Seed Dispersal, Microsites or Competition—What Drives Gap Regeneration in an Old-Growth Forest? An Application of Spatial Point Process Modelling

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.3. Statistical Analysis
3. Results
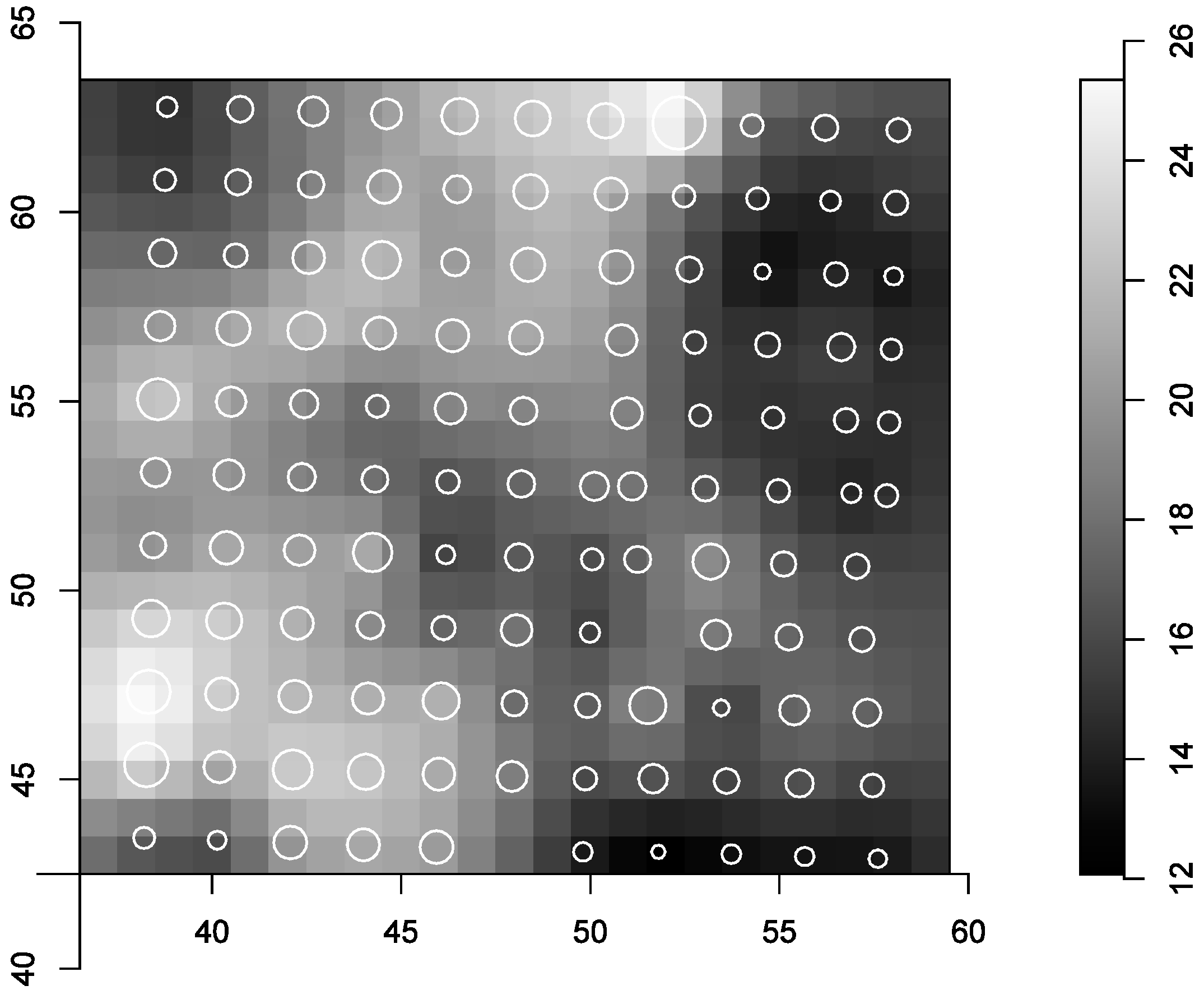
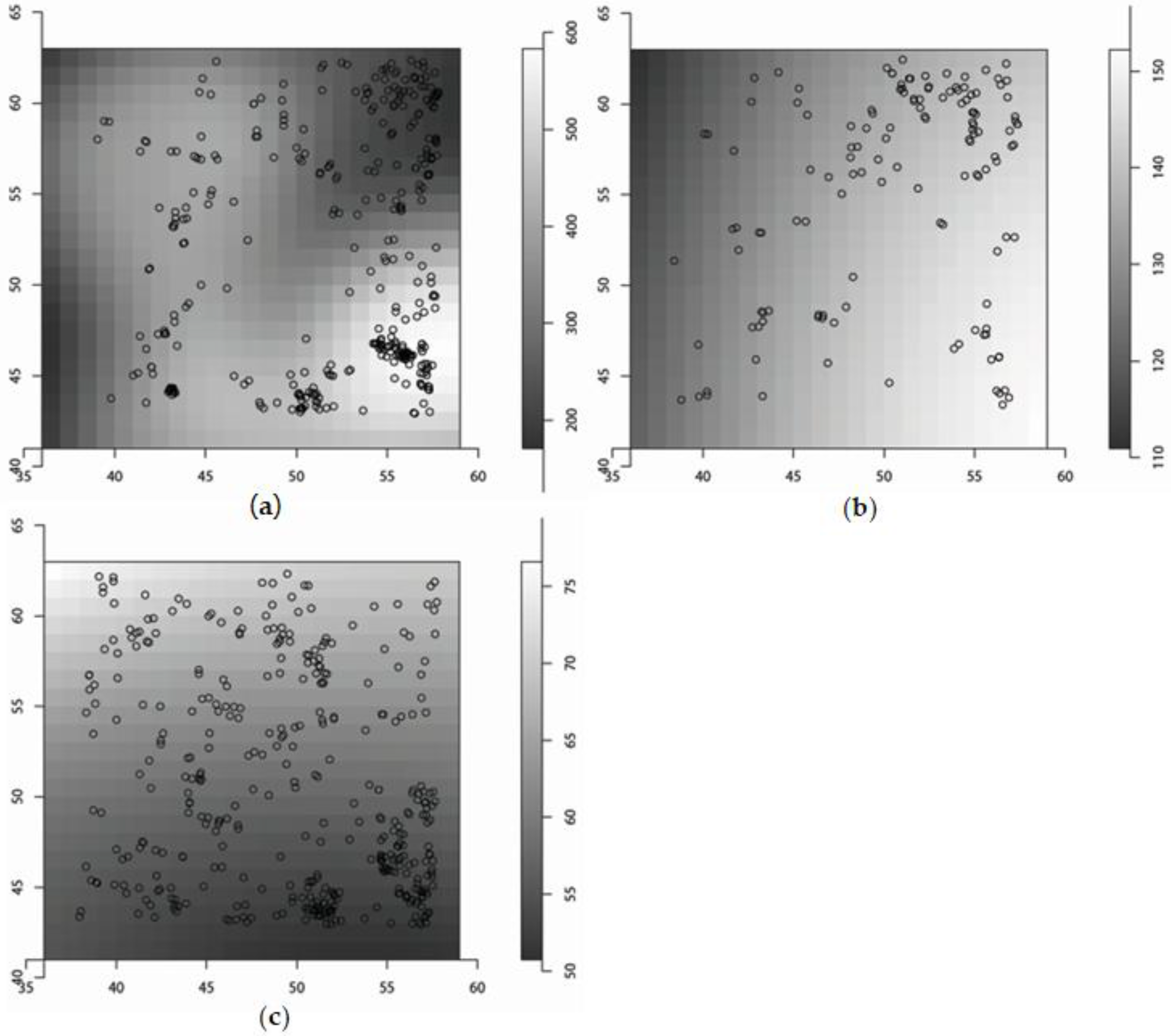
3.1. The Spatial Covariates
3.2. Parameter Estimates
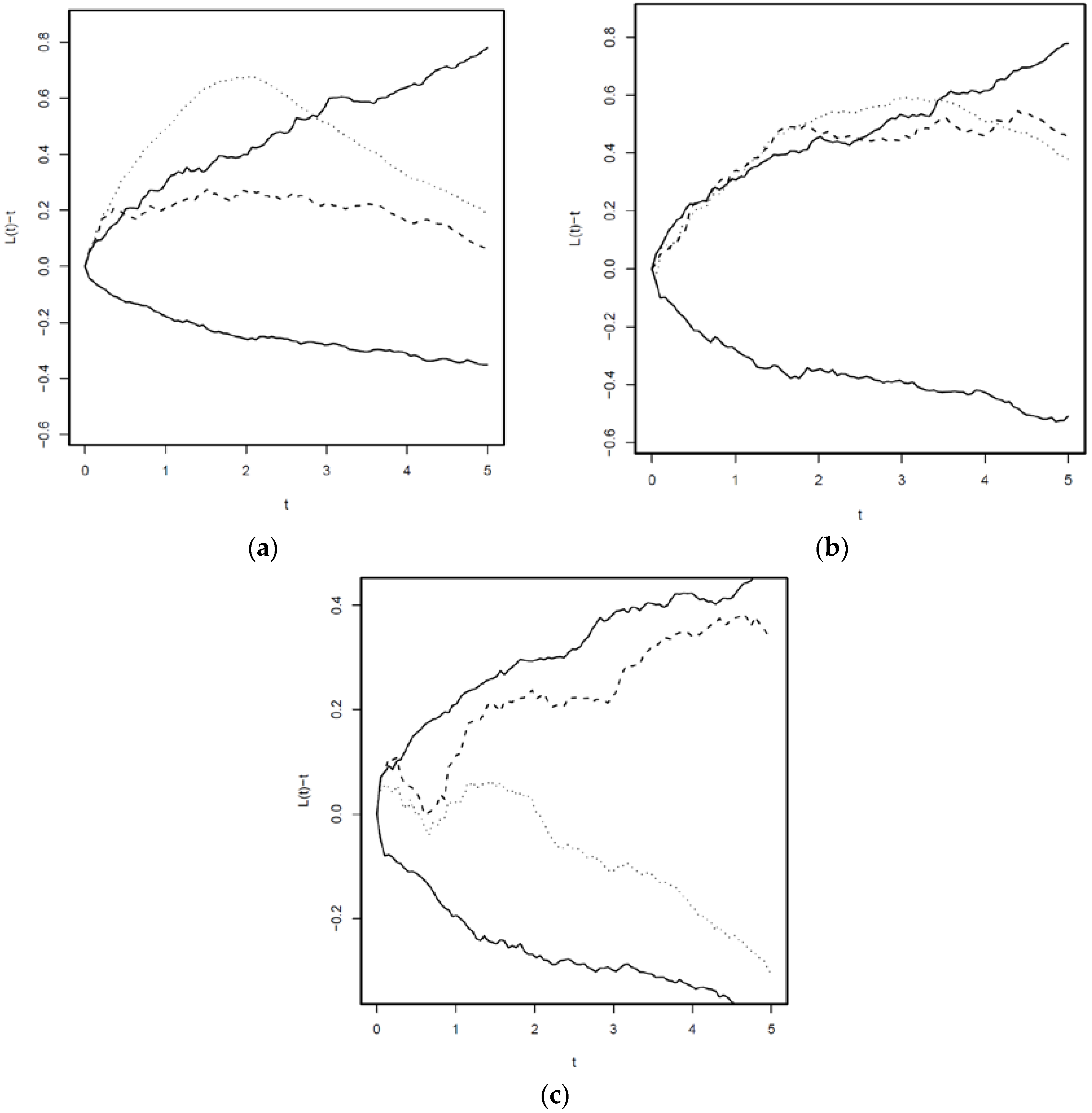
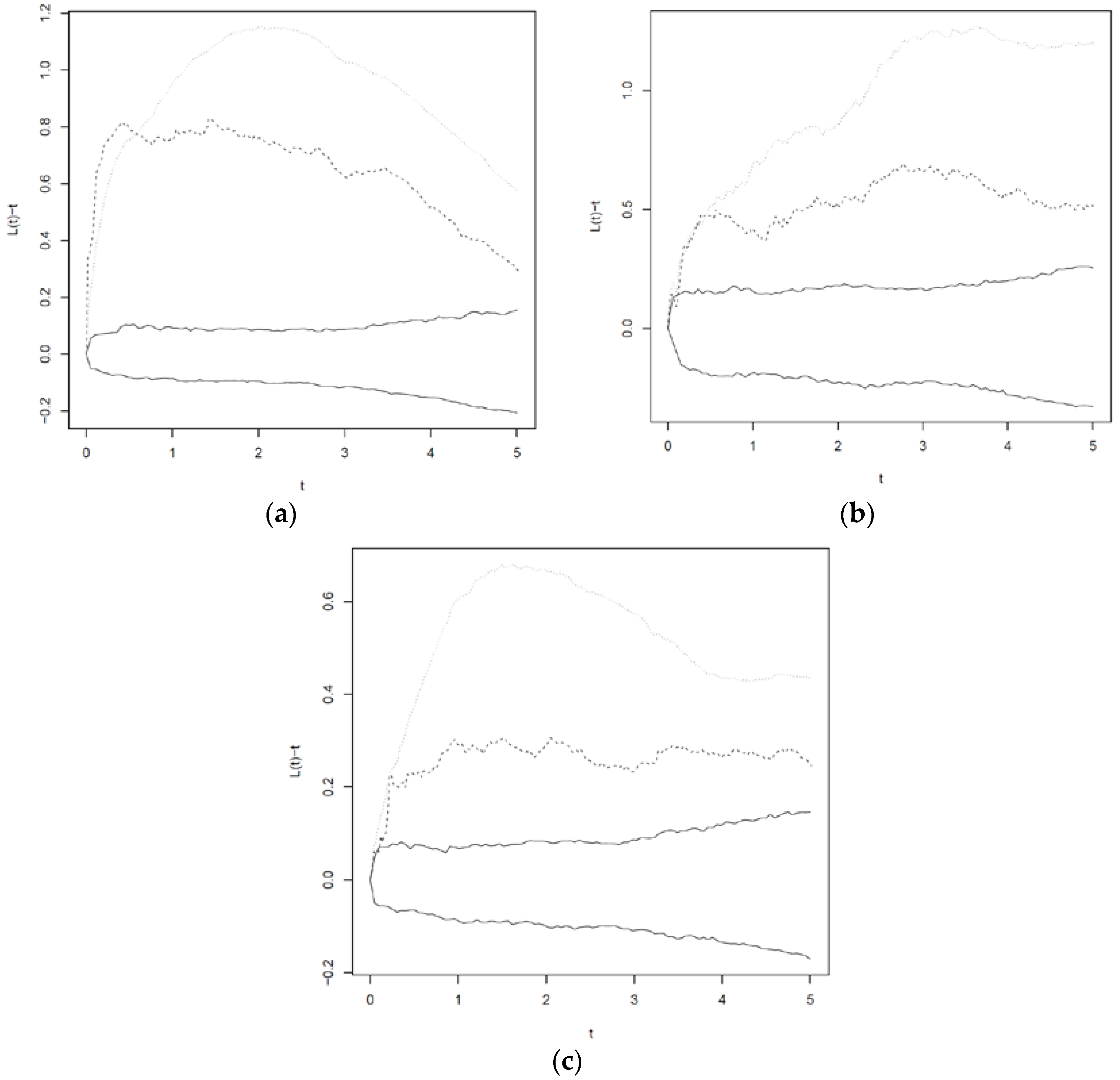
3.3. Cross L—Functions
4. Discussion
4.1. Quantifying Pattern Generating Factors with a Poisson Cluster Model
4.2. The Role of Seed Dispersal Versus Environmental Heterogeneity
4.3. Competition and Spatial Segregation
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Appendix A

References
- Watt, A.S. Pattern and Process in the Plant Community. J. Ecol. 1947, 35, 1–22. [Google Scholar] [CrossRef]
- Harper, J.L. Population biology of plants. In Population Biology of Plants; Academic Press: Waltham, MA, USA, 1977. [Google Scholar]
- Law, R.; Illian, J.B.; Burslem, D.F.R.P.; Gratzer, G.; Gunatilleke, C.V.S.; Gunatilleke, I.A.U.N. Ecological information from spatial patterns of plants: Insights from point process theory. J. Ecol. 2009, 97, 616–628. [Google Scholar] [CrossRef]
- Brown, C.; Illian, J.B.; Burslem, D.F.R.P.; Sandhu, L. Success of spatial statistics in determining underlying process in simulated plant communities. J. Ecol. 2016, 104, 106–172. [Google Scholar] [CrossRef]
- Diggle, P. Statistical Analysis of Spatial Point Patterns; Arnold: London, UK, 2003; ISBN 978-0-340-74070-5. [Google Scholar]
- Illian, D.J.; Penttinen, P.A.; Stoyan, D.H.; Stoyan, D. Statistical Analysis and Modelling of Spatial Point Patterns; John Wiley & Sons: Hoboken, NJ, USA, 2008; ISBN 978-0-470-72515-3. [Google Scholar]
- Møller, J.; Waagepetersen, R.P. Modern Statistics for Spatial Point Processes. Scand. J. Stat. 2007, 34, 643–684. [Google Scholar] [CrossRef]
- Møller, J.; Waagepetersen, R. Some Recent Developments in Statistics for Spatial Point Patterns. Annu. Rev. Stat. Its Appl. 2017, 4, 317–342. [Google Scholar] [CrossRef]
- Getzin, S.; Wiegand, T.; Wiegand, K.; He, F. Heterogeneity influences spatial patterns and demographics in forest stands. J. Ecol. 2008, 96, 807–820. [Google Scholar] [CrossRef]
- Zhu, Y.; Getzin, S.; Wiegand, T.; Ren, H.; Ma, K. The Relative Importance of Janzen-Connell Effects in Influencing the Spatial Patterns at the Gutianshan Subtropical Forest. PLoS ONE 2013, 8, e74560. [Google Scholar] [CrossRef] [PubMed]
- Guochun, S.; Mingjian, Y.; Xinsheng, H.; Xiangcheng, M.; Haibao, R.; I-Fang, S.; Keping, M. Species-area relationships explained by the joint effects of dispersal limitation and habitat heterogeneity. Ecology 2009, 90, 3033–3041. [Google Scholar]
- Lin, Y.-C.; Chang, L.-W.; Yang, K.-C.; Wang, H.-H.; Sun, I.-F. Point patterns of tree distribution determined by habitat heterogeneity and dispersal limitation. Oecologia 2011, 165, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, E.; Martínez, I.; Getzin, S.; Moloney, K.A.; Wiegand, T. An evaluation of the state of spatial point pattern analysis in ecology. Ecography 2016, 39, 1042–1055. [Google Scholar] [CrossRef]
- Motta, R.; Garbarino, M.; Berretti, R.; Meloni, F.; Nosenzo, A.; Vacchiano, G. Development of old-growth characteristics in uneven-aged forests of the Italian Alps. Eur. J. For. Res. 2015, 134, 19–31. [Google Scholar] [CrossRef]
- Burrascano, S.; Keeton, W.S.; Sabatini, F.M.; Blasi, C. Commonality and variability in the structural attributes of moist temperate old-growth forests: A global review. For. Ecol. Manag. 2013, 291, 458–479. [Google Scholar] [CrossRef]
- Keeton, W.S.; Chernyavskyy, M.; Gratzer, G.; Main-Knorn, M.; Shpylchak, M.; Bihun, Y. Structural characteristics and aboveground biomass of old-growth spruce–fir stands in the eastern Carpathian mountains, Ukraine. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2010, 144, 148–159. [Google Scholar] [CrossRef]
- Kern, C.C.; Burton, J.I.; Raymond, P.; D’Amato, A.W.; Keeton, W.S.; Royo, A.A.; Walters, M.B.; Webster, C.R.; Willis, J.L. Challenges facing gap-based silviculture and possible solutions for mesic northern forests in North America. For. Int. J. For. Res. 2017, 90, 4–17. [Google Scholar] [CrossRef]
- Canham, C.D.; Finzi, A.C.; Pacala, S.W.; Burbank, D.H. Causes and consequences of resource heterogeneity in forests: Interspecific variation in light transmission by canopy trees. Can. J. For. Res. 1994, 24, 337–349. [Google Scholar] [CrossRef]
- Latif, Z.A.; Blackburn, G.A. The effects of gap size on some microclimate variables during late summer and autumn in a temperate broadleaved deciduous forest. Int. J. Biometeorol. 2010, 54, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Muscolo, A.; Bagnato, S.; Sidari, M.; Mercurio, R. A review of the roles of forest canopy gaps. J. For. Res. 2014, 25, 725–736. [Google Scholar] [CrossRef]
- Hubbell, S.P.; Foster, R.B.; O’Brien, S.T.; Harms, K.E.; Condit, R.; Wechsler, B.; Wright, S.J.; Lao, S.L. de Light-Gap Disturbances, Recruitment Limitation, and Tree Diversity in a Neotropical Forest. Science 1999, 283, 554–557. [Google Scholar] [CrossRef] [PubMed]
- Hubbell, S.P. Neutral Theory and the Evolution of Ecological Equivalence. Ecology 2006, 87, 1387–1398. [Google Scholar] [CrossRef]
- Bagchi, R.; Henrys, P.A.; Brown, P.E.; Burslem, D.F.R.P.; Diggle, P.J.; Gunatilleke, C.V.S.; Gunatilleke, I.A.U.N.; Kassim, A.R.; Law, R.; Noor, S.; et al. Spatial patterns reveal negative density dependence and habitat associations in tropical trees. Ecology 2011, 92, 1723–1729. [Google Scholar] [CrossRef] [PubMed]
- Uriarte, M.; Muscarella, R.; Zimmerman, J.K. Environmental heterogeneity and biotic interactions mediate climate impacts on tropical forest regeneration. Glob. Chang. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Brokaw, N.; Busing, R.T. Niche versus chance and tree diversity in forest gaps. Trends Ecol. Evol. 2000, 15, 183–188. [Google Scholar] [CrossRef]
- Nakashizuka, T. Species coexistence in temperate, mixed deciduous forests. Trends Ecol. Evol. 2001, 16, 205–210. [Google Scholar] [CrossRef]
- Gray, A.N.; Spies, T.A. Microsite Controls on Tree Seedling Establishment in Conifer Forest Canopy Gaps. Ecology 1997, 78, 2458–2473. [Google Scholar] [CrossRef]
- Szewczyk, J.; Szwagrzyk, J. Tree regeneration on rotten wood and on soil in old-growth stand. Vegetatio 1996, 122, 37–46. [Google Scholar] [CrossRef]
- Brang, P. Early seedling establishment of Picea abies in small forest gaps in the Swiss Alps. Can. J. For. Res. 1998, 28, 626–639. [Google Scholar] [CrossRef]
- Titus, J.H.; del Moral, R. Seedling establishment in different microsites on Mount St. Helens, Washington, USA. Plant Ecol. 1998, 134, 13–26. [Google Scholar] [CrossRef]
- Gratzer, G.; Rai, P.B.; Glatzel, G. The influence of the bamboo Yushania microphylla on regeneration of Abies densa in central Bhutan. Can. J. For. Res. 1999, 29, 1518–1527. [Google Scholar] [CrossRef]
- Pröll, G.; Darabant, A.; Gratzer, G.; Katzensteiner, K. Unfavourable microsites, competing vegetation and browsing restrict post-disturbance tree regeneration on extreme sites in the Northern Calcareous Alps. Eur. J. For. Res. 2015, 134, 293–308. [Google Scholar] [CrossRef]
- Kathke, S.; Bruelheide, H. Interaction of gap age and microsite type for the regeneration of Picea abies. For. Ecol. Manag. 2010, 259, 1597–1605. [Google Scholar] [CrossRef]
- Kupferschmid, A.D.; Bugmann, H. Effect of microsites, logs and ungulate browsing on Picea abies regeneration in a mountain forest. For. Ecol. Manag. 2005, 205, 251–265. [Google Scholar] [CrossRef]
- Van Couwenberghe, R.; Collet, C.; Lacombe, E.; Pierrat, J.-C.; Gégout, J.-C. Gap partitioning among temperate tree species across a regional soil gradient in windstorm-disturbed forests. For. Ecol. Manag. 2010, 260, 146–154. [Google Scholar] [CrossRef]
- Christie, D.A.; Armesto, J.J. Regeneration Microsites and Tree Species Coexistence in Temperate Rain Forests of Chiloé Island, Chile. J. Ecol. 2003, 91, 776–784. [Google Scholar] [CrossRef]
- Grubb, P. The maintenance of species-richness in plant communities: The importance of the regeneration niche. Biol. Rev. 1977, 52, 107–145. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A. Handbook of Spatial Point-Pattern Analysis in Ecology; CRC Press: Boca Raton, FL, USA, 2013; ISBN 978-1-4200-8255-5. [Google Scholar]
- Zukrigl, K.; Eckhart, G.; Nather, J. Standortskundliche und Waldbauliche Untersuchungen in Urwaldresten der Niederösterreichischen Kalkalpen. Mitt FBVA 62; Österreichischer Agrarverlag: Vienna, Austria, 1963. [Google Scholar]
- Willner, W.; Grabherr, G. Die Wälder und Gebüsche Österreichs; Spektrum Akademischer Verlag: Heidelberg, Berlin, 2007; ISBN 978-3-8274-1892-0. [Google Scholar]
- Schrempf, W. Waldbauliche Untersuchungen im Fichten-Tannen-Buchen-Urwald Rothwald und in Urwald-Folge-Beständen. Ph.D. Thesis, Universität für Bodenkultur, Wien, Austria, 1985. [Google Scholar]
- Runkle, J.R. Guidelines and Sample Protocol for Sampling Forest Gaps; General Technical Report PNW-GTR-283; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1992; Volume 283.
- Delta-T Devices Ltd. Hemiview User Manual 2.1; Delta-T Devices: Cambridge, UK, 1998. [Google Scholar]
- Rich, P.M. Characterizing plant canopies with hemispherical photographs. Remote Sens. Rev. 1990, 5, 13–29. [Google Scholar] [CrossRef]
- Ribbens, E.; Silander, J.A.; Pacala, S.W. Seedling Recruitment in Forests: Calibrating Models to Predict Patterns of Tree Seedling Dispersion. Ecology 1994, 75, 1794–1806. [Google Scholar] [CrossRef]
- Kutter, M.; Georg, G. Current methods for estimation of seed dispersal of trees—An example of seed dispersal of Picea abies, Abiel alba and Fagus sylvatica. Aust. J. For. Sci. 2006, 123, 103–120. [Google Scholar]
- Baddeley, A.J.; Møller, J.; Waagepetersen, R. Non- and semi-parametric estimation of interaction in inhomogeneous point patterns. Stat. Neerlandica 2000, 54, 329–350. [Google Scholar] [CrossRef]
- Waagepetersen, R.P. An Estimating Function Approach to Inference for Inhomogeneous Neyman–Scott Processes. Biometrics 2007, 63, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Splechtna, B.E.; Gratzer, G.; Black, B.A.; Franklin, J. Disturbance history of a European old-growth mixed-species forest—A spatial dendro-ecological analysis. J. Veg. Sci. 2005, 16, 511–522. [Google Scholar]
- Stoyan, D.; Penttinen, A. Recent Applications of Point Process Methods in Forestry Statistics. Stat. Sci. 2000, 15, 61–78. [Google Scholar]
- Bagchi, R.; Illian, J.B. A method for analysing replicated point patterns in ecology. Methods Ecol. Evol. 2015, 6, 482–490. [Google Scholar] [CrossRef]
- Simon, A.; Gratzer, G.; Sieghardt, M. The influence of windthrow microsites on tree regeneration and establishment in an old growth mountain forest. For. Ecol. Manag. 2011, 262, 1289–1297. [Google Scholar] [CrossRef]
- Punchi-Manage, R.; Wiegand, T.; Wiegand, K.; Getzin, S.; Gunatilleke, C.V.S.; Gunatilleke, I.A.U.N. Effect of spatial processes and topography on structuring species assemblages in a Sri Lankan dipterocarp forest. Ecology 2014, 95, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Szwagrzyk, J.; Szewczyk, J.; Bodziarczyk, J. Dynamics of seedling banks in beech forest: Results of a 10-year study on germination, growth and survival. For. Ecol. Manag. 2001, 141, 237–250. [Google Scholar] [CrossRef]
- Gillman, L.N.; Wright, S.D.; Ogden, J. Response of forest tree seedlings to simulated litterfall damage. Plant Ecol. 2003, 169, 53–60. [Google Scholar] [CrossRef]
- Gillman, L.N.; Ogden, J. Physical damage by litterfall to canopy tree seedlings in two temperate New Zealand forests. J. Veg. Sci. 2001, 12, 671–676. [Google Scholar] [CrossRef]
- Simard, M.-J.; Bergeron, Y.; Sirois, L. Substrate and litterfall effects on conifer seedling survivorship in southern boreal stands of Canada. Can. J. For. Res. 2003, 33, 672–681. [Google Scholar] [CrossRef]
- Hunziker, U.; Brang, P. Microsite patterns of conifer seedling establishment and growth in a mixed stand in the Southern Alps. For. Ecol. Manag. 2005, 210, 67–79. [Google Scholar] [CrossRef]
- Peterson, C.J.; Campbell, J.E. Microsite Differences and Temporal Change in Plant Communities of Treefall Pits and Mounds in an Old-Growth Forest. Bull. Torrey Bot. Club 1993, 120, 451–460. [Google Scholar] [CrossRef]
- Bazzigher, G. Der schwarze Schneeschimmel der Koniferen [Herpotrichia juniperi (Duby) Petrak und Herpotrichia Coulteri (Peck) Bose). For. Pathol. 1976, 6, 109–122. [Google Scholar] [CrossRef]
- Baier, R.; Meyer, J.; Göttlein, A. Regeneration niches of Norway spruce (Picea abies [L.] Karst.) saplings in small canopy gaps in mixed mountain forests of the Bavarian Limestone Alps. Eur. J. For. Res. 2007, 126, 11–22. [Google Scholar] [CrossRef]
- Hornberg, G.; Ohlson, M.; Zackrisson, O. Influence of bryophytes and microrelief conditions on Picea abies seed regeneration patterns in boreal old-growth swamp forests. Can. J. For. Res. 1997, 27, 1015–1023. [Google Scholar] [CrossRef]
- Tognetti, R.; Minotta, G.; Pinzauti, S.; Michelozzi, M.; Borghetti, M. Acclimation to changing light conditions of long-term shade-grown beech (Fagus sylvatica L.) seedlings of different geographic origins. Trees 1998, 12, 326–333. [Google Scholar] [CrossRef]
- Einhorn, K.S.; Rosenqvist, E.; Leverenz, J.W. Photoinhibition in seedlings of Fraxinus and Fagus under natural light conditions: Implications for forest regeneration? Oecologia 2004, 2, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Dusan, R.; Stjepan, M.; Igor, A.; Jurij, D. Gap regeneration patterns in relationship to light heterogeneity in two old-growth beech–fir forest reserves in South East Europe. For. Int. J. For. Res. 2007, 80, 431–443. [Google Scholar] [CrossRef]
- Čater, M.; Levanič, T. Response of Fagus sylvatica L. and Abies alba Mill. in different silvicultural systems of the high Dinaric karst. For. Ecol. Manag. 2013, 289, 278–288. [Google Scholar] [CrossRef]
- Splechtna, B.E.; Gratzer, G. Natural disturbances in Central European forests: Approaches and preliminary results from Rothwald, Austria. For. Snow Landsc. Res. 2005, 79, 57–67. [Google Scholar]
- Weiner, J.; Stoll, P.; Muller-Landau, H.; Jasentuliyana, A. The Effects of Density, Spatial Pattern, and Competitive Symmetry on Size Variation in Simulated Plant Populations. Am. Nat. 2001, 158, 438–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vospernik, S.; Sterba, H. Do competition-density rule and self-thinning rule agree? Ann. For. Sci. 2015, 72, 379–390. [Google Scholar] [CrossRef]
- Wang, X.; Wiegand, T.; Wolf, A.; Howe, R.; Davies, S.J.; Hao, Z. Spatial patterns of tree species richness in two temperate forests. J. Ecol. 2011, 6, 1382–1393. [Google Scholar] [CrossRef]
- Gray, A.N.; Spies, T.A. Gap Size, Within-Gap Position and Canopy Structure Effects on Conifer Seedling Establishment. J. Ecol. 1996, 84, 635–645. [Google Scholar] [CrossRef]
- Stancioiu, P.T.; O’Hara, K.L. Regeneration growth in different light environments of mixed species, multiaged, mountainous forests of Romania. Eur. J. For. Res. 2006, 125, 151–162. [Google Scholar] [CrossRef]
- Clark, C.J.; Poulsen, J.R.; Levey, D.J.; Osenberg, C.W. Are Plant Populations Seed Limited? A Critique and Meta-Analysis of Seed Addition Experiments. Am. Nat. 2007, 170, 128–142. [Google Scholar] [CrossRef] [PubMed]
- Paluch, J.G. The spatial pattern of a natural European beech (Fagus sylvatica L.)–silver fir (Abies alba Mill.) forest: A patch-mosaic perspective. For. Ecol. Manag. 2007, 253, 161–170. [Google Scholar] [CrossRef]
- Dobrowolska, D.; Bončina, A.; Klumpp, R. Ecology and silviculture of silver fir (Abies alba Mill.): A review. J. For. Res. 2017, 22, 326–335. [Google Scholar] [CrossRef]
- Nopp-Mayr, U.; Kempter, I.; Muralt, G.; Gratzer, G. Seed survival on experimental dishes in a central European old-growth mixed-species forest—Effects of predator guilds, tree masting and small mammal population dynamics. Oikos 2012, 3, 337–346. [Google Scholar] [CrossRef]
- Szwagrzyk, J.; Gratzer, G.; Stępniewska, H.; Szewczyk, J.; Veselinovic, B. High Reproductive Effort and Low Recruitment Rates of European Beech: Is There a Limit for the Superior Competitor? Pol. J. Ecol. 2015, 63, 198–212. [Google Scholar] [CrossRef]
- Jankowiak, R.; Stępniewska, H.; Szwagrzyk, J.; Bilański, P.; Gratzer, G. Characterization of Cylindrocarpon-like species associated with litter in the old-growth beech forests of Central Europe. For. Pathol. 2016, 6. [Google Scholar] [CrossRef]
- Bagchi, R.; Gallery, R.E.; Gripenberg, S.; Gurr, S.J.; Narayan, L.; Addis, C.E.; Freckleton, R.P.; Lewis, O.T. Pathogens and insect herbivores drive rainforest plant diversity and composition. Nature 2014, 506, 85–88. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Density (Individuals ≤ 50 cm Height) m−2 | Density of Seedlings in Micro-Morphology Classes (Individuals ≤ 50 cm Height) m−2 Cover of Micro-Morphology Class | Density of Seedlings on Micro-Site Classes (Individuals ≤ 50 cm Height) m−2 Cover of Micro-Site Class | ||||
|---|---|---|---|---|---|---|
| Species | Equal and convex | Concave | Nurse logs | Litter and lignomoder | Bryophyte covered boulders | |
| Fagus sylvatica | 1.32 | 1.37 | 0 | 0.5 | 1.11 | 0.11 |
| Abies alba | 1.05 | 1.40 | 0 | 0.58 | 1.05 | 0.14 |
| Picea abies | 0.58 | 0.36 | 0 | 0.81 | 0.37 | 0.10 |
| Species | STR | MDD | D ζ | n | r2 |
|---|---|---|---|---|---|
| Fagus sylvatica | 26692 | 6.1 | 169.774 | 199.35 | 0.43 |
| Abies alba | 11712 | 22.6 | 3.481 | 2660.05 | 0.26 |
| Picea abies | 52067 | 38.7 | 0.694 | 7789.65 | 0.20 |
| Species | Picea abies | Abies alba | Fagus sylvatica |
|---|---|---|---|
| Intercept | −1.29 (−10.75; 8.16) | 7.46 ** (3.50; 11.43) | 0.95 * (−0.05; 5.95) |
| Nurse logs | 1.06 ** (0.38; 1.75) | −1.00 ** (−1.61; −0.38) | −0.92 * (−1.80; 0.03) |
| Bryophyte covered boulders | −0.59 (−1.47; 0.28) | −0.76 ** (−1.34; −0.19) | −1.46 ** (−2.49; −0.44) |
| Light levels | −0.08 (−0.26; 0.09) | −0.07 (−0.18; 0.04) | −0.20 ** (−0.35; −0.04) |
| Seed density | 0.01 (−0.04; 0.06) | −0.10 ** (−0.15; −0.04) | 0.002 (−0.002; 0.007) |
| DSL | 0.40 ** (0.14; 0.66) | 0.17 (−0.02; 0.36) | 0.09 (−0.23; 0.41) |
| Nurse log: DSL | −0.47 * (−0.84; −0.10) | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gratzer, G.; Waagepetersen, R.P. Seed Dispersal, Microsites or Competition—What Drives Gap Regeneration in an Old-Growth Forest? An Application of Spatial Point Process Modelling. Forests 2018, 9, 230. https://doi.org/10.3390/f9050230
Gratzer G, Waagepetersen RP. Seed Dispersal, Microsites or Competition—What Drives Gap Regeneration in an Old-Growth Forest? An Application of Spatial Point Process Modelling. Forests. 2018; 9(5):230. https://doi.org/10.3390/f9050230
Chicago/Turabian StyleGratzer, Georg, and Rasmus Plenge Waagepetersen. 2018. "Seed Dispersal, Microsites or Competition—What Drives Gap Regeneration in an Old-Growth Forest? An Application of Spatial Point Process Modelling" Forests 9, no. 5: 230. https://doi.org/10.3390/f9050230
APA StyleGratzer, G., & Waagepetersen, R. P. (2018). Seed Dispersal, Microsites or Competition—What Drives Gap Regeneration in an Old-Growth Forest? An Application of Spatial Point Process Modelling. Forests, 9(5), 230. https://doi.org/10.3390/f9050230