Carbon Pools in a Hemiboreal Over-Mature Norway Spruce Stands



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling and Measurements
2.3. Data Analysis
3. Results
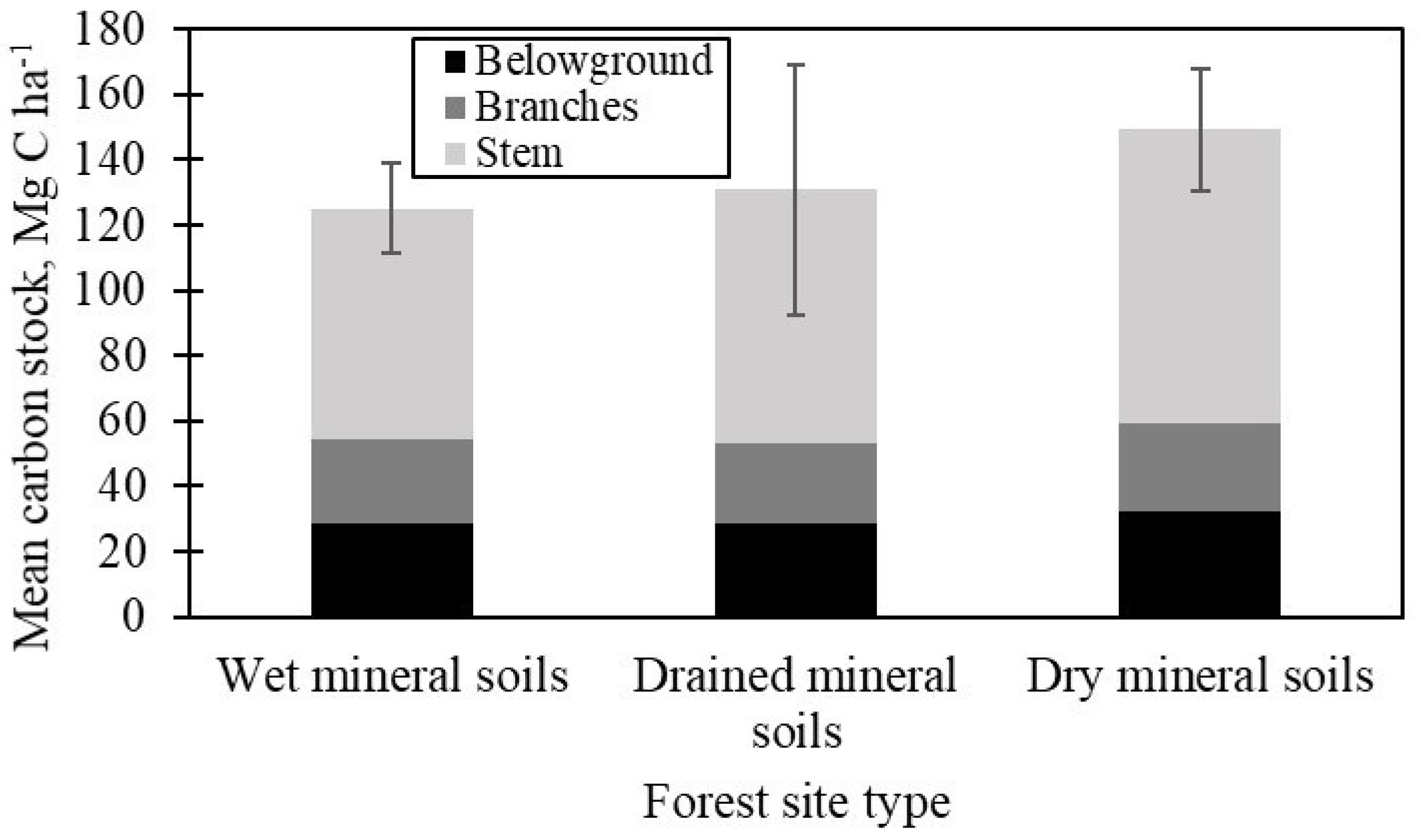
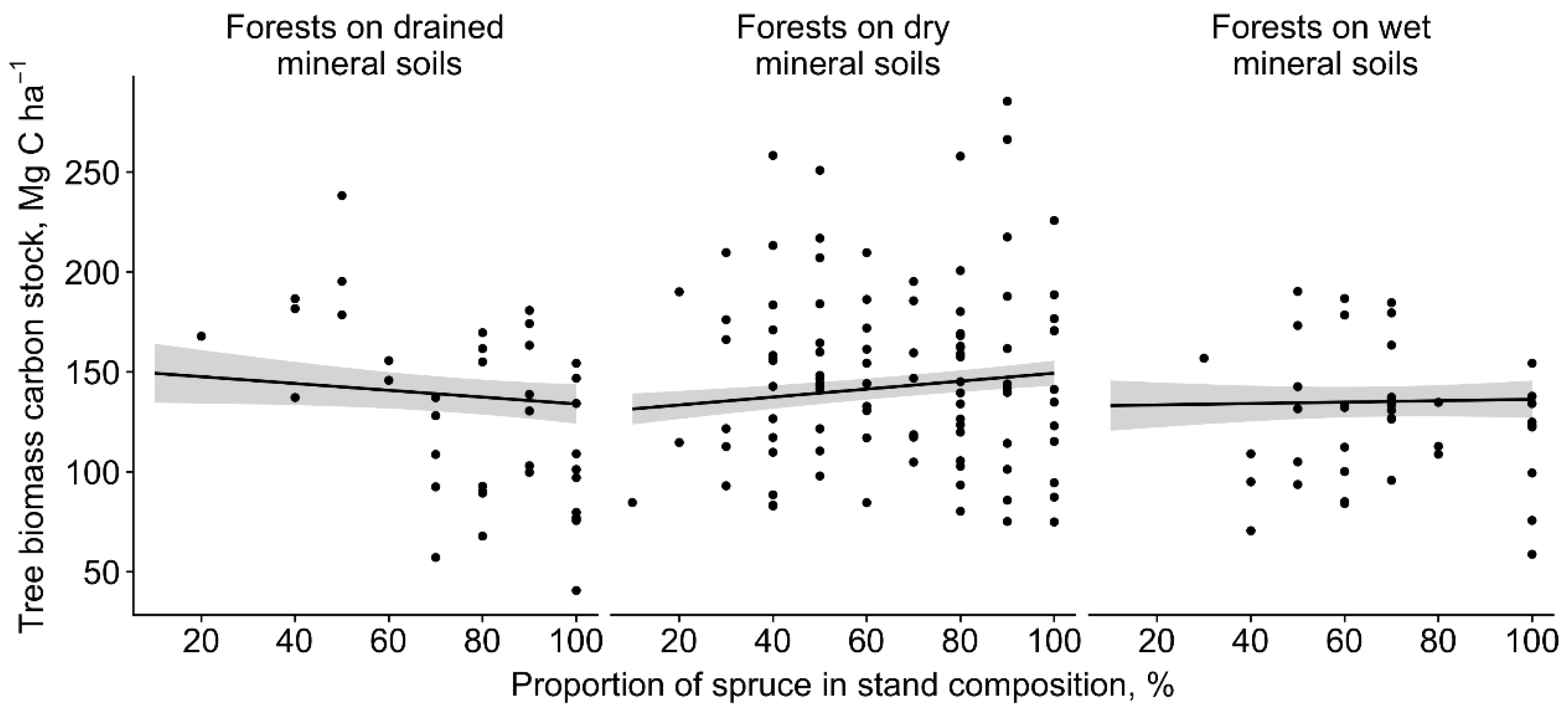
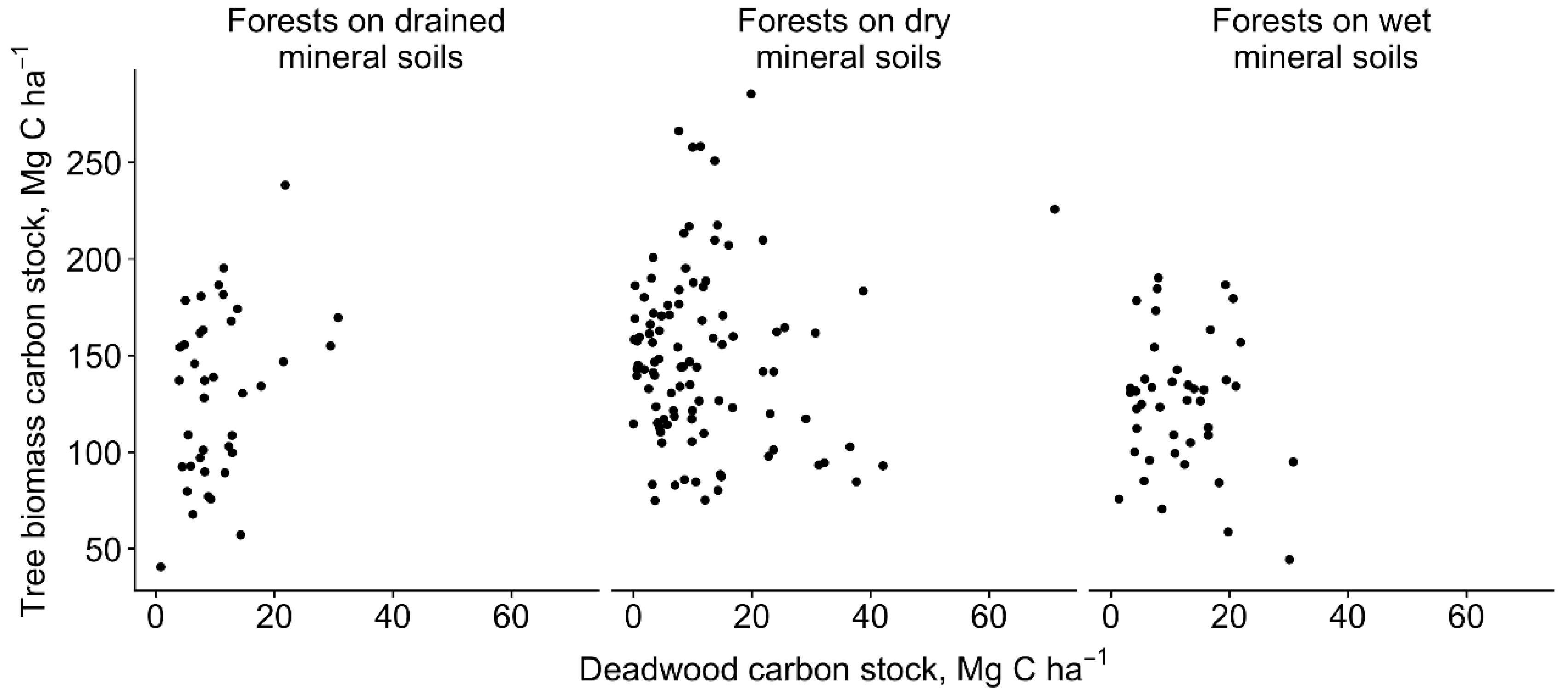
3.1. Tree Biomass Carbon Stock
3.2. Deadwood Carbon Stock
3.3. Soil Carbon
3.4. Total Carbon Stock in Over-Mature Spruce Forests
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Krug, J.H.A. Accounting of GHG emissions and removals from forest management: A long road from Kyoto to Paris. Carbon Balance Manag. 2018, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Finér, L.; Mannerkoski, H.; Piirainen, S.; Starr, M. Carbon and nitrogen pools in an old-growth, Norway spruce mixed forest in eastern Finland and changes associated with clear-cutting. For. Ecol. Manag. 2003, 174, 51–63. [Google Scholar] [CrossRef]
- Bradshaw, C.J.A.; Warkentin, I.G. Global estimates of boreal forest carbon stocks and flux. Glob. Planet. Chang. 2015, 128, 24–30. [Google Scholar] [CrossRef]
- Uri, V.; Kukumägi, M.; Aosaar, J.; Varik, M.; Becker, H.; Morozov, G.; Karoles, K. Ecosystems carbon budgets of differently aged downy birch stands growing on well-drained peatlands. For. Ecol. Manag. 2017, 399, 82–93. [Google Scholar] [CrossRef]
- Neumann, M.; Moreno, A.; Mues, V.; Härkönen, S.; Mura, M.; Bouriaud, O.; Lang, M.; Achten, W.M.J.; Thivolle-Cazat, A.; Bronisz, K.; et al. Comparison of carbon estimation methods for European forests. For. Ecol. Manag. 2016, 361, 397–420. [Google Scholar] [CrossRef]
- Keith, H.; Lindenmayer, D.; MacKey, B.; Blair, D.; Carter, L.; McBurney, L.; Okada, S.; Konishi-Nagano, T. Managing temperate forests for carbon storage: Impacts of logging versus forest protection on carbon stocks. Ecosphere 2014, 5. [Google Scholar] [CrossRef]
- Lundmark, T.; Bergh, J.; Nordin, A.; Fahlvik, N.; Poudel, B.C. Comparison of carbon balances between continuous-cover and clear-cut forestry in Sweden. Ambio 2016, 45, 203–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohjanmies, T.; Triviño, M.; Le Tortorec, E.; Mazziotta, A.; Snäll, T.; Mönkkönen, M. Impacts of forestry on boreal forests: An ecosystem services perspective. Ambio 2017, 46, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Triviño, M.; Pohjanmies, T.; Mazziotta, A.; Juutinen, A.; Podkopaev, D.; Le Tortorec, E.; Mönkkönen, M. Optimizing management to enhance multifunctionality in a boreal forest landscape. J. Appl. Ecol. 2017, 54, 61–70. [Google Scholar] [CrossRef]
- Grassi, G.; House, J.; Dentener, F.; Federici, S.; Den Elzen, M.; Penman, J. The key role of forests in meeting climate targets requires science for credible mitigation. Nat. Clim. Chang. 2017, 7, 220–226. [Google Scholar] [CrossRef]
- Sabatini, S.; Burrascano, W.S.; Keeton, C.; Levers, M.; Lindner, F.; Pötzschner, P.J.; Verkerk, J.; Bauhus, E.; Buchwald, O.; Chaskovsky, N.; et al. Where are Europe’s last primary forests? Divers. Distrib. 2018, 1–14. [Google Scholar] [CrossRef]
- Babst, F.; Bouriaud, O.; Papale, D.; Gielen, B.; Janssens, I.A.; Nikinmaa, E.; Ibrom, A.; Wu, J.; Bernhofer, C.; Köstner, B.; et al. Above-ground woody carbon sequestration measured from tree rings is coherent with net ecosystem productivity at five eddy-covariance sites. New Phytol. 2014, 201, 1289–1303. [Google Scholar] [CrossRef] [PubMed]
- Schelhaas, M.J.; Nabuurs, G.J.; Hengeveld, G.; Reyer, C.; Hanewinkel, M.; Zimmermann, N.E.; Cullmann, D. Alternative forest management strategies to account for climate change-induced productivity and species suitability changes in Europe. Reg. Environ. Chang. 2015, 15, 1581–1594. [Google Scholar] [CrossRef] [Green Version]
- Thom, D.; Seidl, R. Natural disturbance impacts on ecosystem services and biodiversity in temperate and boreal forests. Biol. Rev. Camb. Philos. Soc. 2016, 91, 760–781. [Google Scholar] [CrossRef] [PubMed]
- Vesterdal, L.; Clarke, N.; Sigurdsson, B.D.; Gundersen, P. Do tree species influence soil carbon stocks in temperate and boreal forests? For. Ecol. Manag. 2013, 309, 4–18. [Google Scholar] [CrossRef]
- Odum, E. The strategy of ecosystem development. Science 1969, 164, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Seedre, M.; Kopáček, J.; Janda, P.; Bače, R.; Svoboda, M. Carbon pools in a montane old-growth Norway spruce ecosystem in Bohemian Forest: Effects of stand age and elevation. For. Ecol. Manag. 2015, 346, 106–113. [Google Scholar] [CrossRef]
- Carey, E.V.; Sala, A.; Keane, R.; Callaway, R.M. Are old forest underestimated as global carbon sinks? Glob. Chang. Biol. 2001, 7, 339–344. [Google Scholar] [CrossRef]
- Luyssaert, S.; Schulze, E.D.; Börner, A.; Knohl, A.; Hessenmöller, D.; Law, B.E.; Ciais, P.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213–215. [Google Scholar] [CrossRef] [PubMed]
- Pukkala, T. Does management improve the carbon balance of forestry? Forestry 2017, 90, 125–135. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Euskirchen, E.S. Carbon cycling and storage in world forests: Biome patterns related to forest age. Glob. Chang. Biol. 2004, 10, 2052–2077. [Google Scholar] [CrossRef]
- Varik, M.; Aosaar, J.; Ostonen, I.; Lõhmus, K.; Uri, V. Carbon and nitrogen accumulation in belowground tree biomass in a chronosequence of silver birch stands. For. Ecol. Manag. 2013, 302, 62–70. [Google Scholar] [CrossRef]
- Lõhmus, A.; Kraut, A. Stand structure of hemiboreal old-growth forests: Characteristic features, variation among site types, and a comparison with FSC-certified mature stands in Estonia. For. Ecol. Manag. 2010, 260, 155–165. [Google Scholar] [CrossRef]
- Ahti, T.; Hämet-Ahti, L.; Jalas, J. Vegetation zones and their sections in north-western Europe. Ann. Bot. Fenn. 1968, 5, 169–211. [Google Scholar]
- Baders, E.; Senhofa, S.; Purina, L.; Jansons, A. Natural succession of Norway spruce stands in hemiboreal forests: Case study in Slitere national park, Latvia. Balt. For. 2017, 23, 522–528. [Google Scholar]
- Kasparinskis, R.; Nikodemus, O. Influence of environmental factors on the spatial distribution and diversity of forest soil in Latvia. Est. J. Earth Sci. 2012, 61, 48–64. [Google Scholar] [CrossRef]
- Bušs, K. Basis of Forest Classification in SSR of Latvia; LRZTIPI: Rīga, Latvia, 1976; p. 24. (In Latvian) [Google Scholar]
- Sharma, R.P.; Vacek, Z.; Vacek, S. Nonlinear mixed effect height-diameter model for mixed species forests in the central part of the Czech Republic. J. For. Sci. 2016, 62, 470–484. [Google Scholar] [CrossRef] [Green Version]
- Matuzānis, J. Stand Growth and Yield Models; LatZTIZPLI: Rīga, Latvia, 1983; p. 32. (In Latvian) [Google Scholar]
- Liepiņš, J.; Lazdiņš, A.; Liepiņš, K. Equations for estimating above- and belowground biomass of Norway spruce, Scots pine, birch spp. and European aspen in Latvia. Scand. J. For. Res. 2018, 33, 58–70. [Google Scholar] [CrossRef]
- Thomas, S.C.; Martin, A.R. Carbon content of tree tissues: A synthesis. Forests 2012, 3, 332–352. [Google Scholar] [CrossRef]
- Sandström, F.; Petersson, H.; Kruys, N.; Ståhl, G. Biomass conversion factors (density and carbon concentration) by decay classes for dead wood of Pinus sylvestris, Picea abies and Betula spp. in boreal forests of Sweden. For. Ecol. Manag. 2007, 243, 19–27. [Google Scholar] [CrossRef]
- Köster, K.; Metslaid, M.; Engelhart, J.; Köster, E. Dead wood basic density, and the concentration of carbon and nitrogen for main tree species in managed hemiboreal forests. For. Ecol. Manag. 2015, 354, 35–42. [Google Scholar] [CrossRef]
- Russell, M.B.; Fraver, S.; Aakala, T.; Gove, J.H.; Woodall, C.W.; D’Amato, A.W.; Ducey, M.J. Quantifying carbon stores and decomposition in dead wood: A review. For. Ecol. Manag. 2015, 350, 107–128. [Google Scholar] [CrossRef]
- Fleck, S.; Cools, N.; De Vos, B.; Meesenburg, H.; Fischer, R. The Level II aggregated forest soil condition database links soil physicochemical and hydraulic properties with long-term observations of forest condition in Europe. Ann. For. Sci. 2016, 73, 945–957. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org (accessed on 9 March 2018).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Square Means. R Package Version 1.2.1. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 9 March 2018).
- Vucetich, J.A.; Reed, D.A.; Breymeyer, A.; Degórski, M.; Mroz, G.D.; Solon, J.; Roo-Zielinska, E.; Noble, R. Carbon pools and ecosystem properties along a latitudinal gradient in northern Scots pine (Pinus sylvestris) forests. For. Ecol. Manag. 2000, 128, 135–145. [Google Scholar] [CrossRef]
- Siitonen, J.; Martikainen, P.; Punttila, P.; Rauh, J. Coarse woody debris and stand characteristics in mature managed and old-growth boreal mesic forests in southern Finland. For. Ecol. Manag. 2000, 128, 211–225. [Google Scholar] [CrossRef]
- Fredeen, A.L.; Bois, C.H.; Janzen, D.T.; Sanborn, P.T. Comparison of coniferous forest carbon stocks between old-growth and young second-growth forests on two soil types in central British Columbia, Canada. Can. J. For. Res. 2005, 35, 1411–1421. [Google Scholar] [CrossRef]
- Jacob, M.; Bade, C.; Calvete, H.; Dittrich, S.; Leuschner, C.; Hauck, M. Significance of over-mature and decaying trees for carbon stocks in a Central European natural spruce forest. Ecosystems 2013, 16, 336–346. [Google Scholar] [CrossRef]
- Klein, D.; Höllerl, S.; Blaschke, M.; Schulz, C. The contribution of managed and unmanaged forests to climate change mitigation-A model approach at stand level for the main tree species in Bavaria. Forests 2013, 4, 43–69. [Google Scholar] [CrossRef]
- Mund, M.; Kummetz, E.; Hein, M.; Bauer, G.A.; Schulze, E.D. Growth and carbon stocks of a spruce forest chronosequence in central Europe. For. Ecol. Manag. 2002, 171, 275–296. [Google Scholar] [CrossRef]
- Gao, T.; Hedblom, M.; Emilsson, T.; Nielsen, A.B. The role of forest stand structure as biodiversity indicator. For. Ecol. Manag. 2014, 330, 82–93. [Google Scholar] [CrossRef]
- Shanin, V.; Komarov, A.; Mäkipä, R. Tree species composition affects productivity and carbon dynamics of different site types in boreal forests. Eur. J. For. Res. 2014, 133, 273–286. [Google Scholar] [CrossRef]
- Aakala, T.; Kuuluvainen, T.; Wallenius, T.; Kauhanen, H. Contrasting patterns of tree mortality in late-successional Picea-Abies stands in northern Fennoscandia. J. Veg. Sci. 2009, 20, 1016–1026. [Google Scholar] [CrossRef]
- Angelstam, P.; Kuuluvainen, T. Boreal forest disturbance regimes, successional dynamics and landscape structures: A European perspective. Ecol. Bull. 2004, 51, 117–136. [Google Scholar]
- Seidl, R.; Rammer, W. Climate change amplifies the interactions between wind and bark beetle disturbances in forest landscapes. Landsc. Ecol. 2017, 32, 1485–1498. [Google Scholar] [CrossRef] [PubMed]
- Clarke, N.; Gundersen, P.; Jönsson-belyazid, U.; Kjønaas, O.J.; Persson, T.; Sigurdsson, B.D.; Stupak, I.; Vesterdal, L. Influence of different tree-harvesting intensities on forest soil carbon stocks in boreal and northern temperate forest ecosystems. For. Ecol. Manag. 2015, 351, 9–19. [Google Scholar] [CrossRef]
- Jain, T.B.; Graham, R.T.; Adams, D. Carbon concentrations and carbon pool distributions in dry, moist, and cold mid-aged forests of the rocky mountains. In Integrated Management of Carbon Sequestration and Biomass Utilization Opportunities in a Changing Climate, Proceedings of the 2009 National Silviculture Workshop, Boise, ID, USA, 15–18 June 2009; Jain, T.B., Graham, R.T., Sandquist, J., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2010; pp. 39–59. [Google Scholar]
- Hansson, K.; Olsson, B.A.; Olsson, M.; Johansson, U.; Berggren, D. Differences in soil properties in adjacent stands of Scots pine, Norway spruce and silver birch in SW Sweden. For. Ecol. Manag. 2011, 262, 522–530. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameters ± CI 1 | Dry Mineral Soil | Wet Mineral Soil | Drained Mineral Soil |
|---|---|---|---|
| Mean tree dbh 2, cm | 41.2 ± 4.2 | 32.9 ± 4.8 | 31.1 ± 5.5 |
| Mean tree height, m | 29.7 ± 1.8 | 25.9 ± 2.3 | 26.7 ± 3.3 |
| Mean stand basal area, m2 ha−1 | 37.9 ± 3.4 | 32.7 ± 3.8 | 33.4 ± 8.9 |
| Mean basal area in upper tree layer, m2 ha−1 | 29.9 ± 3.6 | 25.6 ± 3.2 | 26.4 ± 7.2 |
| Mean stand volume, m3 ha−1 | 462.0 ± 66.5 | 357.4 ± 51.4 | 389.4 ± 121.4 |
| Site index, m | 27 | 23 | 25 |
| Mean stand age, years | 181.1 ± 6.2 | 179 ± 11.5 | 185 ± 23.4 |
| Number of sample plots | 98 | 42 | 38 |
| Carbon Pools ± CI 1 | Dry Mineral Soil | Wet Mineral Soil | Drained Mineral Soil |
|---|---|---|---|
| Aboveground tree biomass | 116.8 ± 15.4 | 96.8 ± 11.5 | 102.4 ± 30.3 |
| Belowground tree biomass | 32.3 ± 3.6 | 28.4 ± 2.6 | 28.5 ± 7.9 |
| Total tree biomass carbon stock | 149.2 ± 18.9 | 125.2 ± 14.0 | 130.9 ± 38.1 |
| Lying deadwood | 37.9 ± 3.4 | 32.7 ± 3.8 | 33.4 ± 8.9 |
| Standing snags | 29.9 ± 3.6 | 25.6 ± 3.2 | 26.4 ± 7.2 |
| Total deadwood carbon stock | 11.8 ± 2.3 | 11.8 ± 2.2 | 10.6 ± 2.2 |
| Soil carbon stock | 88.4 ± 23.8 | 88.0 ± 12.4 | 85.6 ± 26.1 * |
| Total carbon stock | 249.3 ± 31.8 | 225.1 ± 11.4 | 227.0 ± 39.8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ķēniņa, L.; Elferts, D.; Bāders, E.; Jansons, Ā. Carbon Pools in a Hemiboreal Over-Mature Norway Spruce Stands. Forests 2018, 9, 435. https://doi.org/10.3390/f9070435
Ķēniņa L, Elferts D, Bāders E, Jansons Ā. Carbon Pools in a Hemiboreal Over-Mature Norway Spruce Stands. Forests. 2018; 9(7):435. https://doi.org/10.3390/f9070435
Chicago/Turabian StyleĶēniņa, Laura, Didzis Elferts, Endijs Bāders, and Āris Jansons. 2018. "Carbon Pools in a Hemiboreal Over-Mature Norway Spruce Stands" Forests 9, no. 7: 435. https://doi.org/10.3390/f9070435
APA StyleĶēniņa, L., Elferts, D., Bāders, E., & Jansons, Ā. (2018). Carbon Pools in a Hemiboreal Over-Mature Norway Spruce Stands. Forests, 9(7), 435. https://doi.org/10.3390/f9070435