1. Introduction
Genetic engineering is a form of breeding known for its high efficiency and relative speed compared with traditional breeding. Since the first transgenic plants were created in 1983, more than 160 million hectares have been planted with genetically modified crops, and this area is expected to increase in the future [
1]. However, commercial production of transgenic plants is currently limited to a few crops, such as maize, soybean, and cotton [
2]. Commercial cultivation of transgenic trees remains very limited. Except for
Carica papaya L. ringspot virus-resistant papaya and small-scale cultivation of transgenic poplar, transgenic trees are not yet grown commercially [
3,
4].
It has been reported that about one-fifth of the world’s irrigated land is adversely affected by soil salinization and, in China, the total area of available salty land is 3.67 × 10
7 hm
2 [
5]. Genetically engineered plants have potential application to these lands. Using transgenic technologies, genes known to increase resistance to saline soils can be transferred into host plants, and the resultant transgenic plants can be selected for stress tolerance and other properties such as productivity. Compared to traditional breeding methods, transgenic breeding is quicker and thus more cost-effective. Transgenic breeding also enables genes to be moved between species, and even between Kingdoms, something that traditional breeding methods cannot achieve.
Populus is cultivated widely in China as an important tree species for timber and urban greening. It is reported that more than 7 million hectares of
Populus have been afforested in China [
6]. Triploid
Populus tomentosa, an economically important species, is famous for its rapid growth and high productivity. It has been cultivated widely for pulpwood, firewood, and timber [
6]. To improve the biotic and abiotic stress resistance genes, such as
mtlD,
JERF,
DREB, and
NHX, have been transformed into several species of
Populus, including triploid
Populus tomentosa [
7,
8,
9,
10]. However, transgenic plants continue to be controversial and resistance to cultivation remains. One of the major issues is uncertainty about their influences on the environment. According to the Cartagena protocol on Biosafety, the possible adverse impacts of transgenic plants include at least six categories: (1) potential impacts on non-target species; (2) potential spread of transgenic crops as weeds; (3) potential for cross-pollination between transgenic crops and non-transgenic crops and wild plants; (4) potential impacts on soil bacteria and the nitrogen cycle; (5) indirect effects on the environment; and (6) potential effects on human health [
11]. To fulfil the terms of the Cartagena protocol on Biosafety, environmental biosafety assessment must be performed to collect data in support of safety of transgenic plants [
12]. Because tree species are commonly perennial and have longer lifespans, longer biodegradation, greater potential for long-distance dispersal of pollen and seeds, and greater size than crops and other annual plants, the potential impact of transgenic trees on the environment may be more substantial and continuous [
13]. To date, many biosafety studies and field supervisory tests for genetically modified plants have been conducted [
1,
14,
15].
To evaluate the potential risks of transgenic plants on soil bacteria and other undergrowth vegetation, the number of the rhizosphere microbe community and allelopathic activity of transgenic plants must be measured because foreign gene expression could change the released substances of the receptor plants, which may impact the rhizosphere microbe community or the growth of undergrowth vegetation [
16]. Although some field trials of transgenic plants have shown no significant difference on the microbial communities or allelopathic activity between transgenic and non-transgenic plants [
1,
16,
17,
18], Fang et al. [
19] found inhibiting the expression of a Phenylalanine Ammonia-lyase by RNAi in rice changed the allelopathy and rhizosphere microflora. Beckers et al. [
20] thought the rhizosphere microflora changes might be due to changes of metabolism in transgenic plants. These studies indicated that overexpress or suppress the expression of some genes may change the allelopathy or rhizosphere microflora of transgenic plants, which may depend on the variety of foreign gene and receptor plants. Thus, the allelopathy or rhizosphere microflora of all the transgenic plants must be measured during the field test.
Another major constraint on the use of genetically engineered plants is gene transfer, including horizontal gene transfer (HGT) and vertical gene transfer. HGT refers to the transfer of genes between sexually incompatible organisms in a manner other than traditional reproduction. Although the frequency of HGT from plants to bacteria is quite low [
21], the transformation of
Acinetobacter sp. BD413 with the neomycin phosphotransferase II (
npt II) gene from transgenic plants has been reported [
22]. DNA could be released into soil from decaying organs, such as fallen leaves, and if such DNA was not rapidly degraded by extracellular DNAses, it might be taken up by competent bacteria [
23,
24]. The latter process involves the transfer of foreign genes into offspring through sexual reproduction and known as gene flow. Gene flow is a primary determinant of potential ecological impacts of transgenic trees [
3], and it has been reported in other plants [
25,
26]. Some experts thought gene flow may lead to the loss of genetic diversity in the germplasm pool, and the loss of valued species, ecosystem resources, or crop yield and quality due to weedless [
27]. Thus, how long the foreign DNA could exist in decomposing leaves and the potential of gene flow must be considered in an environmental biosafety assessment of transgenic plants.
Here, we report an assessment of the environmental impact of transgenic triploid
P. tomentosa harboring an
AhDREB1 gene, which encodes a dehydration responsive element-binding (DREB)-like transcription factor cloned from the halophyte
Atriplex hortensis [
28]. The expression of this transgene improves the salt tolerance of receptor trees significantly by regulating multiple salinity and alkali stress resistance-related genes. Such trees have been cultivated in the field since 2005 [
15]. Based on the experience and knowledge obtained from a previous trial, we assessed the effects on soil microorganisms, pollen fertility, persistence of DNA in decomposing leaves, and allelopathic activity. We intended to provide a reference for large-scale field release and for deployment of genetic engineering tree breeding programs in the future.
2. Materials and Methods
2.1. Plant Materials
The transgenic
Populus line T-46, containing the
DREB gene derived from
A.
hortensis and driven by the cauliflower mosaic virus 35S promoter, was used as the subject transgenic tree [
9]. The non-transgenic
Populus line 401, which is a triploid hybrid
Populus “401” ((
P.
tomentosa ×
P.
bolleana) ×
P.
tomentosa) created by Beijing Forestry University, was used for comparison (control). It was also the receptor plant of transgenic
Populus T-46. The foreign gene did not affect the growth of host plant and improved the saline resistance in the greenhouse condition. The triploid hybrid
Populus “401” was obtained by hybridizing diploid
P.
tomentosa (♂) and “
P.
tomentosa ×
P.
bolleana” (♀), and, to get the 2n female gamete, the flower bud of
P.
tomentosa ×
P.
bolleana was treated with physical method before pollination [
29,
30].
2.2. Field Trial Conditions
The field trial started in 2005 and the field conditions were presented previously [
15]. The experimental field was located in Dongying, Shan Dong Province, China (36°56′ N, 118°17′ E), and the annual average temperature, rainfall, amount of sunshine, and evaporation were 12.3 °C, 597 mm, 2590.1 h, and 2032 mm, respectively. This study was approved by the Chinese State Forestry Administration (Permit (2005) No. 07). In China, a field test of GMO requires and application to Chinese State Forestry Administration, and the transgenic plants can only be released on the specified location after being approved. All of the poplar types were cultivated in the field and were grown in saline soil that was irrigated (only once at planting). The field was managed as for standard
Populus. The trial field, which covered an area of 37,000 m
2, contained transgenic poplars and control poplars distributed in rows. An external border planted with
Fraxinus chinensis was used as a buffer zone to prevent transgene flow through pollen dispersal. To investigate whether the transgenic trees could influence other plants in the field, weeds and other wild plants in the field were not removed. To reduce anthropogenic influences, the field was located far from residential areas. We originally had five fields containing five transgenic lines in total; unfortunately, the other fields were destroyed in 2008 for local construction requirement and only this field was preserved.
2.3. Materials Collection
Soil and leaf samples were collected every season from July 2014 to May 2015, and stems were collected from 10 randomly selected transgenic and 10 randomly selected non-transgenic poplars. All of the leaves were taken near the tip of the lowest branches (in winter, because there are no leaves on the branch, we collected the leaves that were just about to fall from the branches as the leaves of winter). Soil samples were collected ~30 cm from the trunk (the transgenic tree was the center of an equilateral triangle, and the sampling points were at the three angles; the soil samples were obtained as close to the roots as possible and at the same distance from the trunk, and then the three samples were fully mixed together). The floral branches of transgenic and non-transgenic plants were collected in January 2015. In addition, female (as hybrid materials) and male (as control) floral branches were collected from P. tomentosa × P. bolleana “TB04” and “TB16” at the West Campus of China Agricultural University. All branches were water-cultured in a greenhouse (15–25 °C) at Beijing Forestry University. Pollen samples were collected from freshly dehiscent anthers on transgenic and non-transgenic plants.
2.4. Determining Allelopathic Activity in Leaves from Transgenic Populus
Allelopathic activity was determined by the sandwich method and the soil-mix method according to Yu et al. [
16] with a few modifications. Allelopathic activity determined by the sandwich method was accomplished by harvesting leaves from 10 individual trees of transgenic trees and non-transgenic trees. The fresh leaf material was dried at 60 °C for 24 h. The dried leaves were stored desiccated and used for assays within two weeks. Dried leaf tissue (100 mg) was placed into sterilized Petri dishes (60 mm in diameter and 16 mm in depth). Low melting point agar (5 mL, 0.5%
w/
v) was added to each dish; the agar was allowed to cool and solidify and then another 5 mL of low-melting point agar was added. Five lettuce seeds were placed in each dish, and the plates were incubated in the dark for 72 h at 25 °C (six dishes, each individual tree). For the soil-mix method, 1 g of dried leaf tissue was mixed with 100 g of soil and then divided equally into six plastic cells (3.5 cm in length, 3.5 cm in width, and 2.5 cm in depth). Five lettuce seeds were then sown on the surface of the leaf-soil mixture. Water was applied to each cell, and the cells were placed into a plant growth chamber and incubated in the dark for five days at 25 °C. Radicle and hypocotyl lengths were measured after the growth of the seedlings had been terminated by an overnight freezing at −20 °C. The lowest and highest values were discarded, and means were calculated from the values for the three remaining seedlings from all six dishes for each individual poplar, totally 18 seedlings. Allelopathic activity was evaluated as relative growth compared to growth of control seedlings that were grown in agar or soil without
Populus leaves.
Differences in allelopathic activity between trangenic and non-transgenic poplar were assessed by a one-way ANOVA using SPSS 19.0 software (SPSS Inc., Chicago, IL, USA).
2.5. Evaluation of Soil Microbe Populations
To assess the impact of the transgenic trees on soil microbe numbers, soil samples were collected four times (from July 2014 to May 2015) and evaluated for numbers of bacteria, actinomycetes, and fungi using the spread plate method according to Wei et al. [
31]. One half (10 g) of the weighed 20-g soil samples was dried overnight at 80 °C. The remaining 10 g of soil were diluted 10-fold with sterile water and then placed on a shaking incubator (150 rpm) for 30 min. The soil samples were then withdrawn in 1 mL of diluent into a transfer pipette and used to coat medium plates (beef extract-peptone medium for bacteria, Martin agar medium for actinomycetes, and Gause’s No. 1 synthetic medium base, which was modified for fungi). After 1 h at room temperature, the plates were transferred to an incubator at 27 °C for bacteria, 28 °C for actinomycetes, and 25 °C for fungi. Fungal colonies were counted three days after inoculation. Actinomycetes and bacteria were counted seven days after inoculation. To distinguish colonies of actinomycetes and bacteria, all colonies were counted and then the agar surface was gently wiped with 70% ethanol. The remaining colonies were counted as actinomycetes, and the other colonies were counted as bacteria.
Differences in soil microbe populations between transgenic and non-transgenic poplar were assessed by a one-way ANOVA using SPSS 19.0 software (SPSS Inc., Chicago, IL, USA).
2.6. Foreign Gene DNA Persistence in Decomposing Leaves
The experiment was started in July 2014 and completed in December 2014. Air-dried transgenic leaves (10 g) were placed into plastic boxes (50 cm in height, 25 cm in width, and 15 cm in depth), which contained about 3 kg of cultivated soil. The leaves were placed above the soil. The boxes were covered by permeable fiberglass nets to keep the plant material in the boxes as intact as possible. To keep the leaves under outdoor conditions, the boxes were placed on the balcony of our laboratory at Beijing Forestry University.
Samples were collected once each month. Genomic DNA was isolated from the leaves of the transgenic samples using a Plant Genomic DNA Kit (Tiangen Biotech, Beijing, China); polymerase chain reaction (PCR) amplifications were performed to identify the
AhDREB1 gene coding sequence; and three combinations of primers were used at different positions. The characteristics of the primers used in this study are summarized in
Table 1. PCR was performed in a reaction volume of 20 µL containing 50 ng of DNA, 1 µL of each primer at 10 µM, and 10 µL of Taq DNA polymerase (Takara Bio Inc., Otsu, Japan). The PCR profiling conditions for the three combinations of primers consisted of denaturation at 94 °C for 5 min, followed by 30 cycles of 94 °C for 30 s, 65 °C for 40 s, and 72 °C for 1 min 30 s, with a final extension at 72 °C for 5 min (F
1 and R
1 primers); denaturation at 94 °C for 5 min, followed by 30 cycles of 94 °C for 30 s, 58 °C for 40 s, and 72 °C for 1 min 30 s, with a final extension at 72 °C for 10 min (F
2 and R
2 primers); and denaturation at 94 °C for 5 min, followed by 30 cycles of 94 °C for 30 s, 54 °C for 40 s, and 72 °C for 1 min 30 s, with a final extension at 72 °C for 5 min (F
3 and R
3 primers).
As a positive PCR control as well as to monitor the integrity of the genomic DNA extracted, we also performed PCR amplification of the 18S ribosomal region. A 348-bp ribosomal fragment was generated by PCR under the above-mentioned conditions using 50 ng of genomic DNA together with the primers F
18S and R
18S, as described by [
32].
2.7. Pollen Viability and Fertility
Pollen viability was tested using the triphenyl tetrazolium chloride (TTC) staining method and a pollen germination in vitro in solid medium method as soon as samples were collected from freshly dehiscent anthers, according to Lu et al. [
33]. In our study, the pollen of transgenic (T-46) and non-transgenic (401) hybrid
P.
tomentosa was tested. Pollen of
P.
tomentosa ×
P.
bolleana (TB16) was also tested as a control.
In the TTC staining method, pollen was stained with TTC (1.0% by weight in 50% sucrose) and was dusted with a brush onto a microscope slide to which four or five drops of stain had been added. A coverslip was immediately placed on the slide, and its edges were sealed with nail varnish. After a 15-min incubation at 40 °C, the pollen was observed under a microscope, and approximately 400 pollen grains from each replicate from the four different areas were counted (three replicates for each staining treatment) to determine pollen viability. The pollen viability by TTC test was expressed with the following formula: Pollen viability (%) = (number of stained pollen/number of observed pollen) × 100%.
For the pollen germination in vitro in solid medium method, pollen was dusted with a brush onto a solid medium (pH 6.0) containing 120 g·L−1 sucrose, 120 mg·L−1 H3BO3, 50 mg·L−1 CaCl2, and 7 g·L−1 agar. The pollen germination rate was detected after incubation at 25 °C for 0.5, 3, and 1 h. The pollen was observed under a microscope, and approximately 400 pollen grains from each replicate from the four different areas were counted (three replicates for each treatment) to determine pollen germination rates. The pollen viability by the pollen germination in vitro in solid medium method was expressed with the following formula: Pollen viability (%) = (number of germinated pollen/number of observed pollen) × 100%.
According to Li and Kang [
34], the stigma receptivity of “
P.
tomentosa ×
P.
bolleana” catkins can be evaluated through their developmental status in the water-cultured condition. When female catkins of “TB04” showed optimal receptivity, they were pollinated with fresh pollen of “TB16”, “401”, and “T-46”. After treatment, the catkins were water-cultured in the greenhouse until seed maturation. Then, seeds were harvested.
4. Discussion
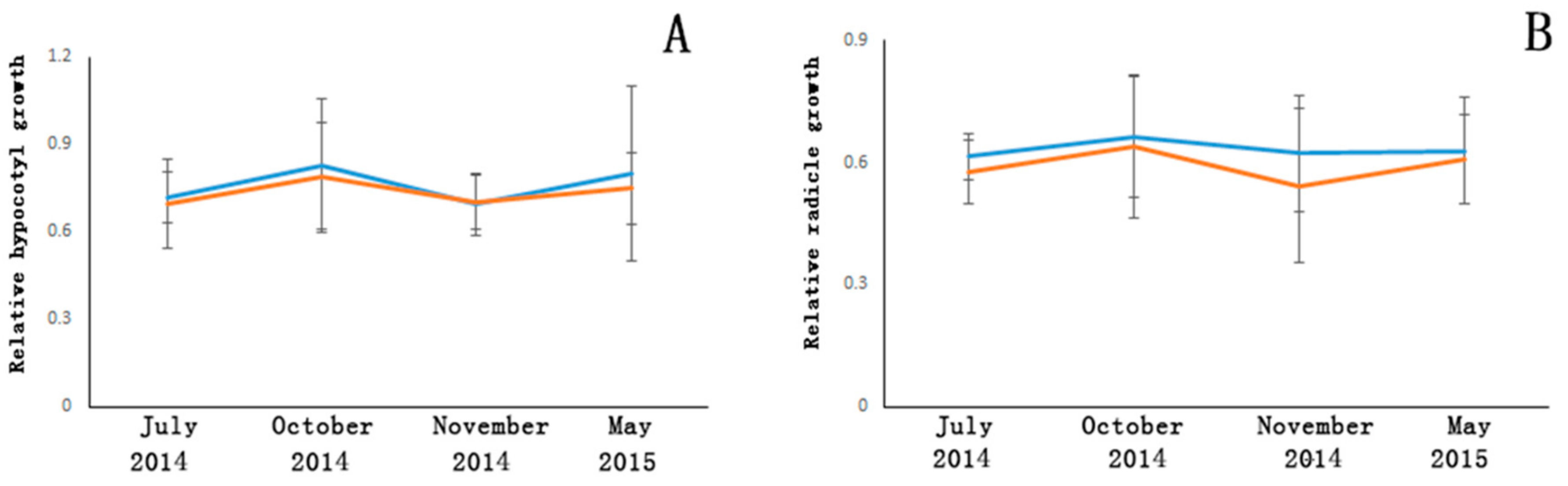
Before transgenic Populus can be commercialized, the possible impacts of transgenic plants on the native ecosystem, such as the rhizosphere microbial community and the surrounding vegetation, must be determined. In our study, we used the sandwich method (and the soil-mix method) and the plate culture method, respectively, to evaluate each of these risks. The impacts of released substances on the allelopathic activity and rhizosphere microorganisms of plants were evaluated by these methods.
In our study, the allelopathic activity of the transgenic
Populus varied with seasonal change, and these changes may have been due to seasonal differences in metabolism. Transgenic and non-transgenic samples showed no significant difference, which indicates that the
AhDREB1 transgenic
Populus may not present risks to surrounding vegetation. Our result was similar to that of Oguchi et al. (2014) [
1], who found in a transgenic
Eucalyptus tree field test that the allelopathic activity of transgenic and non-transgenic trees showed no significant difference by the soil-mix method or the sandwich method. However, the allelopathic activity differed significantly with sampling time in the sandwich method, whereas seasonal periodicity was not found by the soil-mix method. Their study indicated that the experimental method and the sampling timing used to determine allelopathic activity should be considered carefully. In the future, more methods should be tried in allelopathic activity studies.
After cultivation in the field for nine years, we made a comparison between the culturable rhizospheric microorganisms of the transgenic and non-transgenic trees, and significant differences between the numbers of culturable bacteria, actinomycetes, and fungi from the soil samples were not detected between the transgenic and non-transgenic lines (α = 0.05). These results agree with the results of our previous field trials for
AhDREB1-gene transgenic
Populus in 2009 [
31]. These results indicated that expression of the
AhDREB1 gene in
Populus does not affect the number of culturable microorganisms present. Similar results were also shown by Zhu et al. [
35]; in their study, there was no or very limited impact of an eight-year-old transgenic poplar on the bacterial communities in the rhizosphere. However, more than 90% of microorganisms are not culturable [
36]. Hence, only limited numbers of microorganisms were detected in our study, and it remains unclear whether expression of the
AhDREB1 gene in
Populus influenced the overall microorganism diversity in the rhizosphere soil. To develop a sensitive detection method for
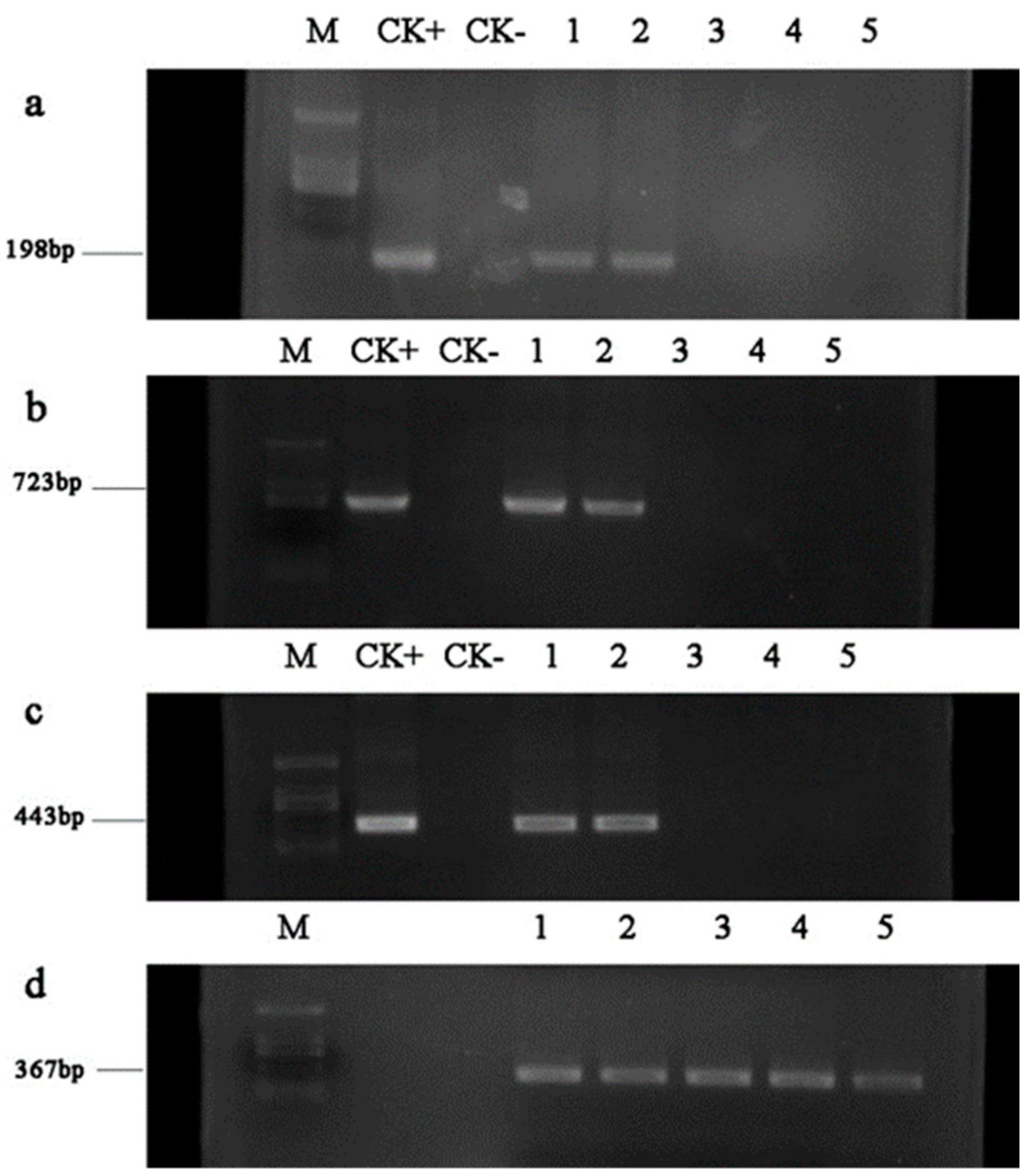
AhDREB1 recombinant sequences from environmental samples, we designed.
PCR primer sets that specifically amplified various regions of the target sequence. To confirm the integrity of the DNA extracted from various samples, we also used a combination of primers to amplify a region of the 18S ribosomal DNA. Our results indicated that recombinant DNA from the transgenic poplar material was digested in as little as three months under natural conditions. In our study, large DNA fragments (723 bp) and small fragments (198 bp) were not detectable at the third month. However, Hay et al. [
24] found that although large recombinant DNA fragments were not detectable by PCR test, smaller target gene fragments could be detected in samples at four months after transgenic poplar leaves had been exposed to external environmental conditions. They suggested that the recombinant DNA sequences may have been cut randomly into shorter fragments over the incubation time. In addition, they also found that the degradation rate of recombinant DNA from transgenic poplar leaf was higher when leaves were placed below the soil than when leaves were placed above the soil or weeds. In their study, the foreign gene was degraded completely in the fourth month, which is slower than in our study, and this difference may be attributable to the host plant in their study being
P.
alba L. ×
P.
grandidentata, rather than (
P.
tomentosa ×
P.
bolleana) ×
P.
tomentosa (this study). Although there is no evidence that degradation rates of recombinant DNA vary among different
Populus species, Widmer and Seidle [
37] found that the recombinant
npt II gene could be detected for up to 77 and 137 days from tobacco and potato transgenic material, respectively, which indicates that the host plant may influence the degradation rate of recombinant DNA. In addition, both soil microbial activity and the activity of free DNase in soil may explain the rapid degradation of DNA; soil type and composition may be other relevant factors. Our study can facilitate the study of possible recombinant DNA from transgenic plant material being available for lateral transfer to soil microorganisms. Our results showed rapid initial degradation of the DNA of a foreign gene in transgenic
Populus and similar results have been obtained in other studies. Hay et al. [
24] thought that because transgenic crop or tree DNA degrades rapidly, possible transfer of recombinant genes from transgenic material to microorganisms is unlikely to occur.
Gene flow is a primary determinant of the potential ecological impacts of transgenic trees, and the possible transgene flow of transgenic
Populus must be considered before large-area planting. According to years of research from our lab,
P.
tomentosa has a very low success rate when hybridized with other
Populus, except for some
P.
tomentosa ×
P.
bolleana and
P.
alba ×
P.
glandulosa lines [
38]. The
P.
tomentosa ×
P.
bolleana “TB04” has proven successful when hybridized with
P.
tomentosa [
39], thus, in our study, we chose “TB04” as the female parent to hybridize with transgenic
Populus. Before the hybridization, we regulated the water culture time of male and female flower branches to ensure that the pollen vitality and stigmatic receptivity were at good levels on the pollination day. If pollen vitality and stigmatic receptivity are not well-known, erroneous conclusions from hybridization experiments are possible. The stigmatic receptivity was best when the stigma was shiny with a large amount of mucus on it and with total cracking. In our study, the stigma was pollinated at the best receptivity day and the pollen was collected one or two days before the pollination. However, in our study, neither the pollen of transgenic (T-46)
Populus nor that of non-transgenic (401)
Populus was viable as determined by TTC test or the pollen germination method, and the result of the hybridization also verified the low pollen fertility. The low pollen fertility of T-46 may be due to the high male sterility of triploid plants. However, Kang and Mao [
39] performed a hybridization using “TB04” as the female parent and the allo-triploid
P.
tomentosa line “B312” [
40] as the male parent and obtained 58 seedlings (gamete fertility was 3.19%), which indicates that triploid may not the main reason of unfertile but low pollen viability of the host plant (401) in 2016. However, we performed hybridization in only one year, and it remains uncertain whether the low pollen fertility is permanent or temporary. Additional experiments to study the pollen fertility will be undertaken in future studies.
It is unfortunate that we had only one transgenic line in our field test (in 2005, we had more than one transgenic line, but the fields of the other lines were destroyed in 2008 and only this one field was conserved). To study the environmental impact of the AhDREB1-gene transgenic Populus further, we conducted another field test in 2016.

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}