Application of Nanoparticle Technologies in the Combat against Anti-Microbial Resistance




Abstract
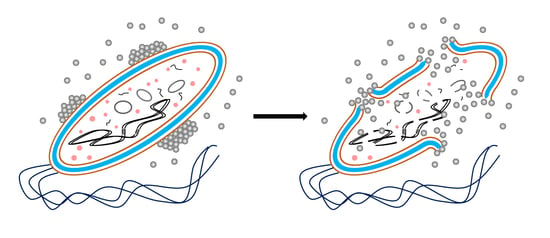
:
1. Introduction
2. Challenges to the Development of New Anti-Microbials
3. Challenges to the Strategies Employed to Develop New Anti-Microbials
4. Nanotechnology in Biomedicine
5. Nanoparticles as Anti-Microbial Agents
5.1. Synergistic Application of Nanoparticles with Antibiotics
5.2. Intrinsic Properties of Nanoparticles Which Confer Activity Against Bacteria
5.2.1. Generation of Reactive Oxidative Species
5.2.2. Effects of Nanoparticles on Bacterial DNA and Metabolism
5.3. Effect of Nanoparticles on Inhibition of Biofilm Formation and Associated Infections
5.4. Physiochemical Properties of Nanoparticles Which Influence Anti-Microbial Activity
5.4.1. Size
5.4.2. Zeta Potential
5.5. Nanoparticles as Drug Carriers
5.6. Nanoparticle Biocompatibility
6. Conclusions and Future Perspectives
Author Contributions
Conflicts of Interest
References
- Ventola, C.L. The Antibiotic Resistance Crisis: Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. The Review on Antimicrobial Resistance. 2016. Available online: https://amr-review.org/Publications.html (accessed on 8 January 2018).
- O’Neill, J. Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations. 2014. Available online: https://amr-review.org/Publications.html (accessed on 8 January 2018).
- World Health Organization. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. 2017. Available online: http://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/ (accessed on 8 December 2017).
- Thomson, R. One Small Step for the Gram Stain, One Giant Leap for Clinical Microbiology. J. Clin. Microbiol. 2016, 54, 1416–1417. [Google Scholar] [CrossRef] [PubMed]
- Navarre, W.W.; Schneewind, O. Surface Proteins of Gram-Positive Bacteria and Mechanisms of Their Targeting to the Cell Wall Envelope. Microbiol. Mol. Biol. Rev. 1999, 63, 174–229. [Google Scholar] [PubMed]
- Kohler, T.; Weidenmaier, C.; Peschel, A. Wall Teichoic Acid Protects Staphylococcus aureus against Anti-microbial Fatty Acids from Human Skin. J. Bacteriol. 2009, 191, 4482–4484. [Google Scholar] [CrossRef] [PubMed]
- Salton, R.J.; Kim, K.S. Structure. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; Chapter 2. [Google Scholar]
- Beveridge, T.J. Structures of Gram-Negative Cell Walls and Their Derived Membrane Vesicles. J. Bacteriol. 1999, 181, 4725–4733. [Google Scholar] [PubMed]
- Cooper, B.; Medley, G.; Stone, S.; Kibbler, C.; Cookson, B.; Roberts, J.; Duckworth, G.; Lai, R.; Ebrahim, S. Methicillin-resistant Staphylococcus aureus in hospitals and the community: Stealth dynamics and control catastrophes. PNAS 2004, 101, 10223–10228. [Google Scholar] [CrossRef] [PubMed]
- Kidd, T.; Mills, G.; Sá-Pessoa, J.; Dumigan, A.; Frank, C.; Insua, J.; Ingram, R.; Hobley, L.; Bengoechea, J. A Klebsiella pneumoniae antibiotic resistance mechanism that subdues host defences and promotes virulence. EMBO Mol. Med. 2017, 9, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Meletis, G. Carbapenem resistance: Overview of the problem and future perspectives. Ther. Adv. Infect. Dis. 2016, 3, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Zumla, A.; Schito, M.; Chakaya, J.; Marais, B.; Mwaba, P.; Migliori, G.; Hoelscher, M.; Maeurer, M.; Wallis, R. World TB Day 2016: Reflections on the global TB emergency. Lancet Respir. Med. 2016, 4, 249–251. [Google Scholar] [CrossRef]
- Udwadia, Z.; Amale, R.; Ajbani, K.; Rodrigues, C. Totally Drug Resistant Tuberculosis on India. Clin. Infect. Dis. 2012, 54, 579–581. [Google Scholar] [CrossRef] [PubMed]
- Miesel, L.; Greene, J.; Black, T. Microbial genetics: Genetic strategies for antibacterial drug discovery. Nat. Rev. Genet. 2003, 4, 442–456. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Park, Y.; Chung, E.; Na, E.; Ko, K. Evolved resistance to colistin and its loss due to genetic reversion in Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 25543. [Google Scholar] [CrossRef] [PubMed]
- Gold, H. Vancomycin-Resistant Enterococci: Mechanisms and Clinical Observations. Clin. Infect. Dis. 2001, 33, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.; Makarova, K.; Aravind, L. Horizontal Gene Transfer in Prokaryotes: Quantification and Classification. Ann. Rev. Microbiol. 2001, 55, 709–742. [Google Scholar] [CrossRef] [PubMed]
- Furuya, E.; Lowy, F. Antimicrobial-resistant bacteria in the community setting. Nat. Rev. Microbiol. 2006, 4, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Fomda, B.A.; Khan, A.; Zahoor, D. NDM-1 (New Delhi metallo beta lactamase-1) producing Gram-negative bacilli: Emergence & clinical implications. Indian J. Med. Res. 2014, 140, 672–678. [Google Scholar] [PubMed]
- Périchon, B.; Courvalin, P. VanA-Type Vancomycin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2009, 53, 4580–4587. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hu, C.; Shao, L. The anti-microbial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2017, 12, 1227–1249. [Google Scholar] [CrossRef] [PubMed]
- DiMasi, J.; Grabowski, H.; Hansen, R. Innovation in the pharmaceutical industry: New estimates of R&D costs. J. Health Econ. 2016, 47, 20–33. [Google Scholar] [PubMed]
- Devasahayam, G.; Scheld, W.; Hoffman, P. Newer anti-bacterial drugs for a new century. Exp. Opin. Investig. Drugs 2010, 19, 215–234. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.; Guidos, R.; Gilbert, D.; Bradley, J.; Boucher, H.; Scheld, W.; Bartlett, J.; Edwards, J. The Epidemic of Antibiotic-Resistant Infections: A Call to Action for the Medical Community from the Infectious Diseases Society of America. Clin. Infect. Dis. 2008, 46, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Power, E. Impact of antibiotic restrictions: The pharmaceutical perspective. Clin. Microbiol. Infect. 2006, 12, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Davies, J. Where have All the Antibiotics Gone? Can. J. Infect. Dis. Med. Microbiol. 2006, 17, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Biedenbach, D.; Bouchillon, S.; Hackel, M.; Miller, L.; Scangarella-Oman, N.; Jakielaszek, C.; Sahm, D. In vitro Activity of Gepotidacin, a Novel Triazaacenaphthylene Bacterial Topoisomerase Inhibitor, Against a Broad Spectrum of Bacterial Pathogens. Antimicrob. Agents Chemother. 2016, 60, 1918–1923. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Antibacterial Agents in Clinical Development: An Analysis of the Antibacterial Clinical Development Pipeline, Including Tuberculosis; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Lobanovska, M.; Pilla, G. Penicillin’s Discovery and Antibiotic Resistance: Lessons for the Future? Yale J. Biol. Med. 2017, 90, 135–145. [Google Scholar] [PubMed]
- Cassir, N.; Rolain, J.; Brouqui, P. A new strategy to fight anti-microbial resistance: The revival of old antibiotics. Front. Microbiol. 2014, 5, 551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khadka, P.; Ro, J.; Kim, H.; Kim, I.; Kim, J.; Kim, H.; Cho, J.; Yun, G.; Lee, J. Pharmaceutical particle technologies: An approach to improve drug solubility, dissolution and bioavailability. Asian J. Pharm. Sci. 2014, 9, 304–316. [Google Scholar] [CrossRef]
- Rodzinski, A.; Guduru, R.; Liang, P.; Hadjikhani, A.; Stewart, T.; Stimphil, E.; Runowicz, C.; Cote, R.; Altman, N.; Datar, R.; et al. Targeted and controlled anticancer drug delivery and release with magnetoelectric nanoparticles. Sci. Rep. 2016, 6, 20867. [Google Scholar] [CrossRef] [PubMed]
- Shahverdi, A.; Fakhimi, A.; Shahverdi, H.; Minaian, S. Synthesis and effect of silver nanoparticles on the anti-bacterial activity of different antibiotics against Staphylococcus aureus and Escherichia coli. Nanomed. Nanotechnol. Biol. Med. 2007, 3, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; McShan, D.; Zhang, Y.; Sinha, S.S.; Arslan, Z.; Ray, P.C.; Yu, H. Mechanistic Study of the Synergistic Anti-bacterial Activity of Combined Silver Nanoparticles and Common Antibiotics. Environ. Sci. Technol. 2016, 50, 8840–8848. [Google Scholar] [CrossRef] [PubMed]
- Banoee, M.; Seif, S.; Nazari, Z.; Jafari-Fesharaki, P.; Shahverdi, H.; Moballegh, A.; Moghaddam, K.; Shahverdi, A. ZnO nanoparticles enhanced anti-bacterial activity of ciprofloxacin against Staphylococcus aureus and Escherichia coli. J. Biomed. Mater. Res. B 2010, 93B, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. Cellular defenses against superoxide and hydrogen peroxide. Ann. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef] [PubMed]
- Iuchi, S.; Weiner, L. Cellular and Molecular Physiology of Escherichia coli in the Adaptation to Aerobic Environments. J. Biochem. 1996, 120, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Glaeser, J.; Nuss, A.M.; Berghoff, B.A.; Klug, G. Singlet oxygen stress in microorganisms. Adv. Microb. Phys. 2011, 58, 141–173. [Google Scholar]
- Li, Y.; Zhang, W.; Niu, J.; Chen, Y. Mechanism of Photogenerated Reactive Oxygen Species and Correlation with the Anti-bacterial Properties of Engineered Metal-Oxide Nanoparticles. ACS Nano 2012, 6, 5164–5173. [Google Scholar] [CrossRef] [PubMed]
- Reddy, L.; Nisha, M.; Joice, M.; Shilpa, P. Anti-microbial activity of zinc oxide (ZnO) nanoparticle againstKlebsiella pneumoniae. Pharm. Biol. 2014, 52, 1388–1397. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, S.; Wahab, R.; Khan, F.; Mishra, Y.K.; Musarrat, J.; Al-Khedhairy, A.A. Reactive Oxygen Species Mediated Bacterial Biofilm Inhibition via Zinc Oxide Nanoparticles and Their Statistical Determination. PLoS ONE 2014, 9, e111289. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Chakraborty, R.; Basu, T. Mechanism of anti-bacterial activity of copper nanoparticles. Nanotechnology 2014, 25, 135101. [Google Scholar] [CrossRef] [PubMed]
- Padmavathy, N.; Vijayaraghavan, R. Interaction of ZnO Nanoparticles with Microbes-A Physio and Biochemical Assay. J. Biomed. Nanotechnol. 2011, 7, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Manikprabhu, D.; Lingappa, K. Anti-bacterial activity of silver nanoparticles against methicillin-resistant Staphylococcus aureus synthesized using model Streptomyces sp. pigment by photo-irradiation method. J. Pharm. Res. 2013, 6, 255–260. [Google Scholar]
- Xie, Y.; He, Y.; Irwin, P.; Jin, T.; Shi, X. Anti-bacterial Activity and Mechanism of Action of Zinc Oxide Nanoparticles against Campylobacter jejuni. Appl. Environ. Microbiol. 2011, 77, 2325–2331. [Google Scholar] [CrossRef] [PubMed]
- Raffi, M.; Mehrwan, S.; Bhatti, T.; Akhter, J.; Hameed, A.; Yawar, W.; ul Hasan, M. Investigations into the anti-bacterial behavior of copper nanoparticles against Escherichia coli. Ann. Microbiol. 2010, 60, 75–80. [Google Scholar] [CrossRef]
- Arakha, M.; Pal, S.; Samantarrai, D.; Panigrahi, T.; Mallick, B.; Pramanik, K.; Mallick, B.; Jha, S. Anti-microbial activity of iron oxide nanoparticle upon modulation of nanoparticle-bacteria interface. Sci. Rep. 2015, 5, 14813. [Google Scholar] [CrossRef] [PubMed]
- Salomoni, R.; Léo, P.; Montemor, A.; Rinaldi, B.; Rodrigues, M. Anti-bacterial effect of silver nanoparticles in Pseudomonas aeruginosa. Nanotechnol. Sci. Appl. 2017, 10, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, Y.; Lin, K.; Ke, W.; Hsieh, C.; Chiang, C.; Tzou, D.; Liu, S. The Anti-microbial Properties of Silver Nanoparticles in Bacillus subtilis are Mediated by Released Ag+ Ions. PLoS ONE 2015, 10, e0144306. [Google Scholar] [CrossRef] [PubMed]
- Laursen, B.S.; Sørensen, H.P.; Mortensen, K.K.; Sperling-Petersen, H.U. Initiation of Protein Synthesis in Bacteria. Microbiol. Mol. Biol. Rev. 2005, 69, 101–123. [Google Scholar] [CrossRef] [PubMed]
- Shamaila, S.; Zafar, N.; Riaz, S.; Sharif, R.; Nazir, J.; Naseem, S. Gold Nanoparticles: An Efficient Anti-microbial Agent against Enteric Bacterial Human Pathogen. Nanomaterials 2016, 6, 71. [Google Scholar] [CrossRef] [PubMed]
- Health Protection Agency (HPA). Healthcare-Associated Infection and Anti-Microbial Resistance: 2010–2011; HPA: London, UK, 2012.
- Soto-Giron, M.; Rodriguez-R, L.; Luo, C.; Elk, M.; Ryu, H.; Hoelle, J.; Santo Domingo, J.; Konstantinidis, K. Biofilms on Hospital Shower Hoses: Characterization and Implications for Nosocomial Infections. Appl. Environ. Microbiol. 2016, 82, 2872–2883. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R. Biofilms: Microbial Life on Surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. AntiMicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L. Microcolony and biofilm formation as a survival strategy for bacteria. J. Theor. Biol. 2008, 251, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Keenan, C.; Sedlak, D. Factors Affecting the Yield of Oxidants from the Reaction of Nanoparticulate Zero-Valent Iron and Oxygen. Environ. Sci. Technol. 2008, 42, 1262–1267. [Google Scholar] [CrossRef] [PubMed]
- Chávez de Paz, L.E.; Resin, A.; Howard, K.A.; Sutherland, D.S.; Wejse, P.L. Anti-microbial Effect of Chitosan Nanoparticles on Streptococcus mutans Biofilms. Appl. Environ. Microbiol. 2011, 77, 3892–3895. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Jia, J.; Guo, X.; Zhao, Y.; Chen, D.; Guo, Y.; Zhang, X. Reduced Staphylococcus aureus biofilm formation in the presence of chitosan-coated iron oxide nanoparticles. Int. J. Nanomed. 2016, 11, 6499–6506. [Google Scholar] [CrossRef] [PubMed]
- Shrifian-Esfahni, A.; Salehi, M.; Nasr-Esfahni, M.; Ekramian, E. Chitosan-modifed superparamagnetic iron oxide nanoparticles: Design, fabrication, characterization and antibacterial activity. CHEMIK 2015, 69, 19–32. [Google Scholar]
- Jesline, A.; John, N.; Narayanan, P.; Vani, C.; Murugan, S. Anti-microbial activity of zinc and titanium dioxide nanoparticles against biofilm-producing methicillin-resistant Staphylococcus aureus. Appl. Nanosci. 2014, 5, 157–162. [Google Scholar] [CrossRef]
- Roy, A.; Parveen, A.; Koppalkar, A.R.; Prasad, M. Effect of Nano—Titanium Dioxide with Different Antibiotics against Methicillin-Resistant Staphylococcus Aureus. J. Biomater. Nanobiotechnol. 2010, 1, 37–41. [Google Scholar] [CrossRef]
- Cao, H.; Qin, H.; Zhao, Y.; Jin, G.; Lu, T.; Meng, F.; Zhang, X.; Liu, X. Nano-thick calcium oxide armed titanium: Boosts bone cells against methicillin-resistant Staphylococcus aureus. Sci. Rep. 2016, 6, 21761. [Google Scholar] [CrossRef] [PubMed]
- Morones, J.; Elechiguerra, J.; Camacho, A.; Holt, K.; Kouri, J.; Ramírez, J.; Yacaman, M. The bactericidal effect of silver nanoparticles. Nanotechnology 2005, 16, 2346–2353. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, B.; Garmaroudi, F.; Hashemi, M.; Nezhad, H.; Nasrollahi, A.; Ardalan, S.; Ardalan, S. Comparison of the anti-bacterial activity on the nanosilver shapes: Nanoparticles, nanorods and nanoplates. Adv. Powder Technol. 2012, 23, 22–26. [Google Scholar] [CrossRef]
- Ayala, V.; Herrera, A.; Latorre-Esteves, M.; Torres-Lugo, M.; Rinaldi, C. Effect of surface charge on the colloidal stability and in vitro uptake of carboxymethyl dextran-coated iron oxide nanoparticles. J. Nanopart. Res. 2013, 15, 1874. [Google Scholar] [CrossRef] [PubMed]
- Halder, S.; Yadav, K.; Sarkar, R.; Mukherjee, S.; Saha, P.; Haldar, S.; Karmakar, S.; Sen, T. Alteration of Zeta potential and membrane permeability in bacteria: A study with cationic agents. SpringerPlus 2015, 4, 672. [Google Scholar] [CrossRef] [PubMed]
- Meléndrez, M.; Cárdenas, G.; Arbiol, J. Synthesis and characterization of gallium colloidal nanoparticles. J. Colloid Interface Sci. 2010, 346, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, K.H.; Gunsolus, I.L.; Kuech, T.R.; Troiano, J.M.; Melby, E.S.; Lohse, S.E.; Hu, D.; Chrisler, W.B.; Murphy, C.J.; Orr, G.; et al. Lipopolysaccharide Density and Structure Govern the Extent and Distance of Nanoparticle Interaction with Actual and Model Bacterial Outer Membranes. Environ. Sci. Technol. 2015, 49, 10642–10650. [Google Scholar] [CrossRef] [PubMed]
- Snyder, D.; McIntosh, T. The Lipopolysaccharide Barrier: Correlation of Antibiotic Susceptibility with Antibiotic Permeability and Fluorescent Probe Binding Kinetics. Biochemistry 2000, 39, 11777–11787. [Google Scholar] [CrossRef] [PubMed]
- Bolla, J.-M.; Alibert-Franco, S.; Handzlik, J.; Chevalier, J.; Mahamoud, A.; Boyer, G.; Kieć-Kononowicz, K.; Pagès, J.-M. Strategies for bypassing the membrane barrier in multidrug resistant Gram-negative bacteria. FEBS Lett. 2011, 585, 1682–1690. [Google Scholar] [CrossRef] [PubMed]
- Arakha, M.; Saleem, M.; Mallick, B.; Jha, S. The effects of interfacial potential on anti-microbial propensity of ZnO nanoparticle. Sci. Rep. 2015, 5, 9578. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, H.; Guo, G.; Cheng, T.; Peng, X.; Mao, X.; Li, J.; Zhang, X. A functionalized surface modification with vanadium nanoparticles of various valences against implant-associated bloodstream infection. Int. J. Nanomed. 2017, 12, 3121–3136. [Google Scholar] [CrossRef] [PubMed]
- Begg, E.J.; Barclay, M.L.; Kirkpatrick, C.J.M. The therapeutic monitoring of anti-microbial agents. Br. J. Clin. Pharmacol. 1999, 47, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Triggs, E.; Charles, B. Pharmacokinetics and Therapeutic Drug Monitoring of Gentamicin in the Elderly. Clin. Pharmacokinet. 1999, 37, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.; Bhattacharya, J.; Mukherjee, A.; Ghosh, A.; Santra, C.; Dasgupta, A.; Karmakar, P. In Vitro Structural and Functional Evaluation of Gold Nanoparticles Conjugated Antibiotics. Nanoscale Res. Lett. 2007, 2, 614–622. [Google Scholar] [CrossRef]
- Brown, A.N.; Smith, K.; Samuels, T.A.; Lu, J.; Obare, S.O.; Scott, M.E. Nanoparticles functionalized with ampicillin destroy multi-antibiotic-resistant isolates of Pseudomonas aerunginosa and Enterobacter aerogenes and methicillin-resistant Staphylococcus aureus. Appl. Environ. Microbiol. 2012, 78, 2768–2774. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.H.; Ranzoni, A.; Phetsang, W.; Blaskovich, M.A.T.; Cooper, M.A. Surface ligand density of anti-biotic-nanoparticle conjugates enhances target avidity and membrane permabilization of vancomycin-resistant bacteria. Bioconjug. Chem. 2017, 28, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Turos, E.; Shim, J.-Y.; Wang, Y.; Greenhalgh, K.; Suresh, G.; Reddy, K.; Dickey, S.; Lim, D.V. Antibiotic-conjugated polyacrylate nanoparticles: New opportunities for development of anti-MRSA agents. Bioorg. Med. Chem. Lett. 2007, 17, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.-K.; Selvanayagam, R.; Ho, K.K.K.; Chen, R.; Kutty, S.K.; Rice, S.A.; Kumar, N.; Barraud, N.; Duong, H.T.T.; Boyer, C. Co-delivery of nitric oxide and antibiotic using polymeric nanoparticles. Chem. Sci. 2016, 7, 1016–1027. [Google Scholar] [CrossRef] [PubMed]
- Hammes, W.P.; Neuhaus, F.C. On the Mechanism of Action of Vancomycin: Inhibition of Peptidoglycan Synthesis in Gaffkya homari. Antimicrob. Agents Chemother. 1974, 6, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Marshall, C.; Broadhead, G.; Leskiw, B.; Wright, G. D-Ala-D-Ala ligases from glycopeptide antibiotic-producing organisms are highly homologous to the enterococcal vancomycin-resistance ligases VanA and VanB. PNAS 1997, 94, 6480–6483. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D. Microbial genetics: Exploiting genomics, genetics and chemistry to combat antibiotic resistance. Nat. Rev. Genet. 2003, 4, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Musser, J.; DeLeo, F. Genomic Analysis of the Emergence of Vancomycin-Resistant Staphylococcus aureus. mBio 2012, 3, e00170-12. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Sahu, S.; Mahapatra, S.; Santra, S.; Bal, M.; Roy, S.; Pramanik, P. Nanoconjugated vancomycin: New opportunities for the development of anti-VRSA agents. Nanotechnology 2010, 21, 105103. [Google Scholar] [CrossRef] [PubMed]
- Abdelghany, S.; Quinn, D.; Ingram, R.; Gilmore, B.; Donnelly, R.; Taggart, C.; Scott, C. Gentamicin-loaded nanoparticles show improved anti-microbial effects towards Pseudomonas aeruginosa infection. Int. J. Nanomed. 2012, 7, 4053–4063. [Google Scholar]
- Smith, I. Mycobacterium tuberculosis Pathogenesis and Molecular Determinants of Virulence. Clin. Microbiol. Rev. 2003, 16, 463–496. [Google Scholar] [CrossRef] [PubMed]
- Kalluru, R.; Fenaroli, F.; Westmoreland, D.; Ulanova, L.; Maleki, A.; Roos, N.; Paulsen Madsen, M.; Koster, G.; Egge-Jacobsen, W.; Wilson, S.; et al. Poly(lactide-co-glycolide)-rifampicin nanoparticles efficiently clear Mycobacterium bovis BCG infection in macrophages and remain membrane-bound in phago-lysosomes. J. Cell Sci. 2013, 126, 3043–3054. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
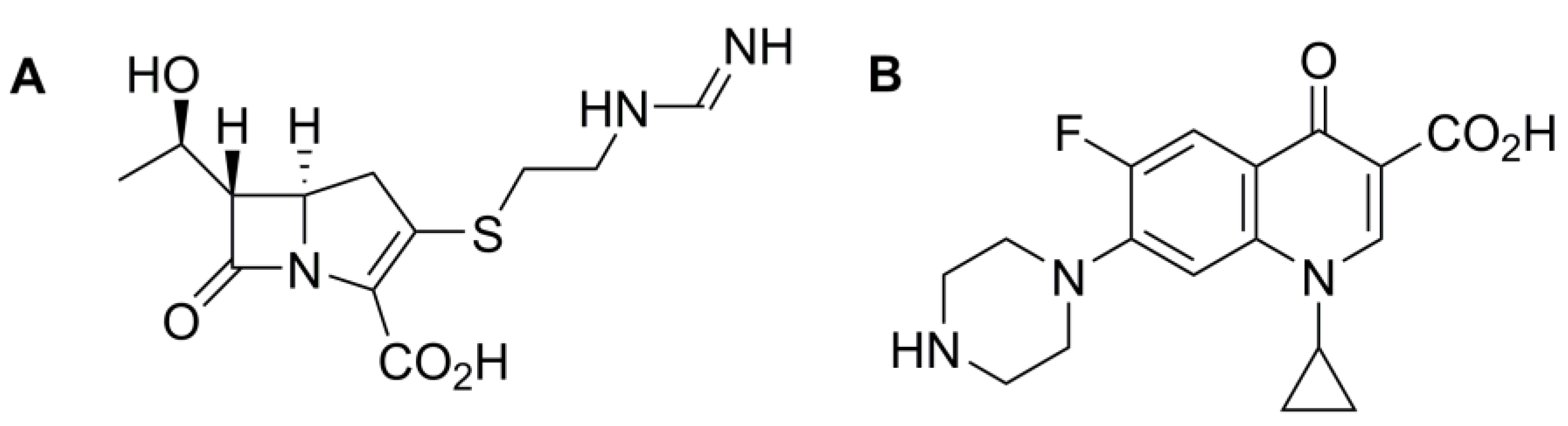{kind=link}
{kind=link}
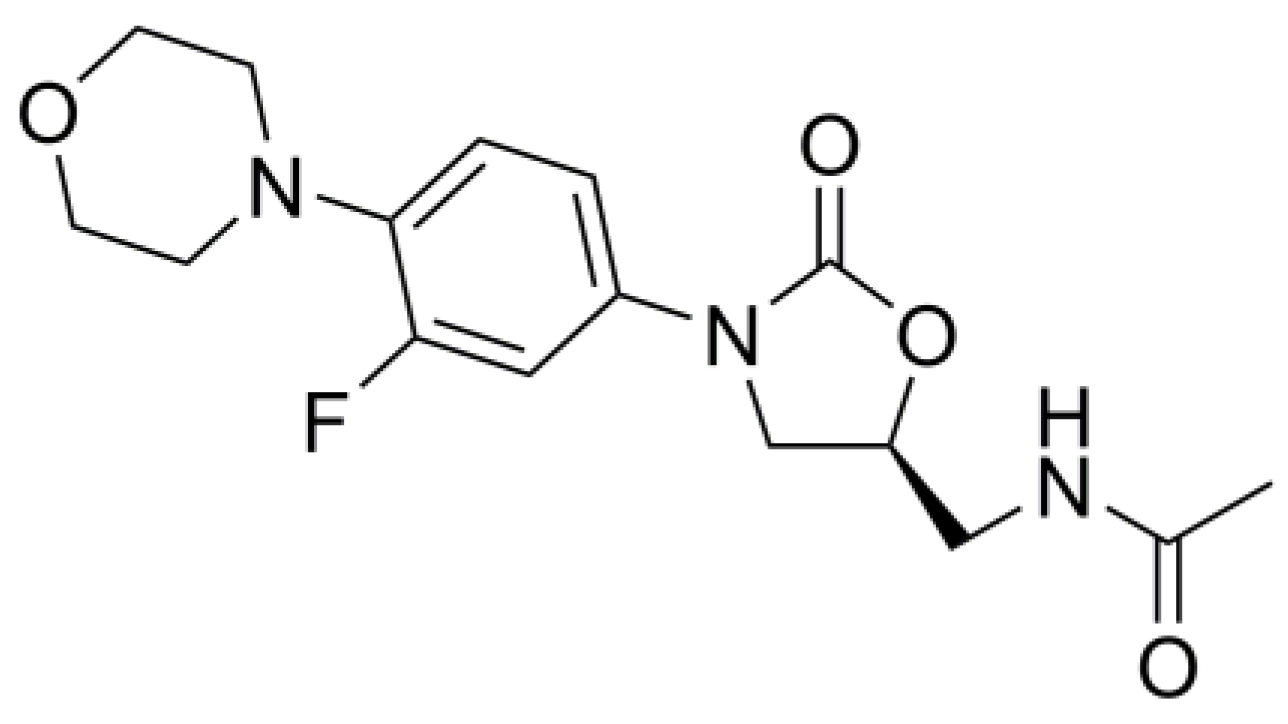{kind=link}
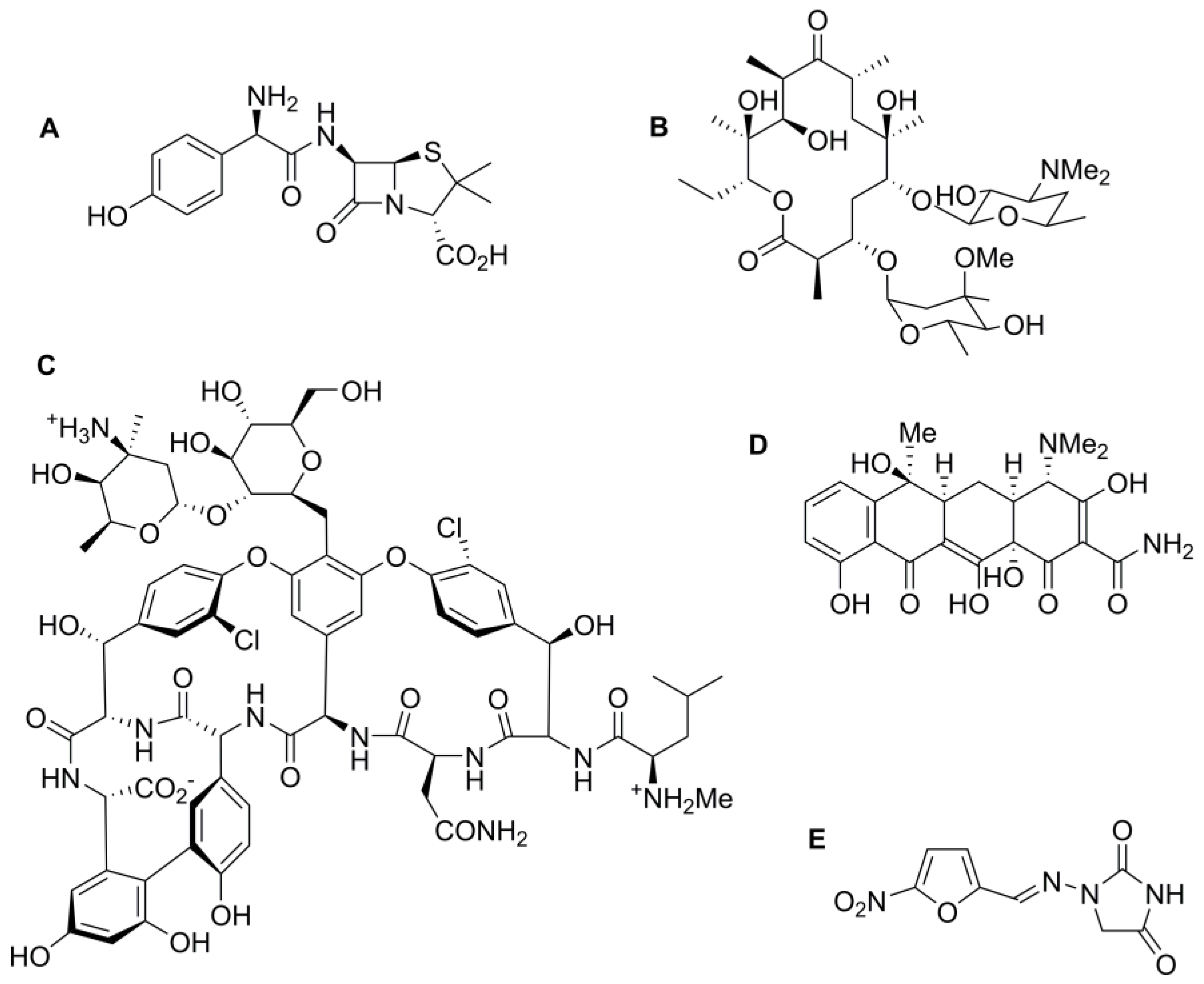{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Species | Gram Stain of the Species | Nanoparticle(s) with Anti-Microbial Activity | Mechanism of Action | Reference |
|---|---|---|---|---|
| Staphylococcus aureus | Gram-positive | Ag | Cell wall damage | [46] |
| Campylobacter jejuni | Gram-negative | ZnO | Growth inhibition by ROS | [47] |
| Escherichia coli | Gram-negative | Cu IONPs | Growth inhibition by ROS ROS generation | [48,49] |
| Pseudomonas aeruginosa | Gram-negative | Ag Zn | Enzyme inhibition in respiratory chain complex ROS generation | [43,50] |
| Klebsiella pneumoniae | Gram-negative | ZnO | Growth inhibition | [42] |
| Bacillus subtilis | Gram-positive | Ag IONPs | DNA degradation ROS generation | [49,51] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, M.; Curtis, A.; Hoskins, C. Application of Nanoparticle Technologies in the Combat against Anti-Microbial Resistance. Pharmaceutics 2018, 10, 11. https://doi.org/10.3390/pharmaceutics10010011
Kumar M, Curtis A, Hoskins C. Application of Nanoparticle Technologies in the Combat against Anti-Microbial Resistance. Pharmaceutics. 2018; 10(1):11. https://doi.org/10.3390/pharmaceutics10010011
Chicago/Turabian StyleKumar, Mayur, Anthony Curtis, and Clare Hoskins. 2018. "Application of Nanoparticle Technologies in the Combat against Anti-Microbial Resistance" Pharmaceutics 10, no. 1: 11. https://doi.org/10.3390/pharmaceutics10010011
APA StyleKumar, M., Curtis, A., & Hoskins, C. (2018). Application of Nanoparticle Technologies in the Combat against Anti-Microbial Resistance. Pharmaceutics, 10(1), 11. https://doi.org/10.3390/pharmaceutics10010011