Impact of Glycosylation and Species Origin on the Uptake and Permeation of IgGs through the Nasal Airway Mucosa

 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. IgGs Used in This Study
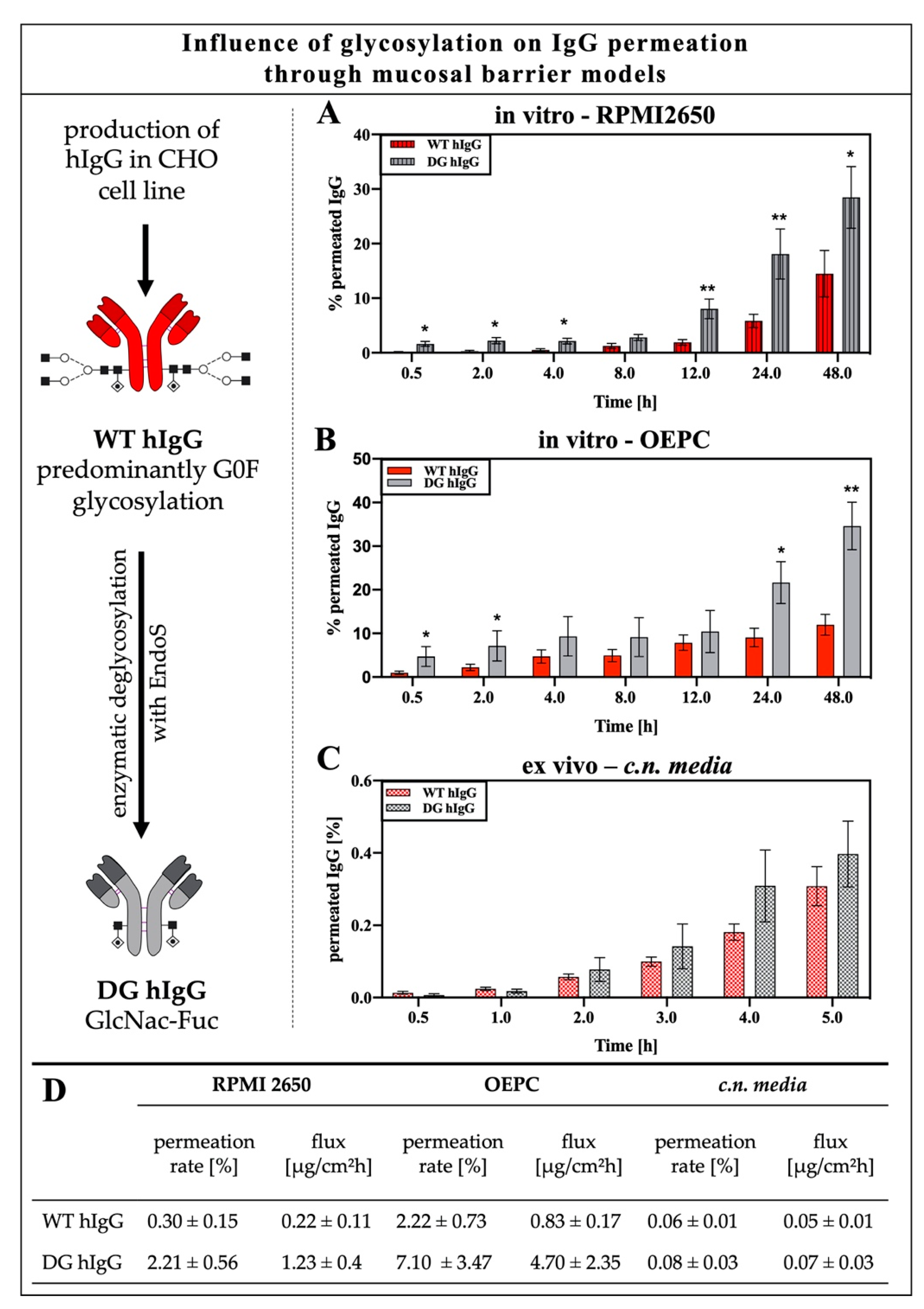
2.2. IgG Deglycosylation
2.3. Tissue Preparation and Ex Vivo Permeation
2.4. Cell Culture
2.4.1. Primary Cells
2.4.2. RPMI 2650
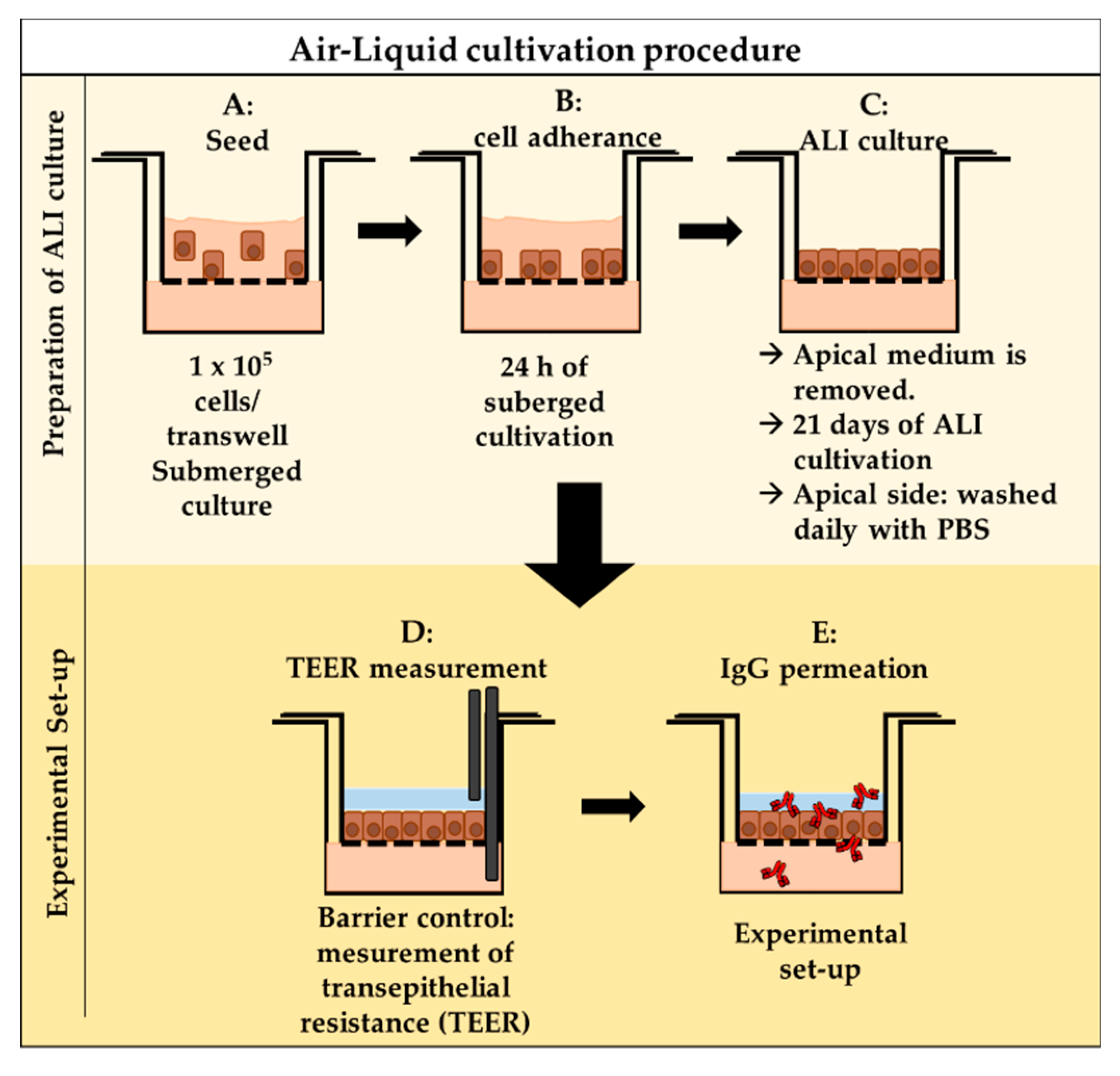
2.4.3. Air–Liquid Interface (ALI) Cell Culture
2.5. In Vitro Permeation
2.5.1. TEER Measurement
2.5.2. In Vitro Permeation—Experimental Procedure
2.6. Immunofluorescence Staining of Tissue Explants and Cell Culture Insert Membranes
2.7. Western Blot Analysis
2.8. Immunoprecipitation
2.9. Statistics
3. Results
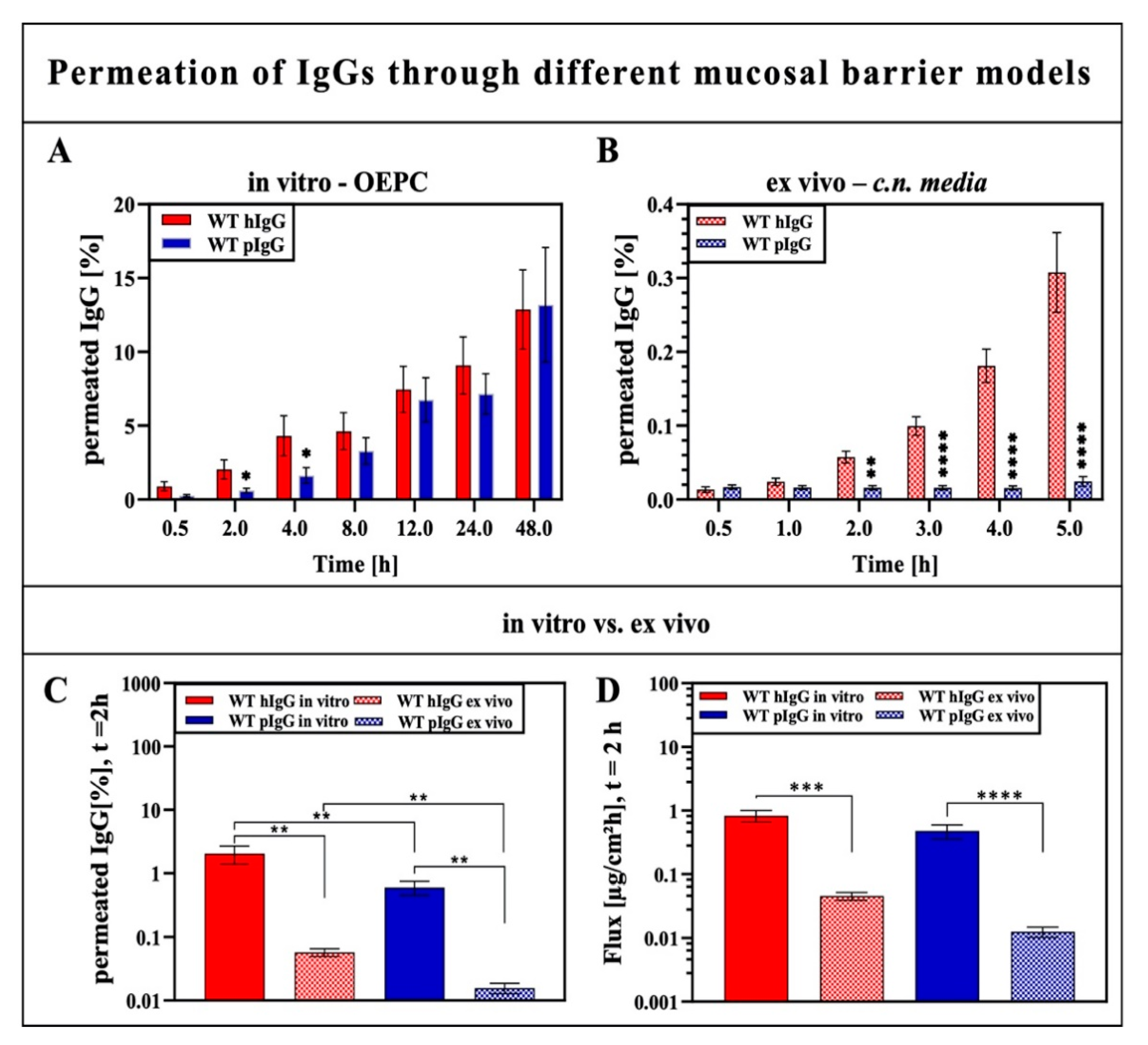
3.1. IgG Permeation through Different Models of the Olfactory Mucosa and Olfactory Epithelium
3.1.1. Impact of IgG Species Origin on Trans-Epithelial Permeation of IgGs
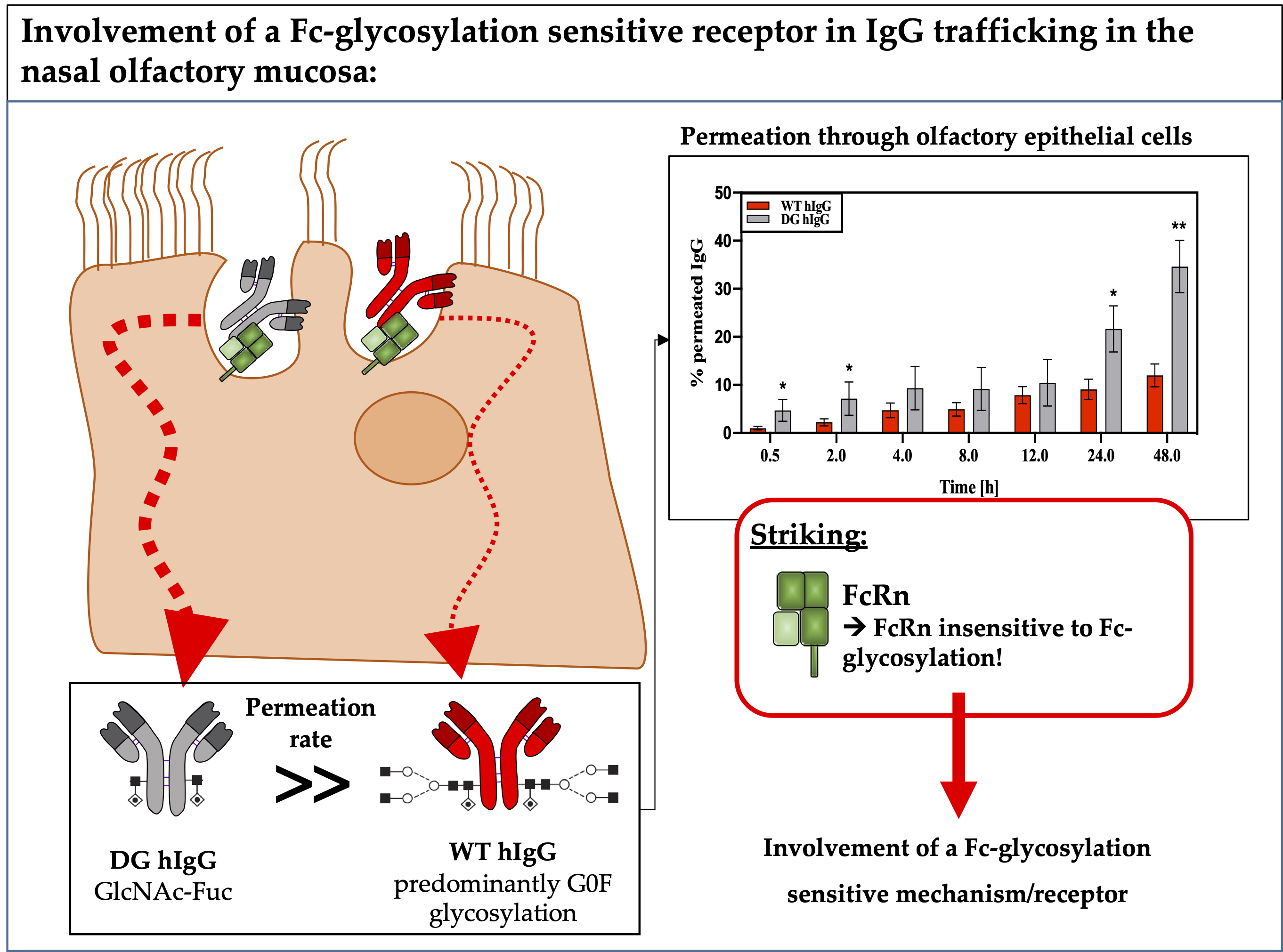
3.1.2. Impact of IgG Glycosylation on Permeation through the Nasal Epithelium or Nasal Mucosa
3.2. Analysis of IgG Transporter in Porcine Nasal Mucosa, Primary Epithilial Cells and the RPMI 2650 Model
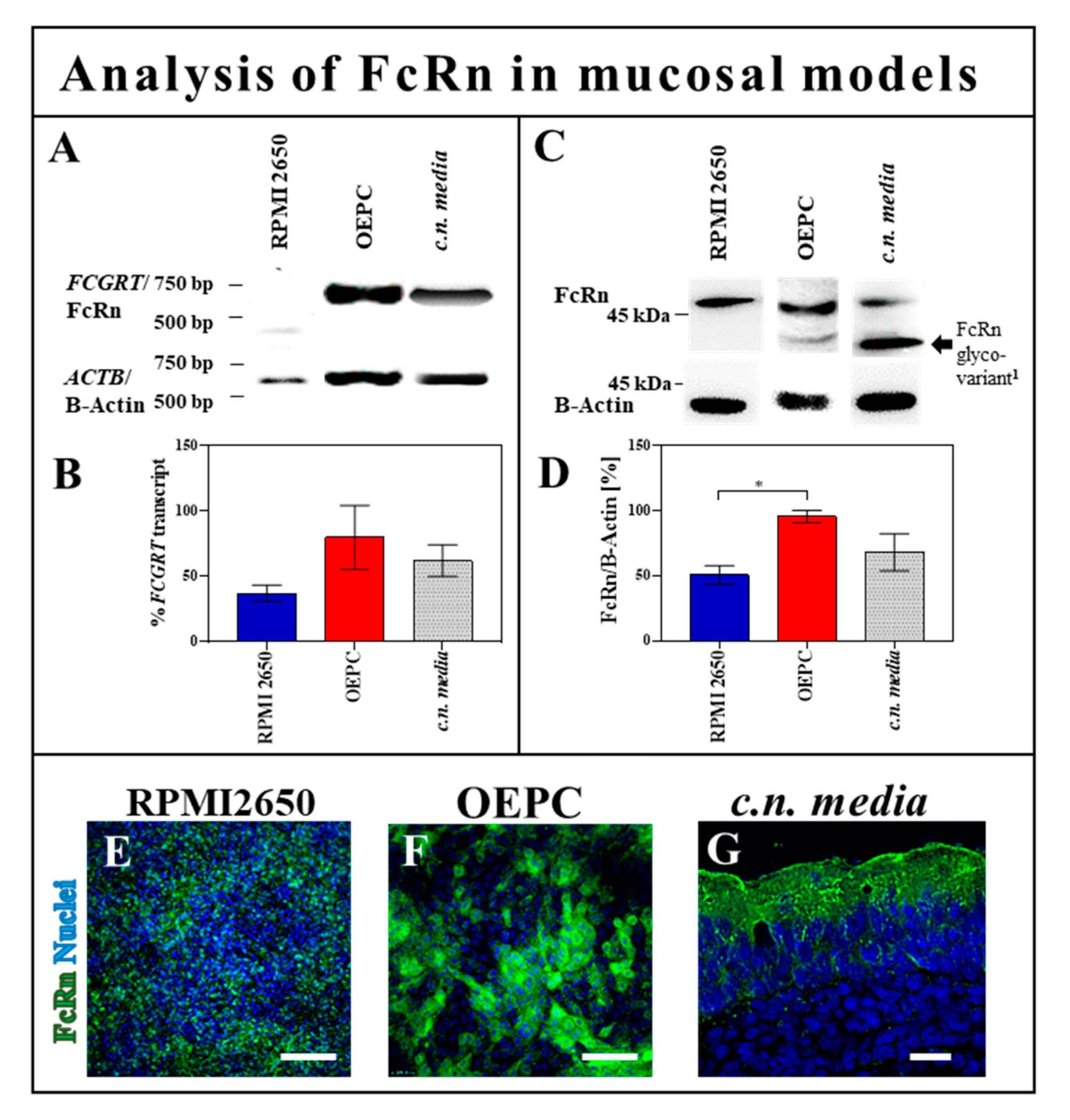
3.2.1. FcRn Expression in OEPC and RPMI 2650
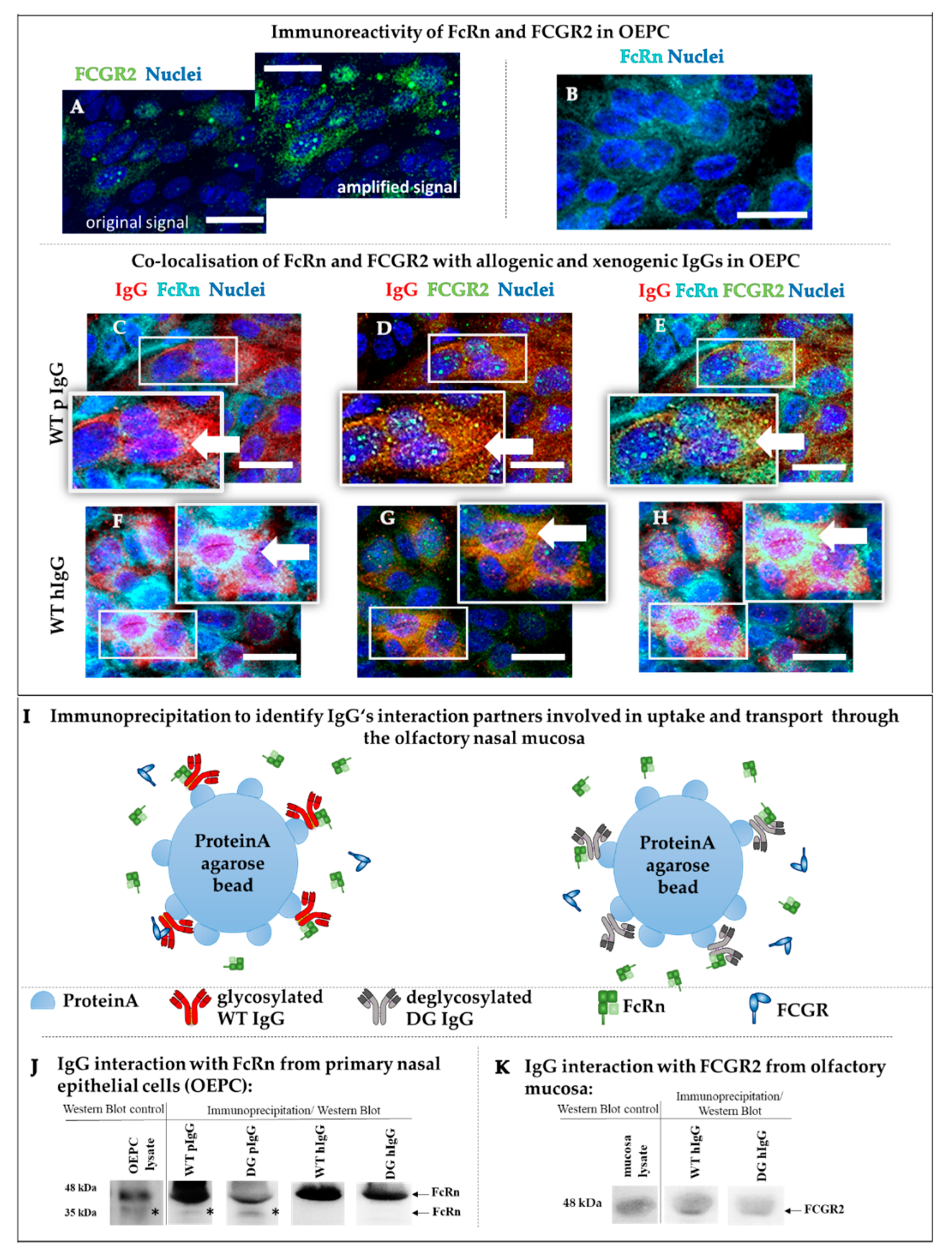
3.2.2. Protein Interaction Study of FcRn and FCGR2 with Wild-Type and Deglycosylated IgG
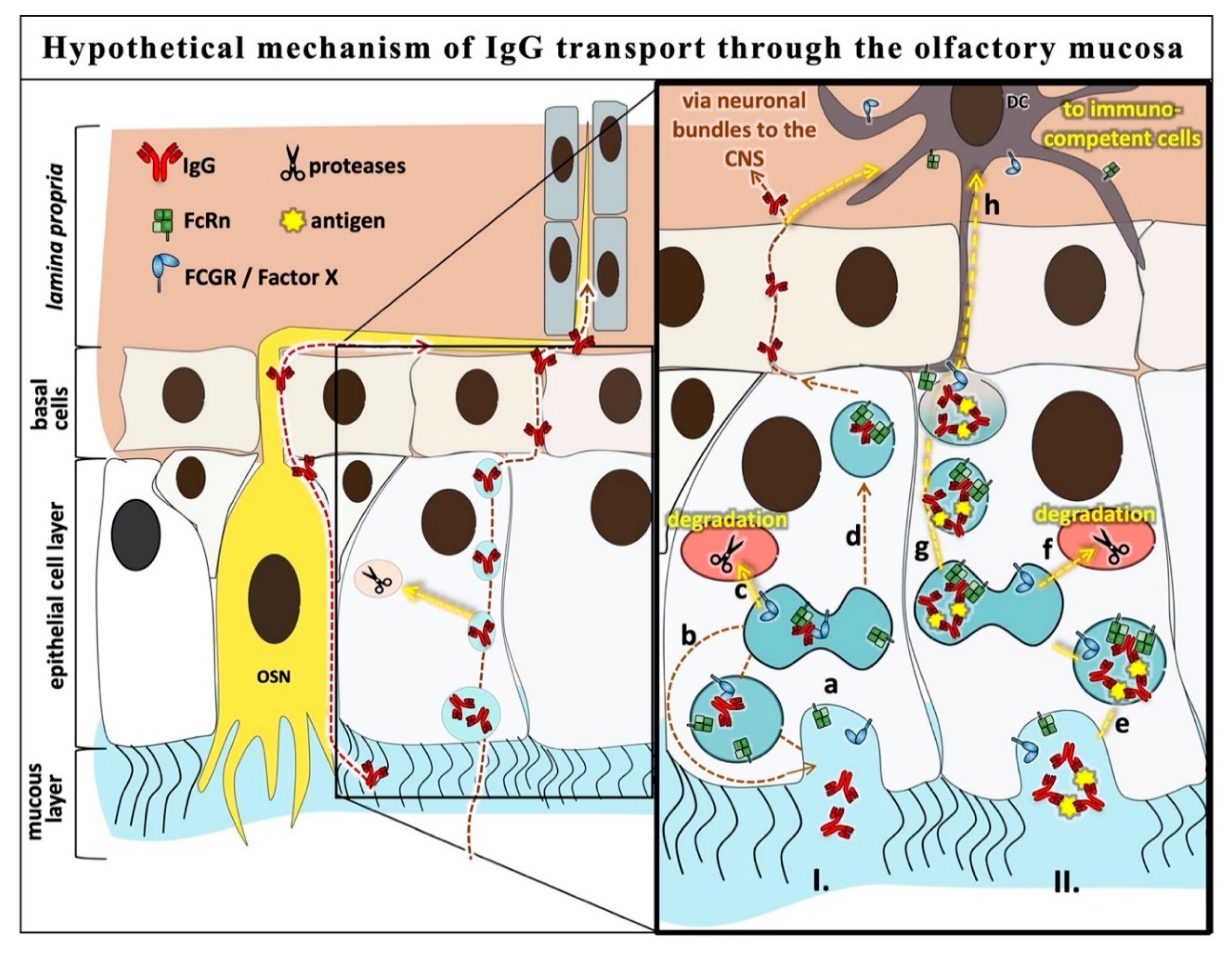
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kaplon, H.; Reichert, J.M. Antibodies to watch in 2019. MAbs 2019, 11, 219–238. [Google Scholar] [CrossRef] [PubMed]
- Hay, M.; Thomas, D.W.; Craighead, J.L.; Economides, C.; Rosenthal, J. Clinical development success rates for investigational drugs. Nat. Biotechnol. 2014, 32, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Freskgård, P.O.; Urich, E. Antibody therapies in CNS diseases. Neuropharmacology 2017, 120, 38–55. [Google Scholar] [CrossRef] [PubMed]
- Carman, A.J.; Mills, J.H.; Krenz, A.; Kim, D.-G.; Bynoe, M.S. Adenosine Receptor Signaling Modulates Permeability of the Blood-Brain Barrier. J. Neurosci. 2011, 31, 13272–13280. [Google Scholar] [CrossRef]
- Rodriguez, A.; Tatter, S.B.; Debinski, W. Neurosurgical techniques for disruption of the blood–brain barrier for glioblastoma treatment. Pharmaceutics 2015, 7, 175–187. [Google Scholar] [CrossRef] [Green Version]
- Burgess, A.; Shah, K.; Hough, O.; Hynynen, K.; Burgess, A. Expert Review of Neurotherapeutics through the blood—Brain barrier Focused ultrasound-mediated drug delivery through the blood – brain barrier. Expert. Rev. Neurother. 2015, 17, 477–491. [Google Scholar] [CrossRef] [Green Version]
- Manich, G.; Cabezón, I.; del Valle, J.; Duran-Vilaregut, J.; Camins, A.; Pallàs, M.; Pelegrí, C.; Vilaplana, J. Study of the transcytosis of an anti-transferrin receptor antibody with a Fab′ cargo across the blood–brain barrier in mice. Eur. J. Pharm. Sci. 2013, 49, 556–564. [Google Scholar] [CrossRef]
- Fishman, J.B.; Rubin, J.B.; Handrahan, J.V.; Connor, J.R.; Fine, R.E. Receptor-mediated transcytosis of transferrin across the blood-brain barrier. J. Neurosci. Res. 1987, 18, 299–304. [Google Scholar] [CrossRef]
- Urayama, A.; Grubb, J.H.; Sly, W.S.; Banks, W.A. Mannose 6-Phosphate Receptor–mediated Transport of Sulfamidase Across the Blood–brain Barrier in the Newborn Mouse. Mol. Ther. 2008, 16, 1261–1266. [Google Scholar] [CrossRef]
- Gänger, S.; Schindowski, K. Tailoring formulations for intranasal Nose-to-Brain delivery via the olfactory area: A review on physico-chemical characteristics and mucociliary clearance of the nasal olfactory mucosa. Pharmaceutics 2018, 10, 116. [Google Scholar] [CrossRef] [Green Version]
- Mittal, D.; Ali, A.; Md, S.; Baboota, S.; Sahni, J.K.; Ali, J. Insights into direct nose to brain delivery: Current status and future perspective. Drug Deliv. 2014, 21, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Pardeshi, C.V.; Belgamwar, V.S. Direct nose to brain drug delivery via integrated nerve pathways bypassing the blood–brain barrier: An excellent platform for brain targeting. Expert Opin. Drug Deliv. 2013, 10, 957–972. [Google Scholar] [CrossRef] [PubMed]
- Ladel, S.; Flamm, J.; Zadeh, A.S.; Filzwieser, D.; Walter, J.C.; Schlossbauer, P.; Kinscherf, R.; Lischka, K.; Luksch, H.; Schindowski, K. Allogenic fc domain-facilitated uptake of IgG in nasal Lamina propria: Friend or foe for intranasal CNS delivery? Pharmaceutics 2018, 10, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stirling, C.M.A.; Charleston, B.; Takamatsu, H.; Claypool, S.; Lencer, W.; Blumberg, R.S.; Wileman, T.E. Characterization of the porcine neonatal Fc receptor—Potential use for trans-epithelial protein delivery. Immunology 2005, 114, 542–553. [Google Scholar] [CrossRef] [PubMed]
- Spiekermann, G.M.; Finn, P.W.; Ward, E.S.; Dumont, J.; Dickinson, B.L.; Blumberg, R.S.; Lencer, W.I. Receptor-mediated Immunoglobulin G Transport Across Mucosal Barriers in Adult Life. J. Exp. Med. 2002, 196, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Masuda, A.; Kuo, T.T.; Kobayashi, K.; Claypool, S.M.; Takagawa, T.; Kutsumi, H.; Azuma, T.; Lencer, W.I.; Blumberg, R.S. IgG transport across mucosal barriers by neonatal Fc receptor for IgG and mucosal immunity. Springer Semin. Immunopathol. 2006, 28, 397–403. [Google Scholar] [CrossRef]
- Roopenian, D.C.; Akilesh, S. FcRn: The neonatal Fc receptor comes of age. Nat. Rev. Immunol. 2007, 7, 715–725. [Google Scholar] [CrossRef]
- de Taeye, S.W.; Rispens, T.; Vidarsson, G. The Ligands for Human IgG and Their Effector Functions. Antibodies 2019, 8, 30. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Wang, W.; Li, S.; Jia, S.; Zhang, X.; Cao, T. Localization of neonatal Fc receptor for IgG in aggregated lymphoid nodules area in abomasum of Bactrian camels (Camelus bactrianus) of different ages. BMC Vet. Res. 2016, 237. [Google Scholar] [CrossRef] [Green Version]
- Xiao, G.; Gan, L.-S. Receptor-mediated endocytosis and brain delivery of therapeutic biologics. Int. J. Cell Biol. 2013, 2013, 703545. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, A. Immune Regulation of Antibody Access to Neuronal Tissues. Trends Mol. Med. 2017, 23, 227–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, A.; Balthasar, J.P. Investigation of the influence of FcRn on the distribution of IgG to the brain. AAPS J. 2009, 11, 553–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruano-salguero, J.S.; Lee, K.H. Antibody transcytosis across brain endothelial-like cells occurs nonspecifically and independent of FcRn. Sci. Rep. 2020, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilleminault, L.; Azzopardi, N.; Arnoult, C.; Sobilo, J.; Herve, V.; Montharu, J.; Diot, P. Fate of inhaled monoclonal antibodies after the deposition of aerosolized particles in the respiratory system. J. Control Release 2014, 196, 344–354. [Google Scholar] [CrossRef]
- Abuqayyas, L.; Balthasar, J.P. Investigation of the role of FcγR and FcRn in mAb distribution to the brain. Mol. Pharm. 2013, 10, 1505–1513. [Google Scholar] [CrossRef]
- Ishikawa, T.; Takizawa, T.; Iwaki, J.; Mishima, T.; Ui-Tei, K.; Takeshita, T.; Matsubara, S.; Takizawa, T. Fc gamma receptor IIb participates in maternal IgG trafficking of human placental endothelial cells. Int. J. Mol. Med. 2015, 35, 1273–1289. [Google Scholar] [CrossRef] [Green Version]
- Subedi, G.P.; Barb, A.W. The immunoglobulin G1 N-glycan composition affects binding to each low affinity Fc γ receptor. MAbs 2016, 8, 1512–1524. [Google Scholar] [CrossRef] [Green Version]
- Stewart, R.; Hammond, S.A.; Oberst, M.; Wilkinson, R.W. The role of Fc gamma receptors in the activity of immunomodulatory antibodies for cancer. J. Immunother. Cancer 2014, 2, 9. [Google Scholar] [CrossRef] [Green Version]
- Allhorn, M.; Olin, A.I.; Nimmerjahn, F.; Colin, M. Human IgG/FcγR interactions are modulated by streptococcal IgG glycan hydrolysis. PLoS ONE 2008, 3. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.M.; Wormald, M.R.; Rudd, P.M.; Davey, G.P. Fc gamma receptors: Glycobiology and therapeutic prospects. J. Inflamm. Res. 2016, 9, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Lux, A.; Yu, X.; Scanlan, C.N.; Nimmerjahn, F. Impact of Immune Complex Size and Glycosylation on IgG Binding to Human FcγRs. J. Immunol. 2013, 190, 4315–4323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cambay, F.; Raymond, C.; Brochu, D.; Gilbert, M.; Tu, T.M.; Cantin, C.; Lenferink, A.; Grail, M.; Henry, O.; De Crescenzo, G.; et al. Impact of IgG1 N-glycosylation on their interaction with Fc gamma receptors. Curr. Res. Immunol. 2020, 1, 23–37. [Google Scholar] [CrossRef]
- Franz-Montan, M.; Serpe, L.; Martinelli, C.C.M.; Da Silva, C.B.; dos Santos, C.P.; Novaes, P.D.; Volpato, M.C.; De Paula, E.; Lopez, R.F.V.; Groppo, F.C. Evaluation of different pig oral mucosa sites as permeability barrier models for drug permeation studies. Eur. J. Pharm. Sci. 2016, 81, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Tulinski, P.; Fluit, A.C.; van Putten, J.P.M.; de Bruin, A.; Glorieux, S.; Wagenaar, J.A.; Duim, B. An Ex Vivo Porcine Nasal Mucosa Explants Model to Study MRSA Colonization. PLoS ONE 2013, 8, e053783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egli, J.; Schlothauer, T.; Spick, C.; Seeber, S.; Singer, T.; Odermatt, A.; Iglesias, A. The Binding of Human IgG to Minipig FcγRs–Implications for Preclinical Assessment of Therapeutic Antibodies. Pharm. Res. 2019, 36, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladel, S.; Schlossbauer, P.; Flamm, J.; Luksch, H.; Mizaikoff, B.; Schindowski, K. Improved In Vitro Model for Intranasal Mucosal Drug Delivery: Primary Olfactory and Respiratory Epithelial Cells Compared with the Permanent Nasal Cell Line RPMI 2650. Pharmaceutics 2019, 11, 367. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.C.; Peter, H.; Lang, S.R.; Ditzinger, G.; Merkle, H.P. In vitro cell models to study nasal mucosal permeability and metabolism. Adv. Drug Deliv. Rev. 1998, 29, 51–79. [Google Scholar] [CrossRef]
- Pozzoli, M.; Sonvico, F.; Ong, H.X.; Traini, D.; Bebawy, M.; Young, P.M. Optimization of RPMI 2650 Cells as a Model for Nasal Mucosa. Respir. Drug Deliv. 2014, 2, 739–742. [Google Scholar]
- Mercier, C.; Hodin, S.; He, Z.; Perek, N.; Delavenne, X. Pharmacological Characterization of the RPMI 2650 Model as a Relevant Tool for Assessing the Permeability of Intranasal Drugs. Mol. Pharm. 2018, 15, 2246–2256. [Google Scholar] [CrossRef]
- Jadhav, K.; Gambhire, M.; Shaikh, I.; Kadam, V.; Pisal, S. Nasal Drug Delivery System-Factors Affecting and Applications. Curr. Drug Ther. 2008, 2, 27–38. [Google Scholar] [CrossRef]
- Collin, M.; Shannon, O.; Björck, L. IgG glycan hydrolysis by a bacterial enzyme as a therapy against autoimmune conditions. Proc. Natl. Acad. Sci. USA 2008, 105, 4265–4270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samson, G.; García De La Calera, A.; Dupuis-Girod, S.; Faure, F.; Decullier, E.; Paintaud, G.; Vignault, C.; Scoazec, J.Y.; Pivot, C.; Plauchu, H.; et al. Ex vivo study of bevacizumab transport through porcine nasal mucosa. Eur. J. Pharm. Biopharm. 2012, 80, 465–469. [Google Scholar] [CrossRef] [PubMed]
- de Courcey, F.; Zholos, A.V.; Atherton-Watson, H.; Williams, M.T.S.; Canning, P.; Danahay, H.L.; Elborn, J.S.; Ennis, M. Development of primary human nasal epithelial cell cultures for the study of cystic fibrosis pathophysiology. Am. J. Physiol. Physiol. 2012, 303, C1173–C1179. [Google Scholar] [CrossRef] [Green Version]
- Mackness, B.C.; Jaworski, J.A.; Boudanova, E.; Park, A.; Valente, D.; Mauriac, C.; Pasquier, O.; Schmidt, T.; Kabiri, M.; Kandira, A.; et al. Antibody Fc engineering for enhanced neonatal Fc receptor binding and prolonged circulation half-life. MAbs 2019, 11, 1276–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, M.; Olsén, A. Effect of SpeB and EndoS from Streptococcus pyogenes on human immunoglobulins. Infect. Immun. 2001, 69, 7187–7189. [Google Scholar] [CrossRef] [Green Version]
- Collin, M.; Björck, L. Toward Clinical use of the IgG Specifi c Enzymes IdeS and EndoS against Antibody-Mediated Diseases. Methods Mol. Biol. 2017, 1535, 357. [Google Scholar] [CrossRef]
- Reichl, S.; Becker, K. Cultivation of RPMI 2650 cells as an in-vitro model for human transmucosal nasal drug absorption studies: Optimization of selected culture conditions. J. Pharm. Pharmacol. 2012, 64, 1621–1630. [Google Scholar] [CrossRef]
- Wengst, A.; Reichl, S. RPMI 2650 epithelial model and three-dimensional reconstructed human nasal mucosa as in vitro models for nasal permeation studies. Eur. J. Pharm. Biopharm. 2010, 74, 290–297. [Google Scholar] [CrossRef]
- Mercier, C.; Perek, N.; Delavenne, X. Is RPMI 2650 a Suitable In Vitro Nasal Model for Drug Transport Studies? Eur. J. Drug Metab. Pharmacokinet. 2018, 43, 13–24. [Google Scholar] [CrossRef]
- Foss, S.; Grevys, A.; Sand, K.M.K.; Bern, M.; Blundell, P.; Michaelsen, T.E.; Pleass, R.J.; Sandlie, I.; Andersen, J.T. Enhanced FcRn-dependent transepithelial delivery of IgG by Fc-engineering and polymerization. J. Control Release 2016, 223, 42–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, R.; Matsui, M.; Kawasaki, N. Influence of N-glycosylation on effector functions and thermal stability of glycoengineered IgG1 monoclonal antibody with homogeneous glycoforms. MAbs 2019, 11, 350–372. [Google Scholar] [CrossRef] [PubMed]
- Sockolosky, J.T.; Szoka, F.C. The neonatal Fc receptor, FcRn, as a target for drug delivery and therapy. Adv. Drug Deliv. Rev. 2016, 91, 109–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, Q.M.; Barb, A.W. A perspective on the structure and receptor binding properties of immunoglobulin G Fc. Biochemistry 2015, 54, 2931–2942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, J.M.; Cosgrave, E.F.J.; Struwe, W.B.; Wormald, M.; Davey, G.P.; Jefferis, R.; Rudd, P.M. Fc Receptors: Humanized Mice to Study FcγR Function; SpringerLink: New York, NY, USA, 2014; Volume 382, ISBN 978-3-319-07910-3. [Google Scholar]
- Pyzik, M.; Sand, K.M.K.; Hubbard, J.J.; Andersen, J.T.; Sandlie, I.; Blumberg, R.S. The Neonatal Fc Receptor (FcRn): A Misnomer? Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Golebski, K.; Hoepel, W.; van Egmond, D.; de Groot, E.J.; Amatngalim, G.D.; Beekman, J.M.; Fokkens, W.J.; van Drunen, C.M.; den Dunnen, J. FcγRIII stimulation breaks the tolerance of human nasal epithelial cells to bacteria through cross-talk with TLR4. Mucosal Immunol. 2019, 12, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Nimmerjahn, F.; Ravetch, J.V. Fcγ receptors: Old friends and new family members. Immunity 2006, 24, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Zeng, R.; Bai, Y.; Roopenian, D.C.; Zhu, X. Efficient mucosal vaccination mediated by the neonatal Fc receptor. Nat. Biotechnol. 2011, 29, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Tzaban, S.; Massol, R.H.; Yen, E.; Hamman, W.; Frank, S.R.; Lapierre, L.A.; Hansen, S.H.; Goldenring, J.R.; Blumberg, R.S.; Lencer, W.I. The recycling and transcytotic pathways for IgG transport by FcRn are distinct and display an inherent polarity. J. Cell Biol. 2009, 185, 673–684. [Google Scholar] [CrossRef] [Green Version]
- Schlachetzki, F.; Zhu, C.; Pardridge, W.M. Expression of the neonatal Fc receptor (FcRn) at the blood-brain barrier. J. Neurochem. 2002, 81, 203–206. [Google Scholar] [CrossRef]
- Stapleton, N.M.; Brinkhaus, M.; Armour, K.L.; Bentlage, A.E.H.; de Taeye, S.W.; Temming, A.R.; Mok, J.Y.; Brasser, G.; Maas, M.; van Esch, W.J.E.; et al. Reduced FcRn-mediated transcytosis of IgG2 due to a missing Glycine in its lower hinge. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Schneider, Z.; Jani, P.K.; Szikora, B.; Végh, A.; Kövesdi, D.; Iliás, A.; Cervenak, J.; Balogh, P.; Kurucz, I.; Kacskovics, I. Overexpression of bovine FcRn in mice enhances T-dependent immune responses by amplifying T helper cell frequency and germinal center enlargement in the spleen. Front. Immunol. 2015, 6, 357. [Google Scholar] [CrossRef] [PubMed]
- Claypool, S.M.; Dickinson, B.L.; Wagner, J.S.; Johansen, F.; Venu, N.; Borawski, J.A.; Lencer, W.I.; Blumberg, R.S. Bidirectional Transepithelial IgG Transport by a Strongly Polarized Basolateral Membrane Fcγ-Receptor. Mol. Biol. Cell 2004, 15, 1746–1759. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Pardridge, W.M. Mediated efflux of IgG molecules from brain to blood across the blood–brain barrier. J. Neuroimmunol. 2001, 114, 168–172. [Google Scholar] [CrossRef]
- Dickinson, B.L.; Blumberg, R.S.; Wayne, I.; Invest, J.C.; Dickinson, B.L.; Badizadegan, K.; Wu, Z.; Ahouse, J.C.; Zhu, X.; Simister, N.E.; et al. Bidirectional FcRn-dependent IgG transport in a polarized human intestinal epithelial cell line. J. Clin. Investig. 1999, 104, 903–911. [Google Scholar] [CrossRef] [Green Version]
- Neuber, T.; Frese, K.; Jaehrling, J.; Jäger, S.; Daubert, D.; Felderer, K.; Linnemann, M.; Höhne, A.; Kaden, S.; Kölln, J.; et al. Characterization and screening of IgG binding to the neonatal Fc receptor. MAbs 2014, 6, 928–942. [Google Scholar] [CrossRef] [Green Version]
- Kacskovics, I. Fc receptors in livestock species. Vet. Immunol. Immunopathol. 2004, 102, 351–362. [Google Scholar] [CrossRef]
- Ober, R.J.; Radu, C.G.; Ghetie, V.; Ward, E.S. Differences in promiscuity for antibody–FcRn interactions across species: Implications for therapeutic antibodies. Int. Immunol. 2001, 13, 1551–1559. [Google Scholar] [CrossRef] [Green Version]
- Martins, J.P.; Kennedy, P.J.; Santos, H.A.; Barrias, C.; Sarmento, B. A comprehensive review of the neonatal Fc receptor and its application in drug delivery. Pharmacol. Ther. 2016, 161, 22–39. [Google Scholar] [CrossRef]
- England, R.J.; Homer, J.J.; Knight, L.C.; Ell, S.R. Nasal pH measurement: A reliable and repeatable parameter. Clin. Otolaryngol. Allied Sci. 1999, 24, 67–68. [Google Scholar] [CrossRef] [Green Version]
- Hornby, P.J.; Cooper, P.R.; Kliwinski, C.; Ragwan, E.; Mabus, J.R.; Harman, B.; Thompson, S.; Kauffman, A.L.; Yan, Z.; Tam, S.H.; et al. Human and non-human primate intestinal FcRn expression and immunoglobulin G transcytosis. Pharm. Res. 2014, 31, 908–922. [Google Scholar] [CrossRef] [Green Version]
- Cianga, C.; Cianga, P.; Plamadeala, P.; Amalinei, C. Nonclassical major histocompatibility complex I-like Fc neonatal receptor (FcRn) expression in neonatal human tissues. Hum. Immunol. 2011, 72, 1176–1187. [Google Scholar] [CrossRef] [PubMed]
- Debertin, A.S.; Tschernig, T.; Tönjes, H.; Kleemann, W.J.; Tröger, H.D.; Pabst, R. Nasal-associated lymphoid tissue (NALT): Frequency and localization in young children. Clin. Exp. Immunol. 2003, 134, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Debertin, A.S.; Tschernig, T.; Schürmann, A.; Bajanowski, T.; Brinkmann, B.; Pabst, R. Coincidence of different structures of mucosa-associated lymphoid tissue (MALT) in the respiratory tract of children: No indications for enhanced mucosal immunostimulation in sudden infant death syndrome (SIDS). Clin. Exp. Immunol. 2006, 146, 54–59. [Google Scholar] [CrossRef]
- Brandtzaeg, P.; Pabst, R. Let’s go mucosal: Communication on slippery ground. Trends Immunol. 2004, 25, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A. Mucosal Dendritic Cells. Annu. Rev. Immunol. 2007, 25, 381–418. [Google Scholar] [CrossRef]
- Kang, H.; Yan, M.; Yu, Q.; Yang, Q. Characteristics of nasal-associated lymphoid tissue (NALT) and nasal absorption capacity in chicken. PLoS ONE 2013, 8, e084097. [Google Scholar] [CrossRef]
- Li, X.; Ptacek, T.S.; Brown, E.E.; Edberg, J.C. Fcγ receptors: Structure, function and role as genetic risk factors in SLE. Genes Immun. 2009, 10, 380–389. [Google Scholar] [CrossRef] [Green Version]
- Gitlin, D.; Koch, C. On the mechanisms of maternofetal transfer of human albumin and gamma-G globulin in the mouse. J. Clin. Investig. 1968, 47, 1204–1209. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, B.; Hill, M.; Reynaud, L.; Hey, A.; Barrow, P. FcRn Expression on Placenta and Fetal Jejunum during Early, Mid-, and Late Gestation in Minipigs. Toxicol. Pathol. 2016, 44, 486–491. [Google Scholar] [CrossRef] [Green Version]
- De Fraissinette, A.; Brun, R.; Felix, H.; Vonderscher, J.; Rummelt, A. Evaluation of the human cell line RPMI 2650 as an in vitro nasal model. Rhinology 1995, 33, 194–198. [Google Scholar]
- O’Connell, L.Y.; Lai, J.; Presta, L.G.; Keck, R.; Meng, Y.G.; Hong, K.; Weikert, S.H.A.; Shields, R.L. Lack of Fucose on Human IgG1 N -Linked Oligosaccharide Improves Binding to Human FcγRIII and Antibody-dependent Cellular Toxicity. J. Biol. Chem. 2002, 277, 26733–26740. [Google Scholar] [CrossRef] [Green Version]
- Mathiesen, L.; Nielsen, L.K.; Andersen, J.T.; Grevys, A.; Sandlie, I.; Michaelsen, T.E.; Hedegaard, M.; Knudsen, L.E.; Dziegiel, M.H. Maternofetal transplacental transport of recombinant IgG antibodies lacking effector functions. Blood 2013, 122, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Bakchoul, T.; Walek, K.; Krautwurst, A.; Rummel, M.; Bein, G.; Santoso, S.; Sachs, U.J. Glycosylation of autoantibodies: Insights into the mechanisms of immune thrombocytopenia. Thromb. Haemost. 2013, 110, 1259–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stapleton, N.M.; Armstrong-Fisher, S.S.; Andersen, J.T.; van der Schoot, C.E.; Porter, C.; Page, K.R.; Falconer, D.; de Haas, M.; Williamson, L.M.; Clark, M.R.; et al. Human IgG lacking effector functions demonstrate lower FcRn-binding and reduced transplacental transport. Mol. Immunol. 2018, 95, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Malek, A. Ex vivo human placenta models: Transport of immunoglobulin G and its subclasses. Vaccine 2003, 21, 3362–3364. [Google Scholar] [CrossRef]
- Stapleton, N.M.; Andersen, J.T.; Stemerding, A.M.; Bjarnarson, S.P.; Verheul, R.C.; Gerritsen, J.; Zhao, Y.; Kleijer, M.; Sandlie, I.; de Haas, M.; et al. Competition for FcRn-mediated transport gives rise to short half-life of human IgG3 and offers therapeutic potential. Nat. Commun. 2011, 2, 599. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Antigen | Host | Source, Catalog Number |
|---|---|---|---|
| Anti-rabbit IgG-Rhodamine RedTM-X | Whole molecule rabbit IgG | Donkey | Jackson Immuno Research Europe Ltd., Cambridgeshire, UK, Cat. #711-295-152 |
| Anti-murine IgG-Alexa Fluor® 488 | Whole molecule mouse IgG | Goat | Jackson Immuno Research Europe Ltd., Ely, UK, Cat. #115-545-003 |
| AffiniPure Anti swine IgG (H+L)-Alexa Fluor® 647 | Whole molecule porcine IgG | Goat | Jackson Immuno Research Europe Ltd., Ely, UK, Cat. #114-605-003 |
| AffiniPure Anti human-Alexa Fluor® 647 | Whole molecule human IgG | Donkey | Jackson Immuno Research Europe Ltd., Ely, UK, Cat. #709-605-149 |
| Anti-rabbit IgG-HRP | Whole molecule rabbit IgG | Goat | Jackson Immuno Research Europe Ltd., Ely, UK, Cat. #111-035-003 |
| Anti-murine IgG-HRP | Whole molecule rabbit IgG | Goat | Sigma Aldrich, Taufkirchen, Germany, Cat. #AP5278 |
| Anti-human κ light chain IgG-HRP | Human κ light chain | Goat | SouthernBiotech, Birmingham, AL, USA, Cat. #2060-05 |
| AffiniPure Anti swine IgG (H+L)-HRP | Whole molecule porcine IgG | Goat | Jackson Immuno Research Europe Ltd., Ely, UK, Cat. #114-035-003 |
| AffiniPure Anti swine IgG (H+L) | Whole molecule porcine IgG | Goat | Jackson Immuno Research Europe Ltd., Ely, UK, Cat. #114-005-003 |
| Fc Receptor | Sensitivity to Deglycosylated IgG | Potential Relevance in Epithelial IgG Trafficking | Refs. |
|---|---|---|---|
| FCGR1 | Rather insensitive due alternative binding domain. | Has not yet been implicated in IgG trafficking; expression in epithelial has not yet been demonstrated. | [30,32,53] |
| FCGR2 | Endo S treatment of IgG decreases their interaction with FCGR2, but fucose does not alter the affinity to FCGR2. | Expression in epithelial cells from placenta was shown in several studies. Was linked to IgG trafficking of maternal IgG to the fetus in placental cell lines (FCGR2b). | [26,29,32,53,54] |
| FCGR3a | Endo S treatment abolishes the interaction with FCGR3, but also fucosylated IgGs such as hIgG used here were shown to bind with significantly decreased affinity. | Expression in human nasal epithelium was demonstrated. Controversially discussed for placental IgG trafficking. | [32,53,55,56] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ladel, S.; Maigler, F.; Flamm, J.; Schlossbauer, P.; Handl, A.; Hermann, R.; Herzog, H.; Hummel, T.; Mizaikoff, B.; Schindowski, K. Impact of Glycosylation and Species Origin on the Uptake and Permeation of IgGs through the Nasal Airway Mucosa. Pharmaceutics 2020, 12, 1014. https://doi.org/10.3390/pharmaceutics12111014
Ladel S, Maigler F, Flamm J, Schlossbauer P, Handl A, Hermann R, Herzog H, Hummel T, Mizaikoff B, Schindowski K. Impact of Glycosylation and Species Origin on the Uptake and Permeation of IgGs through the Nasal Airway Mucosa. Pharmaceutics. 2020; 12(11):1014. https://doi.org/10.3390/pharmaceutics12111014
Chicago/Turabian StyleLadel, Simone, Frank Maigler, Johannes Flamm, Patrick Schlossbauer, Alina Handl, Rebecca Hermann, Helena Herzog, Thomas Hummel, Boris Mizaikoff, and Katharina Schindowski. 2020. "Impact of Glycosylation and Species Origin on the Uptake and Permeation of IgGs through the Nasal Airway Mucosa" Pharmaceutics 12, no. 11: 1014. https://doi.org/10.3390/pharmaceutics12111014
APA StyleLadel, S., Maigler, F., Flamm, J., Schlossbauer, P., Handl, A., Hermann, R., Herzog, H., Hummel, T., Mizaikoff, B., & Schindowski, K. (2020). Impact of Glycosylation and Species Origin on the Uptake and Permeation of IgGs through the Nasal Airway Mucosa. Pharmaceutics, 12(11), 1014. https://doi.org/10.3390/pharmaceutics12111014