Species Identification of Pinus Pollen Found in Belukha Glacier, Russian Altai Mountains, Using a Whole-Genome Amplification Method

Abstract
:1. Introduction
2. Materials and Methods
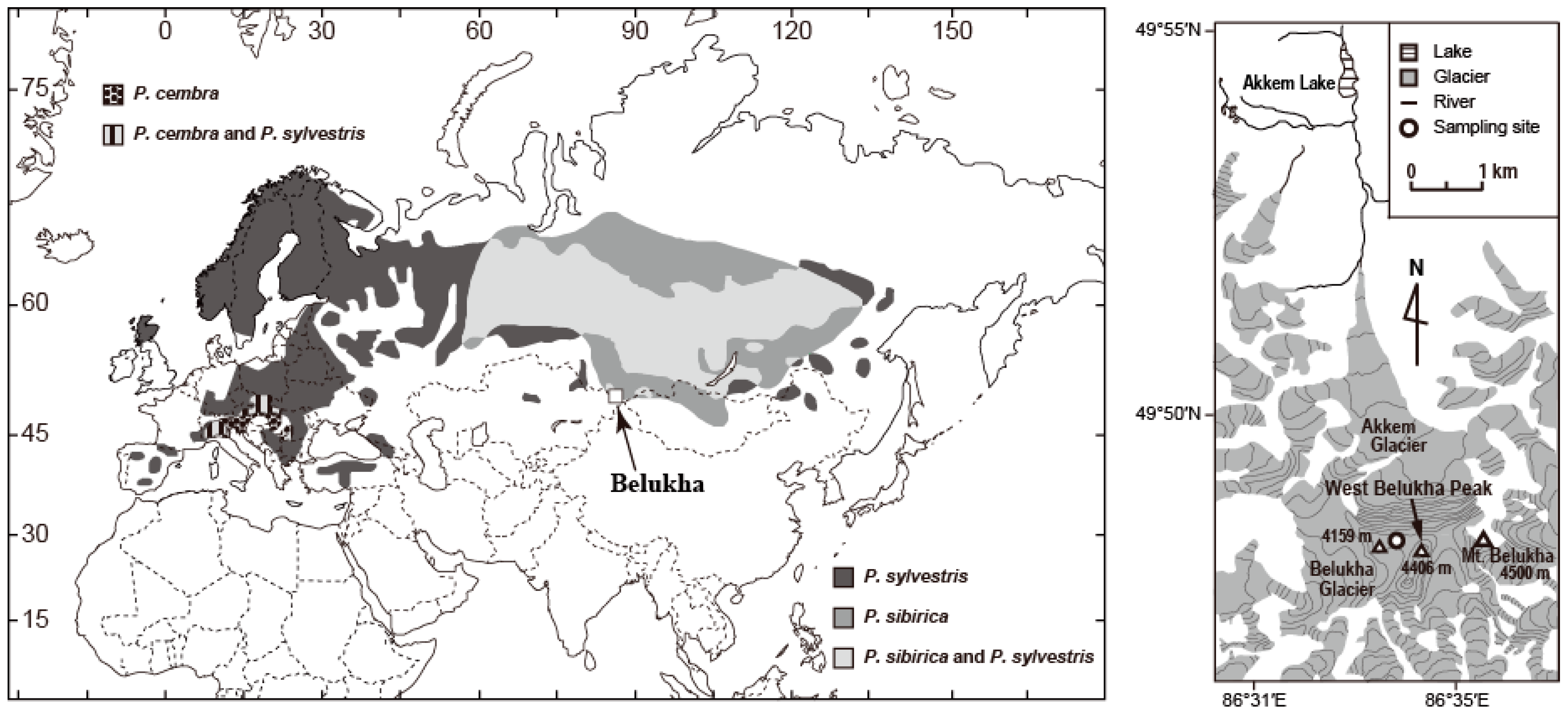
2.1. Study Area and Pollen Samples
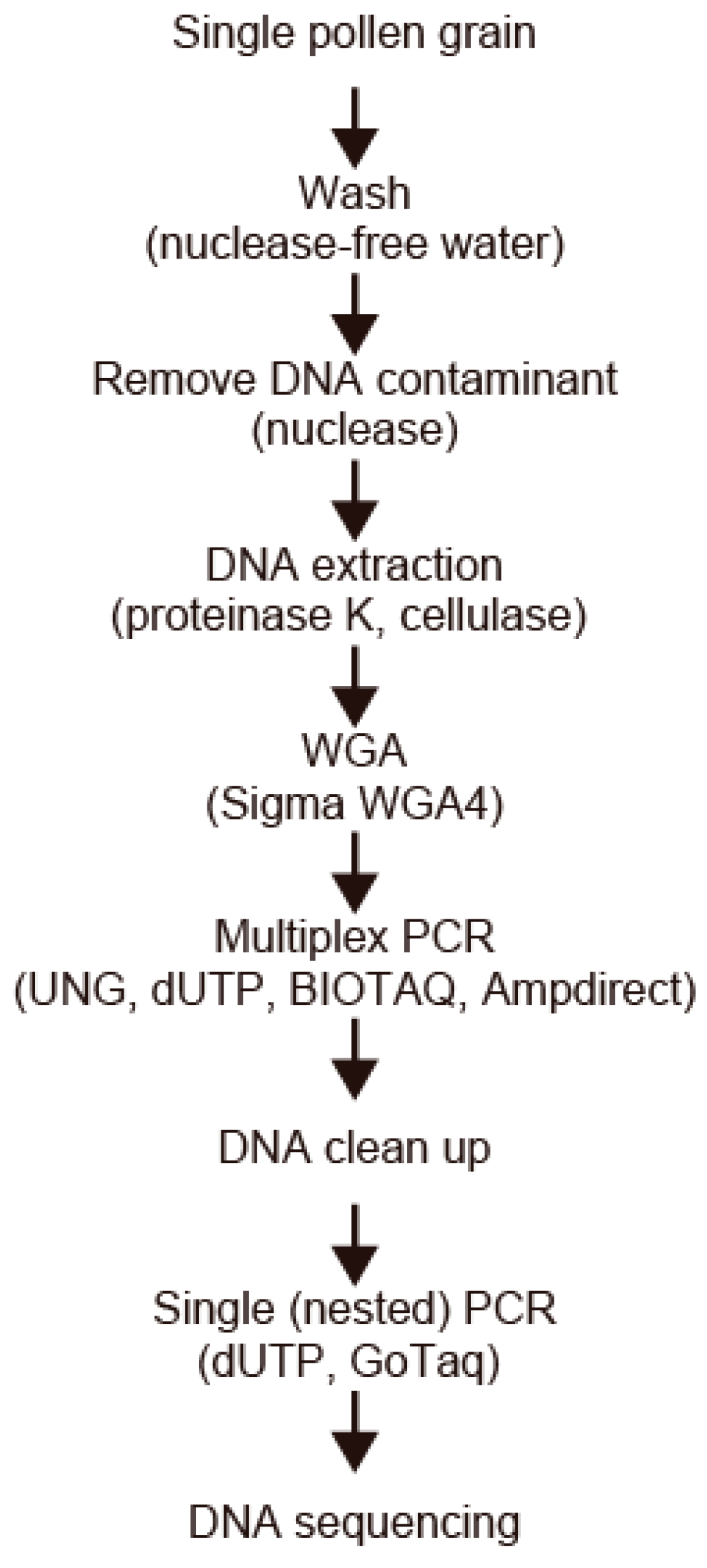
2.2. DNA Extraction from a Single Pollen Grain
2.3. Whole-Genome Amplification
2.4. UNG Treatment and Multiplex PCR Amplification
2.5. Nested PCR
2.6. Identification of Individual Pinus Pollen Grains
3. Results and Discussion
3.1. Pinus Pollen Identification at the Section or Subsection Level
3.2. Species Identification of Pinus Pollen Grains
3.3. Potential Use of Pollen Grains as a Tracer for Emission Sources
3.4. Improvement in the Amplification Success Rate
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Richardson, D.M.; Rundel, P.W. Ecology and biogeography of Pinus: An introduction. In Ecology and Biogeography of Pinus; Richardson, D.M., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 3–46. ISBN 0521789109. [Google Scholar]
- Fredskild, B.; Wagner, P. Pollen and fragments of plant tissue in core samples from the Greenland Ice Cap. Boreas 1974, 3, 105–108. [Google Scholar] [CrossRef]
- Short, S.K.; Holdsworth, G. Pollen, oxygen isotope content and seasonality in an ice core from the Penny Ice Cap, Baffin island. Arctic 1985, 38, 214–218. [Google Scholar] [CrossRef]
- Bourgeois, J.C. Seasonal and interannual pollen variability in snow layers of arctic ice caps. Rev. Palaeobot. Palynol. 2000, 108, 17–36. [Google Scholar] [CrossRef]
- Nakazawa, F.; Fujita, K.; Uetake, J.; Kohno, M.; Fujiki, T.; Arkhipov, S.M.; Kameda, T.; Suzuki, K.; Fujii, Y. Application of pollen analysis to dating of ice cores from lower-latitude glaciers. J. Geophys. Res. 2004, 109, F04001. [Google Scholar] [CrossRef]
- Nakazawa, F.; Fujita, K.; Takeuchi, N.; Fujiki, T.; Uetake, J.; Aizen, V.; Nakawo, M. Dating of seasonal snow/firn accumulation layers using pollen analysis. J. Glaciol. 2005, 51, 483–490. [Google Scholar] [CrossRef]
- Hicks, S.; Isaksson, E. Assessing source areas of pollutants from studies of fly ash, charcoal, and pollen from Svalbard snow and ice. J. Geophys. Res. 2006, 111, D02113. [Google Scholar] [CrossRef]
- Nakazawa, F.; Konya, K.; Kadota, T.; Ohata, T. Reconstruction of the depositional environment upstream of Potanin Glacier, Mongolian Altai, from pollen analysis. Environ. Res. Lett. 2012, 7, 035402. [Google Scholar] [CrossRef] [Green Version]
- Festi, D.; Kofler, W.; Bucher, E.; Mair, V.; Gabrielli, P.; Carturan, L.; Oeggl, K. A novel pollen-based method to detect seasonality in ice cores: a case study from the Ortles Glacier (South Tyrol, Italy). J. Glaciol. 2015, 61, 815–824. [Google Scholar] [CrossRef]
- Nakazawa, F.; Uetake, J.; Suyama, Y.; Kaneko, R.; Takeuchi, N.; Fujita, K.; Motoyama, H.; Imura, S.; Kanda, H. DNA analysis for section identification of individual Pinus pollen grains from Belukha glacier, Altai Mountains, Russia. Environ. Res. Lett. 2013, 8, 014032. [Google Scholar] [CrossRef]
- Yazdani, R.; Lindgren, D.; Stewart, S. Gene dispersion within a population of Pinus sylvestris. Scand. J. For. Res. 1989, 4, 295–306. [Google Scholar] [CrossRef]
- Savolainen, O.; Pyhäjarvi, T.; Knürr, T. Gene flow and local adaptation in trees. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 595–619. [Google Scholar] [CrossRef]
- Wolfe, K.H.; Li, W.H.; Sharp, P.M. Rates of nucleotide substitution vary greatly among plant mitochondrial, chloroplast, and nuclear DNAs. Proc. Natl. Acad. Sci. USA 1987, 84, 9054–9058. [Google Scholar] [CrossRef] [PubMed]
- Bell, K.L.; de Vere, N.; Keller, A.; Richardson, R.T.; Gous, A.; Burgess, K.S.; Brosi, B.J. Pollen DNA barcoding: current applications and future. Genome 2016, 59, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Xu, C.; Li, C.; Sun, J.; Zuo, Y.; Shi, S.; Cheng, T.; Guo, J.; Zhou, S. ycf1, the most promising plastid DNA barcode of land plants. Sci. Rep. 2015, 5, 8348. [Google Scholar] [CrossRef] [PubMed]
- Nachtigall, P.G.; Rodrigues-Filho, L.F.S.; Sodré, D.C.A.; Vallinoto, M.; Pinhal, D. A multiplex PCR approach for the molecular identification and conservation of the Critically Endangered daggernose shark. Endanger. Spec. Res. 2017, 32, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Hong, J.; Lim, J.A.; Heu, S.; Roh, E. Improved multiplex PCR primers for rapid identifcation of coagulase-negative staphylococci. Arch. Microbiol. 2018, 200, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Miner, B.E.; Stöger, R.J.; Burden, A.F.; Laird, C.D.; Hansen, R.S. Molecular barcodes detect redundancy and contamination in hairpin-bisulfite PCR. Nucleic Acids Res. 2004, 32, e135. [Google Scholar] [CrossRef] [PubMed]
- McCloskey, M.L.; Stöger, R.; Hansen, R.S.; Laird, C.D. Encoding PCR products with batch-stamps and barcodes. Biochem. Genet. 2007, 45, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, F.; Miyake, T.; Fujita, K.; Takeuchi, N.; Uetake, J.; Fujiki, T.; Aizen, V.B.; Nakawo, M. Establishing the timing of chemical deposition events on Belukha glacier, Altai Mountains, Russia, using pollen analysis. Arctic Antarct. Alp. Res. 2011, 43, 66–72. [Google Scholar] [CrossRef]
- Eichler, A.; Tinner, W.; Brütsch, S.; Olivier, S.; Papina, T.; Schwikowski, M. An ice-core based history of Siberian forest fires since AD 1250. Quat. Sci. Rev. 2011, 30, 1027–1034. [Google Scholar] [CrossRef]
- Farjon, A. Pines: Drawings and Descriptions of the Genus Pinus, 2nd ed.; Brill: Leiden, The Netherlands, 2005; p. 236. ISBN 9004139168. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Gernandt, D.S.; GeadaLópez, G.; Garcia, S.O.; Liston, A. Phylogeny and classification of Pinus. Taxon 2005, 54, 29–42. [Google Scholar] [CrossRef]
- Little, E.L., Jr.; Critchfield, W.B. Subdivisions of the Genus Pinus (Pines); Miscellaneous Publications 1144; U.S. Department of Agriculture: Washington, DC, USA, 1969; p. 51.
- Price, R.A.; Liston, A.; Strauss, S.H. Phylogeny and systematics of Pinus. In Ecology and Biogeography of Pinus; Richardson, D.M., Ed.; Cambridge University Press: Cambridge, UK, 1988; pp. 49–68. ISBN 0521789109. [Google Scholar]
- Pääbo, S. Ancient DNA: Extraction, characterization, molecular cloning, and enzymatic amplification. Proc. Natl. Acad. Sci. USA 1989, 86, 1939–1943. [Google Scholar] [CrossRef] [PubMed]
- Suyama, Y.; Kawamuro, K.; Kinoshita, I.; Yoshimura, K.; Tsumura, Y.; Takahara, H. DNA sequence from a fossil pollen of Abies spp. from Pleistocene peat. Genes Genet. Syst. 1996, 71, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Parducci, L.; Suyama, Y.; Lascoux, M.; Bennett, K.D. Ancient DNA from pollen: a genetic record of plant population history. Mol. Ecol. 2005, 14, 2873–2882. [Google Scholar] [CrossRef] [PubMed]
- Parks, M.; Cronn, R.; Liston, A. Increasing phylogenetic resolution at low taxonomic levels using massively parallel sequencing of chloroplast genomes. BMC Biol. 2009, 7, 84. [Google Scholar] [CrossRef] [PubMed]
- Gugerli, F.; Senn, J.; Anzidei, M.; Madaghiele, A.; Büchler, U.; Sperisen, C.; Vendramin, G.G. Chloroplast microsatellites and mitochondrial nad1 intron 2 sequences indicate congruent phylogenetic relationships among Swiss stone pine (Pinus cembra), Siberian stone pine (Pinus sibirica), and Siberian dwarf pine (Pinus pumila). Mol. Ecol. 2001, 10, 1489–1497. [Google Scholar] [CrossRef] [PubMed]
- Heinze, B.; Holzer, K. A review of research on Pinus cembra in Austria, with special reference to the conservation of genetic resources. In Proceedings of the 5th Symposium for Research in Protected Areas, Mittersill, Austria, 10–12 June 2013; pp. 279–284. [Google Scholar]
- Bourgeois, J.C.; Koerner, R.M.; Alt, B.T. Airborne pollen: a unique mass tracer, its influx to the Canadian High Arctic. Ann. Glaciol. 1985, 7, 109–116. [Google Scholar] [CrossRef]
- Rogers, C.A.; Levetin, E. Evidence of long-distance transport of mountain cedar pollen into Tulsa, Oklahoma. Int. J. Biometeorol. 1998, 42, 65–72. [Google Scholar] [CrossRef]
- Rousseau, D.D.; Duzer, D.; Cambon, G.; Jolly, D.; Poulsen, U.; Ferrier, J.; Schevin, P.; Gros, R. Long distance transport of pollen to Greenland. Geophys. Res. Lett. 2003, 30, 1765. [Google Scholar] [CrossRef]
- Rousseau, D.D.; Schevin, P.; Duzer, D.; Cambon, G.; Ferrier, J.; Jolly, D.; Poulsen, U. New evidence of long distance pollen transport to southern Greenland in late spring. Rev. Palaeobot. Palynol. 2006, 141, 277–286. [Google Scholar] [CrossRef]
- Mohanty, R.P.; Buchheim, M.A.; Anderson, J.; Levetin, E. Molecular analysis confirms the long-distance transport of Juniperus ashei pollen. PLoS ONE 2017, 12, e0173465. [Google Scholar] [CrossRef] [PubMed]
- Bory, A.J.M. A 10,000 km dust highway between the Taklamakan Desert and Greenland. In Past Global Changes Magazine; Merkel, U., Rousseau, D.D., Stuut, J.B., Winckler, G., von Gunten, L., Kiefer, T., Eds.; PAGES International Project Office: Bern, Switzerland, 2014; Volume 22, pp. 72–73. [Google Scholar]
- Nagatsuka, N.; Takeuchi, N.; Nakano, T.; Shin, K.; Kokado, E. Geographical variations in Sr and Nd isotopic ratios of cryoconite on Asian glaciers. Environ. Res. Lett. 2014, 9, 045007. [Google Scholar] [CrossRef] [Green Version]
- Vallelonga, P. The enigma of dust provenance: Where else does Antarctic dust come from? In Past Global Changes Magazine; Merkel, U., Rousseau, D.D., Stuut, J.B., Winckler, G., von Gunten, L., Kiefer, T., Eds.; PAGES International Project Office: Bern, Switzerland, 2014; Volume 22, pp. 74–75. [Google Scholar]
- Matsushita, M. Pollen Analysis and Archaeology; Douseisha: Tokyo, Japan, 2004; p. 135. ISBN 4886213030. (In Japanese) [Google Scholar]
- Willis, K.J.; Bennett, K.D.; Birks, H.J.B. The late Quaternary dynamics of Pines in Europe. In Ecology and Biogeography of Pinus; Richardson, D.M., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 107–121. ISBN 0521789109. [Google Scholar]
- Suyama, Y.; Kawamuro, K.; Takahara, H.; Shichi, K.; Yoshimaru, H.; Kinoshita, I.; Yoshimura, K.; Tsumura, Y. Study on vegetation dynamics and biological evolution from DNA analyses of ancient pollen. Chikyu Mon. Symp. 2003, 42, 187–192. (In Japanese) [Google Scholar]



{kind=link}
{kind=link}
| Mixture for Multiplex PCR | Primer ID | Region | Primers for Multiplex PCR | Sequence (5′-3′) | Product Size (bp) | Primers for Nested PCR | Sequence (5′-3′) | Product Size (bp) |
|---|---|---|---|---|---|---|---|---|
| Mixture 1 (Matsu1-Matsu5) | Matsu1 | clpP | Matsu m1F | CAACTTGGGTCGACTTATACAACC | 185 | Matsu n1F | CGACTTATACAACCGACTTTATCG | 174 |
| Matsu m1R | ACCTACCCCTGGTATTACTGATCC | Matsu n1R | CCTACCCCTGGTATTACTGATCC | |||||
| Matsu2 | rpoA | Matsu m2F | CTGGGTCCAACAATATAAATAGAAGC | 174 | Matsu n2F | CAATATAAATAGAAGCTTCTCGGATTCC | 164 | |
| Matsu m2R | AGTAGAAGGAACATGTATCACACG | Matsu n2R | same as Matsu m2R | |||||
| Matsu3 | atpB | Matsu m3F | GGAGAACCTGTCGATAATTTGGG | 130 | Matsu n3F | CCTGTCGATAATTTGGGTCCTG | 115 | |
| Matsu m3R | GATCTACTACTTTAATGCCTGTTTCG | Matsu n3R | CTTTAATGCCTGTTTCGAAGATGG | |||||
| Matsu4 | rpoC1 | Matsu m4F | GCCTAGTAAATTTTCACGAAATCTTCCC | 134 | Matsu n4F | TTCCCTCTTTGCCTTCGATCAC | 111 | |
| Matsu m4R | AGGAAGCCGTAGATGCACTTC | Matsu n4R | same as Matsu m4R | |||||
| Matsu5 | ycf3 | Matsu m5F | CTGGAGATAGAACAATTCCTTCTGTC | 143 | Matsu n5F | GTATCCCCGGTCAATGCAC | 115 | |
| Matsu m5R | CCTATCAAATAGGTTCAACTATACAAGC | Matsu n5R | same as Matsu m5R | |||||
| Mixture 2 (Matsu6-Matsu10) | Matsu6 | ycf4 | Matsu m6F | GGTTCACATTATCACCAGTACGAG | 160 | Matsu n6F | CACATTATCACCAGTACGAGTTAAAGG | 157 |
| Matsu m6R | CCCGGAATAAATCGTCGTATTTTC | Matsu n6R | same as Matsu m6R | |||||
| Matsu7 | rpoB | Matsu m7F | ACTCCAGAATGGCTTTTTCC | 133 | Matsu n7F | AATGGCTTTTTCCCTCGAC | 109 | |
| Matsu m7R | TCAGSTATGGGTTTAAATCTCG | Matsu n7R | TCTCGAAGAGATTCTGGACAATAC | |||||
| Matsu8 | rpoC1 | Matsu m8F | AGGCATAAGACCATCCATTTTGG | 186 | Matsu n8F | CCATCCATTTTGGTTCTATATTTGTTCGG | 177 | |
| Matsu m8R | ACATATGGAATGGAAGAACTTGGTG | Matsu n8R | CACATATGGAATGGAAGAACTTGGTG | |||||
| Matsu9 | ycf3 | Matsu m9F | GATTAATCCCCGGAGAATACAGG | 147 | Matsu n9F | GGAGAATACAGGGCGTTAAGAAC | 136 | |
| Matsu m9R | ATTAAMAGGGGCTAGTGTATTTCC | Matsu n9R | same as Matsu m9R | |||||
| Matsu10 | ycf4 | Matsu m10F | AGTATTTGCTGAGATGACAATAGGG | 142 | Matsu n10F | same as Matsu m10F | 118 | |
| Matsu m10R | AACCTATAACAGGGTCTCGAAAAAG | Matsu n10R | GAAGTAATTTCTTTTGGGCTTGTATCC | |||||
| Mixture 3 (Strbs1, 3, 5, 7, 9, 11) | Strbs1 | ycf1 | Strbs m1F | TTCGGATCGAGTGAAAGCTC | 146 | Strbs n1F | GAAAGCTCTAAGCCATGGATCTC | 113 |
| Strbs m1R | GGTCTATTGTTCCACGCAATG | Strbs n1R | CCCCATTAAGCAATGGATCATAC | |||||
| Strbs3 | ycf1 | Strbs m3F | CTGAGCATTGGCAGGAATTG | 148 | Strbs n3F | CAGGAATTGGAACACAAAAGC | 125 | |
| Strbs m3R | GCTAATGGATAAARCCGTTTCG | Strbs n3R | AGCCGTTTCGAAATAGGTTC | |||||
| Strbs5 | ycf1 | Strbs m5F | GGAAATGCAGATCCAAGAAATC | 182 | Strbs n5F | same as Strbs m5F | 148 | |
| Strbs m5R | CAACGTTTCTARCATCAATTCG | Strbs n5R | CACTAAAGAGTTTGTGTAGATCCGTTC | |||||
| Strbs7 | ycf1 | Strbs m7F | GGATGATTCAACGCAAACG | 199 | Strbs n7F | CGGATGATTCAACGCAAAC | 180 (186) | |
| Strbs m7R | TTTGACCTTTCTTGTACGAATCC | Strbs n7R | TCCTACTCGTTGATATTTGAATTGG | |||||
| Strbs9 | ycf1 | Strbs m9.1F | CCTAAAGATTATTATGACACGTTCG | 194 | Strbs n9.1F | CGATTCTGGTAGAGTGAATCAGG | 169 | |
| Strbs m9.1R | TTTGATTGAGCCACTAATATGAGAC | Strbs n9.1R | same as Strbs m9.2R | |||||
| Strbs m9.2F | TTATGACACGTTCGATTCTGG | 181 | ||||||
| Strbs m9.2R | TGATTGAGCCACTAATATGAGACC | |||||||
| Strbs11 | ycf1 | Strbs m11F | CGTTTGAAGCCTTGGCATAG | 211 | Strbs n11F | same as Strbs m11F | 177 | |
| Strbs m11R | CTCTCTTCAATCCTTTCTTCAATCC | Strbs n11R | TCCTTTCTTCAATCCTTTCTTCAC | |||||
| Mixture 4 (Strbs2, 4, 6, 8, 10, 12) | Strbs2 | ycf1 | Strbs m2.1F | GGCATTGCGTGGAACAATAG | 178 | Strb n2.1F | same as Strb m2.1F | 129 |
| Strbs m2.1R | CAATTCCTGCCAATGCTCAG | Strb n2.1R | TTCGGAAATCCCTCTTTACAGTC | |||||
| Strbs m2.2F | GGGGATTCTTTTACTGAAATGTATG | 169 | ||||||
| Strbs m2.2R | TCGGAAATCCCTCTTTACAGTC | |||||||
| Strbs4 | ycf1 | Strbs m4F | GTTCCGAGCACTAAATCATCG | 196 | Strb n4F | CGAAAAGAGGAAAAGTTGAACC | 177 | |
| Strbs m4R | GGATCTGCATTTCCAACAAATC | Strb n4R | same as Strb m4R | |||||
| Strbs6 | ycf1 | Strbs m6F | ACCGAATTGATGGTAGAAACG | 167 | Strb n6F | CCGAATTGATGGTAGAAACG | 137 | |
| Strbs m6R | TTGCGTTGAATCATCCGTAG | Strb n6R | CGCGACAATTTCGTAGTATTG | |||||
| Strbs8 | ycf1 | Strbs m8F | CATGCCGAGTCAGATTATCGTC | 178 | Strb n8F | same as Strb m8F | 140 | |
| Strbs m8R | TCACTCTACCAGAATCGAACGTG | Strb n8R | GGATAAGTGGGTATTTCCATTCTTCTC | |||||
| Strbs10 | ycf1 | Strbs m10F | GAGGTCTCATATTAGTGGCTCAATC | 256 | Strb n10F | same as Strb m10F | 228 | |
| Strbs m10R | CAAGGCTTCAAACGAAAAGG | Strb n10R | GCTTTATCTGCATACCATATTTGTACC | |||||
| Strbs12 | ycf1 | Strbs m12F | GGATTGAAGAAAGGATTGAAGAGAG | 174 | Strb n12F | GGATTGAAGAAAGGATTGAAGAG | 135 | |
| Strbs m12R | ACCCATAGGGGTAGTCTCAGTCTC | Strb n12R | TTGTATCCGGTCATTAAGTTCAC | |||||
| Mixture 5 (Pns1-Pns5) | Pns1 | ycf1 | Pns m1F | TTTCGGATCGAGTGAAAGCTC | 150 | |||
| GGTTTATTGTTCCACGGAATGC | ||||||||
| Pns2 | ycf1 | Pns m2F | GGTCAAGTAGAAGATCAACAAACTG | 149 | ||||
| Pns3 | ycf1 | Pns m3F | CCAACCATATCGTTTATCAAGC | 213 | ||||
| TCTCTACGACGTTTTGGAAGC | ||||||||
| Pns4 | rbcL | Pns m4F | TTGTACACAAGCTTCTAGAGCAACC | 190 | ||||
| AGATTGGGTATCTATGCCAGGTG | ||||||||
| Pns5 | rpl20-rps18 | Pns m5F | AGAGGCAGTTGCTTCCAAATC | 161 | ||||
| TCCGGGAGAATCTGTTCTATCC | ||||||||
| Mixture 6 (Pns6-Pns7) | Pns6 | ycf1 | Pns m6F | TTGCTCTTCAGAGGAATGTTCG | 126 | |||
| TATACATCAGGAATTGGTCATCCAC | ||||||||
| Pns7 | ycf1 | Pns m7F | CTCGGCAATAATGAGCCAAAG | 149 | ||||
| GGGACATTATTTGAATGCTACTGC | ||||||||
| Genus | Subgenus | Section | Subsection |
|---|---|---|---|
| Pinus | Strobus | Parrya | Cembroides |
| Nelsoniae | |||
| Balfourianae | |||
| Quinquefoliae | Strobus | ||
| Krempfianae | |||
| Gerardianae | |||
| Pinus | Trifoliae | Australes | |
| Ponderosae | |||
| Contortae | |||
| Pinus | Pinus | ||
| Pinaster |
| Primer ID | Region | Belukha 1 | Belukha 2 | Belukha 3 | Belukha 4 | Belukha 5 |
|---|---|---|---|---|---|---|
| Matsu1 | clpP | Q | - | - | - | - |
| Matsu2 | rpoA | Q | Q | Q | - | - |
| Matsu3 | atpB | Q | Q | Q | Q | P |
| Matsu4 | rpoC1 | S | S | S | - | - |
| Matsu5 | ycf3 | Q | Q | Q | - | - |
| Matsu6 | Ycf4 | St | - | - | - | - |
| Matsu7 | rpoB | - | S | Q | - | - |
| Matsu8 | rpoC1 | St | St | - | - | - |
| Matsu9 | ycf3 | Q | - | Q | - | - |
| Matsu10 | Ycf4 | Q | - | - | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakazawa, F.; Suyama, Y.; Imura, S.; Motoyama, H. Species Identification of Pinus Pollen Found in Belukha Glacier, Russian Altai Mountains, Using a Whole-Genome Amplification Method. Forests 2018, 9, 444. https://doi.org/10.3390/f9080444
Nakazawa F, Suyama Y, Imura S, Motoyama H. Species Identification of Pinus Pollen Found in Belukha Glacier, Russian Altai Mountains, Using a Whole-Genome Amplification Method. Forests. 2018; 9(8):444. https://doi.org/10.3390/f9080444
Chicago/Turabian StyleNakazawa, Fumio, Yoshihisa Suyama, Satoshi Imura, and Hideaki Motoyama. 2018. "Species Identification of Pinus Pollen Found in Belukha Glacier, Russian Altai Mountains, Using a Whole-Genome Amplification Method" Forests 9, no. 8: 444. https://doi.org/10.3390/f9080444
APA StyleNakazawa, F., Suyama, Y., Imura, S., & Motoyama, H. (2018). Species Identification of Pinus Pollen Found in Belukha Glacier, Russian Altai Mountains, Using a Whole-Genome Amplification Method. Forests, 9(8), 444. https://doi.org/10.3390/f9080444