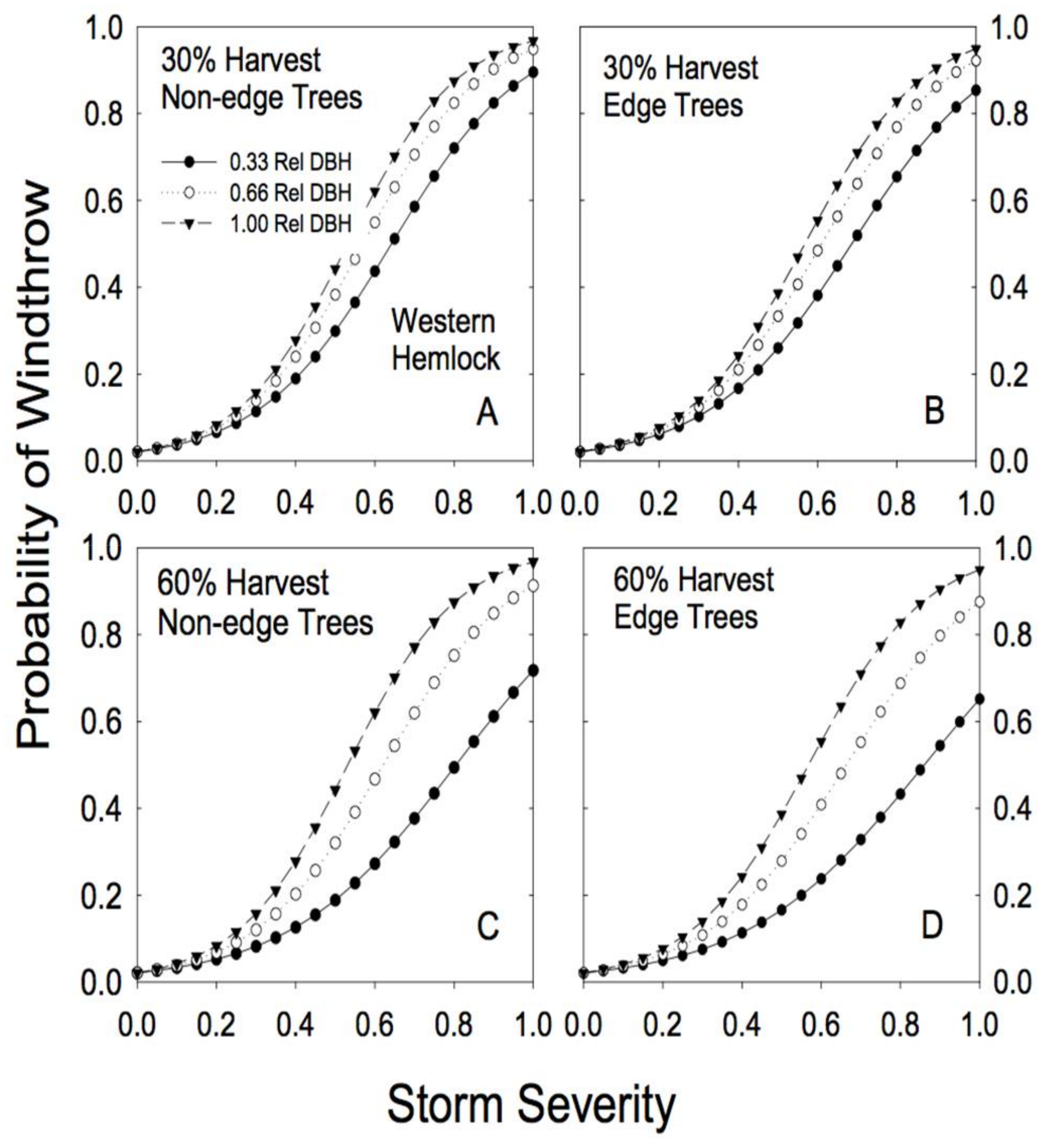
3.3. Windthrow Susceptibility in Partially Cut Forests
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BA Removal | Hemlock | Redcedar | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Plot | % | Mean | Min | Max | n | Mean | Min | Max | n |
| 1 | 0 | 26.3 | 13 | 44 | 19 | 24.6 | 12.8 | 39.4 | 5 |
| 2 | 0 | 23.1 | 12.5 | 36.5 | 26 | 20.9 | 15 | 30.8 | 6 |
| 3 | 0 | 24.6 | 11.2 | 64.2 | 43 | 37.3 | 14.8 | 66.8 | 7 |
| 4 | 0 | 26.3 | 18 | 39.5 | 4 | 18.7 | 10.8 | 37.6 | 10 |
| 5 | 0 | 33 | 16.9 | 47.7 | 20 | 0 | 0 | 0 | 0 |
| 6 | 0 | 23.6 | 12.5 | 36.8 | 19 | 25.2 | 15.4 | 40.3 | 5 |
| 7 | 22 | 27.8 | 10.6 | 55.2 | 49 | 24.9 | 10 | 47.3 | 18 |
| 8 | 22 | 32.7 | 11.8 | 50.5 | 36 | 20.5 | 10.2 | 40.5 | 8 |
| 9 | 24 | 30 | 10.6 | 62 | 56 | 25.7 | 12.2 | 41.1 | 7 |
| 10 | 33 | 33.8 | 17.8 | 84.8 | 17 | 30.6 | 22.4 | 39.4 | 3 |
| 11 | 45 | 25.9 | 10.9 | 42.6 | 22 | 22.1 | 11.2 | 37.9 | 11 |
| 12 | 47 | 31.2 | 12.8 | 42 | 16 | 34.5 | 17.3 | 42.5 | 10 |
| 13 | 53 | 32.8 | 10.8 | 52 | 25 | 20.5 | 11.2 | 26.3 | 9 |
| 14 | 56 | 33.4 | 16.1 | 46.8 | 16 | 35.1 | 28.1 | 44.9 | 6 |
| 15 | 59 | 32.6 | 13.8 | 49.9 | 17 | 21.8 | 10.3 | 43.4 | 22 |
| 16 | 62 | 25.9 | 14.1 | 57.5 | 27 | 15.4 | 15.4 | 15.4 | 1 |
| 17 | 63 | 21.4 | 10.6 | 42.4 | 27 | 17.3 | 10.8 | 27.4 | 13 |
| 18 | 63 | 42.2 | 13.1 | 83.2 | 20 | 53.4 | 46.1 | 61.5 | 3 |
| 19 | 64 | 36.1 | 23.9 | 57.2 | 9 | 19 | 12.2 | 37.6 | 10 |
| 20 | 69 | 33.6 | 17 | 41.7 | 11 | 21.1 | 13.6 | 29.3 | 5 |
| 21 | 70 | 19.4 | 10.5 | 38.8 | 34 | 18.8 | 10.6 | 26.9 | 2 |
| 22 | 72 | 28.9 | 10.5 | 85.5 | 45 | 44.7 | 28.2 | 54.9 | 7 |
| 23 | 74 | 35.8 | 18.9 | 48.1 | 12 | 24.4 | 20.9 | 27.8 | 4 |
| 24 | 76 | 25.6 | 10.6 | 36.7 | 22 | 23.1 | 11.3 | 34.2 | 11 |
| BA Removal | Other Conifers | Deciduous | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Plot | % | Mean | Min | Max | n | Mean | Min | Max | n |
| 1 | 0 | 40.3 | 36.4 | 44.1 | 2 | 31.8 | 31.8 | 31.8 | 1 |
| 2 | 0 | 34.9 | 28.5 | 41.8 | 7 | 0 | 0 | 0 | 0 |
| 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 4 | 0 | 43.1 | 37.9 | 48.2 | 2 | 31.5 | 24.7 | 39.2 | 5 |
| 5 | 0 | 29 | 22 | 36.3 | 5 | 0 | 0 | 0 | 0 |
| 6 | 0 | 0 | 0 | 0 | 0 | 19.5 | 19.5 | 19.5 | 1 |
| 7 | 22 | 41.6 | 22.5 | 66 | 14 | 0 | 0 | 0 | 0 |
| 8 | 22 | 52.7 | 35.3 | 70 | 2 | 40.7 | 31.5 | 48.1 | 4 |
| 9 | 24 | 39.3 | 30.2 | 52.1 | 6 | 26.7 | 20.3 | 33 | 7 |
| 10 | 33 | 40.5 | 31.9 | 46.2 | 5 | 0 | 0 | 0 | 0 |
| 11 | 45 | 32.1 | 20.8 | 48.9 | 10 | 26.4 | 17.6 | 45.4 | 5 |
| 12 | 47 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 13 | 53 | 37.6 | 24.5 | 53.7 | 9 | 30.6 | 24 | 36.3 | 8 |
| 14 | 56 | 38 | 36.5 | 39.4 | 2 | 24.5 | 17.9 | 31 | 2 |
| 15 | 59 | 35.8 | 18.1 | 57.3 | 3 | 0 | 0 | 0 | 0 |
| 16 | 62 | 33.1 | 29.4 | 36.7 | 2 | 0 | 0 | 0 | 0 |
| 17 | 63 | 0 | 0 | 0 | 0 | 31.4 | 20.6 | 41.2 | 4 |
| 18 | 63 | 10.5 | 10.5 | 10.5 | 1 | 0 | 0 | 0 | 0 |
| 19 | 64 | 34.2 | 28 | 42 | 7 | 30.7 | 20.4 | 43.4 | 5 |
| 20 | 69 | 29.6 | 26.8 | 33.7 | 4 | 0 | 0 | 0 | 0 |
| 21 | 70 | 0 | 0 | 0 | 0 | 24.5 | 24.5 | 24.5 | 1 |
| 22 | 72 | 28.8 | 23.8 | 34.7 | 3 | 0 | 0 | 0 | 0 |
| 23 | 74 | 33.8 | 26.5 | 39.8 | 4 | 27.9 | 27.9 | 27.9 | 1 |
| 24 | 76 | 28.7 | 24.4 | 33.3 | 8 | 0 | 0 | 0 | 0 |
References
- Gustafsson, L.; Baker, S.C.; Bauhus, J.; Beese, W.J.; Brodie, A.; Kouki, J.; Lindenmayer, D.B.; Lõhmus, A.; Pastur, G.M.; Messier, C.; et al. Retention forestry to maintain multifunctional forests: A world perspective. BioScience 2012, 62, 633–645. [Google Scholar] [CrossRef]
- Messier, C.; Puettmann, K.; Chazdon, R.; Andersson, K.P.; Angers, V.A.; Brotons, L.; Filotas, E.; Tittler, R.; Parrott, L.; Levin, S.A. From management to stewardship: Viewing forests as complex adaptive systems in an uncertain world. Conserv. Lett. 2015, 8, 368–377. [Google Scholar] [CrossRef] [Green Version]
- Nocentini, S.; Buttoud, G.; Ciancio, O.; Corona, P. Managing forests in a changing world: The need for a systemic approach. A review. For. Syst. 2017, 26, eR01. [Google Scholar] [CrossRef]
- Albrecht, A.; Hanewinkel, M.; Bauhus, J.; Kohnle, U. How does silviculture affect storm damage in forests of south-western Germany? Results from empirical modeling based on long-term observations. Eur. J. For. Res. 2012, 131, 229–247. [Google Scholar] [CrossRef]
- Mitchell, S.J. Wind as a natural disturbance agent in forests: A synthesis. Forestry 2013, 86, 147–157. [Google Scholar] [CrossRef]
- Maxwell, S.; Green, D.S.; Zhang, W. Identifying the determinants of windthrow damage in wildlife tree patches in the Boreal White and Black Spruce biogeoclimatic zone of northeastern British Columbia. J. Ecosyst. Manag. 2010, 10, 1–8. [Google Scholar]
- Moore, J.R.; Mitchell, S.J.; Maguire, D.A.; Quine, C.P. Wind damage in alternative silvicultural systems: Review and synthesis of previous studies. In Proceedings of the International Conference ‘Wind Effects on Trees’, Karlsruhe, Germany, 16–18 September 2003; pp. 191–198. [Google Scholar]
- Anyomi, K.A.; Ruel, J.-C. A multi-scale analysis of the effects of alternative silvicultural treatments on windthrow within balsam fir dominated stands. Can. J. For. Res. 2015, 45, 1739–1747. [Google Scholar] [CrossRef]
- Anyomi, K.A.; Mitchell, S.J.; Ruel, J.-C. Windthrow modelling in old-growth and multi-layered boreal forests. Ecol. Model. 2016, 327, 105–114. [Google Scholar] [CrossRef]
- Pukkala, T.; Laiho, O.; Lähde, E. Continuous cover management reduces wind damage. For. Ecol. Manag. 2016, 372, 120–127. [Google Scholar] [CrossRef]
- Coates, K.D. Windthrow damage two years after partial cutting of the Date Creek silvicultural systems study in the Interior Cedar-Hemlock forests of northwestern British Columbia. Can. J. For. Res. 1997, 27, 1695–1701. [Google Scholar]
- Canham, C.D.; Papaik, M.J.; Latty, E.F. Interspecific variation in susceptibility to windthrow as a function of tree size and storm severity for northern temperate tree species. Can. J. For. Res. 2001, 31, 1–10. [Google Scholar] [CrossRef]
- Canham, C.D.; Thompson, J.; Zimmerman, J.K.; Uriarte, M. Variation in susceptibility to hurricane damage as a function of storm intensity in Puerto Rican tree species. Biotropica 2010, 42, 87–94. [Google Scholar] [CrossRef]
- Coates, K.D.; Banner, A.; Steventon, J.D.; LePage, P.; Bartemucci, P. The Date Creek Silvicultural Systems Study in the Interior Cedar-Hemlock Forests of Northwestern British Columbia: Overview and Treatment Summaries; Land Management Handbook 38; B.C. Ministry of Forests: Victoria, BC, Canada, 1997.
- Banner, A.; MacKenzie, W.; Haeussler, S.; Thomson, S.; Pojar, J.; Trowbridge, R. A Field Guide to Site Identification and Interpretation for the Prince Rupert Forest Region; Land Management Handbook 26; B.C. Ministry of Forests: Victoria, BC, Canada, 1993.
- Pojar, J.; Klinka, K.; Meidinger, D.V. Biogeoclimatic ecosystem classification in British Columbia. For. Ecol. Manag. 1987, 22, 119–154. [Google Scholar] [CrossRef]
- Meidinger, D.; Pojar, J. Ecosystems of British Columbia. Special Report Series 6; B.C. Ministry of Forests: Victoria, BC, Canada, 1991.
- Agriculture Canada Expert Committee on Soil Survey. The Canadian System of Soil Classification; Agriculture and Agri-Food Canada Publication No. 1646; Agriculture Canada Publication: Ottawa, ON, Canada, 1987. [Google Scholar]
- Scott, R.E.; Mitchell, S.J. Empirical modelling of windthrow risk in partially harvested stands using tree, neighbourhood, and stand attributes. For. Ecol. Manag. 2005, 218, 193–209. [Google Scholar] [CrossRef]
- Valinger, E.; Fridman, J. Factors affecting the probability of windthrow at stand level as a result of Gudrun winter storm in southern Sweden. For. Ecol. Manag. 2011, 262, 398–403. [Google Scholar] [CrossRef]
- Huggard, D.J.; Klenner, W.; Vyse, A. Windthrow following four harvest treatments in an Engelmann spruce—Subalpine fir forest in southern interior British Columbia, Canada. Can. J. For. Res. 1999, 29, 1547–1556. [Google Scholar] [CrossRef]
- Kamimura, K.; Gardiner, B.; Dupont, S.; Guyon, D.; Meredieu, C. Mechanistic and statistical approaches to predicting wind damage to individual maritime pine (Pinus pinaster) trees in forests. Can. J. For. Res. 2015, 46, 88–100. [Google Scholar] [CrossRef]
- Ruel, J.-C.; Raymond, P.; Pineau, M. Windthrow after shelterwood cutting in balsam fir stands. North. J. Appl. For. 2003, 20, 5–13. [Google Scholar]
- Coutts, M.P.; Grace, J. (Eds.) Wind and Trees; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Ruel, J.-C.; Quine, C.P.; Meunier, S.; Suarez, J. Estimating windthrow risk in balsam fir stands with the ForestGales model. For. Chron. 2000, 76, 329–337. [Google Scholar] [CrossRef]
- Gardiner, B.; Byrne, K.; Hale, S.; Kamimura, K.; Mitchell, S.J.; Peltola, H.; Ruel, J.C. A review of mechanistic modelling of wind damage risk to forests. Forestry 2008, 81, 447–463. [Google Scholar] [CrossRef]
- Seidl, R.; Rammer, W.; Blennow, K. Simulating wind disturbance impacts on forest landscapes: Tree-level heterogeneity matters. Environ. Model. Softw. 2014, 51, 1–11. [Google Scholar] [CrossRef]
- Anyomi, K.A.; Mitchell, S.J.; Perera, A.H.; Ruel, J.C. Windthrow dynamics in boreal Ontario: A simulation of the vulnerability of several stand types across a range of wind speeds. Forests 2017, 8, 233. [Google Scholar] [CrossRef]
- Hale, S.E.; Gardiner, B.; Peace, A.; Nicoll, B.; Taylor, P.; Pizzirani, S. Comparison and validation of three versions of a forest wind risk model. Environ. Model. Softw. 2015, 68, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, S.J.; Hailemariam, T.; Kulis, Y. Empirical modeling of cutblock edge windthrow risk on Vancouver Island, Canada, using stand level information. For. Ecol. Manag. 2001, 154, 117–130. [Google Scholar] [CrossRef]







| Model | DBH | AIC | DBH | AIC |
|---|---|---|---|---|
| 1 | Absolute | 1033 | Relative | 1023.4 |
| 1a | Absolute | 1027.5 | Relative | 1018.4 |
| 2 | Absolute | 1039.9 | Relative | 1039.9 |
| 3 | Absolute | 1035.5 | Relative | 1025.8 |
| 3a | Absolute | 1146.8 | Relative | 1017.4 |
| 4 | Absolute | 1027.6 | Relative | 1015.5 |
| 4a | Absolute | 1147.7 | Relative | 1009.3 |
| Tree Species | as | bs | cnon-edge | cedge | hs |
|---|---|---|---|---|---|
| Western hemlock | −3.85 | −0.034 | 7.24 | 6.78 | 0.68 |
| −4.06 to −3.67 | −0.079 to 0.029 | 6.87 to 7.59 | 5.88 to 7.73 | 0.57 to 0.80 | |
| Western redcedar | −4.24 | −0.093 | 3.96 | 4.38 | 0.082 |
| −4.61 to −3.83 | −0.22 to 0.04 | 3.47 to 4.49 | 2.70 to 5.75 | −0.14 to 0.35 | |
| Other Conifers | −7.36 | −0.31 | 8.95 | 13.92 | 0.9 |
| −7.90 to −6.85 | −0.43 to −0.17 | 8.20 to 9.66 | 11.51 to 16.37 | 0.68 to 1.19 | |
| Deciduous | −6.88 | −0.85 | 7.42 | 17.69 | 0.9 |
| −7.86 to −6.14 | −1.01 to −0.66 | 6.43 to 8.25 | 12.71 to 24.69 | 0.55 to 1.32 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coates, K.D.; Hall, E.C.; Canham, C.D. Susceptibility of Trees to Windthrow Storm Damage in Partially Harvested Complex-Structured Multi-Species Forests. Forests 2018, 9, 199. https://doi.org/10.3390/f9040199
Coates KD, Hall EC, Canham CD. Susceptibility of Trees to Windthrow Storm Damage in Partially Harvested Complex-Structured Multi-Species Forests. Forests. 2018; 9(4):199. https://doi.org/10.3390/f9040199
Chicago/Turabian StyleCoates, K. David, Erin C. Hall, and Charles D. Canham. 2018. "Susceptibility of Trees to Windthrow Storm Damage in Partially Harvested Complex-Structured Multi-Species Forests" Forests 9, no. 4: 199. https://doi.org/10.3390/f9040199
APA StyleCoates, K. D., Hall, E. C., & Canham, C. D. (2018). Susceptibility of Trees to Windthrow Storm Damage in Partially Harvested Complex-Structured Multi-Species Forests. Forests, 9(4), 199. https://doi.org/10.3390/f9040199