Targeting Small Molecule Delivery to the Brain and Spinal Cord via Intranasal Administration of Rabies Virus Glycoprotein (RVG29)-Modified PLGA Nanoparticles

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Nanoparticle Preparation
2.3. Nanoparticle Characterization
2.4. Nanoparticle Administration
2.5. Tissue Homogenization
2.6. DiR Imaging
2.7. Data Analysis
3. Results
3.1. Nanoparticle Characterization
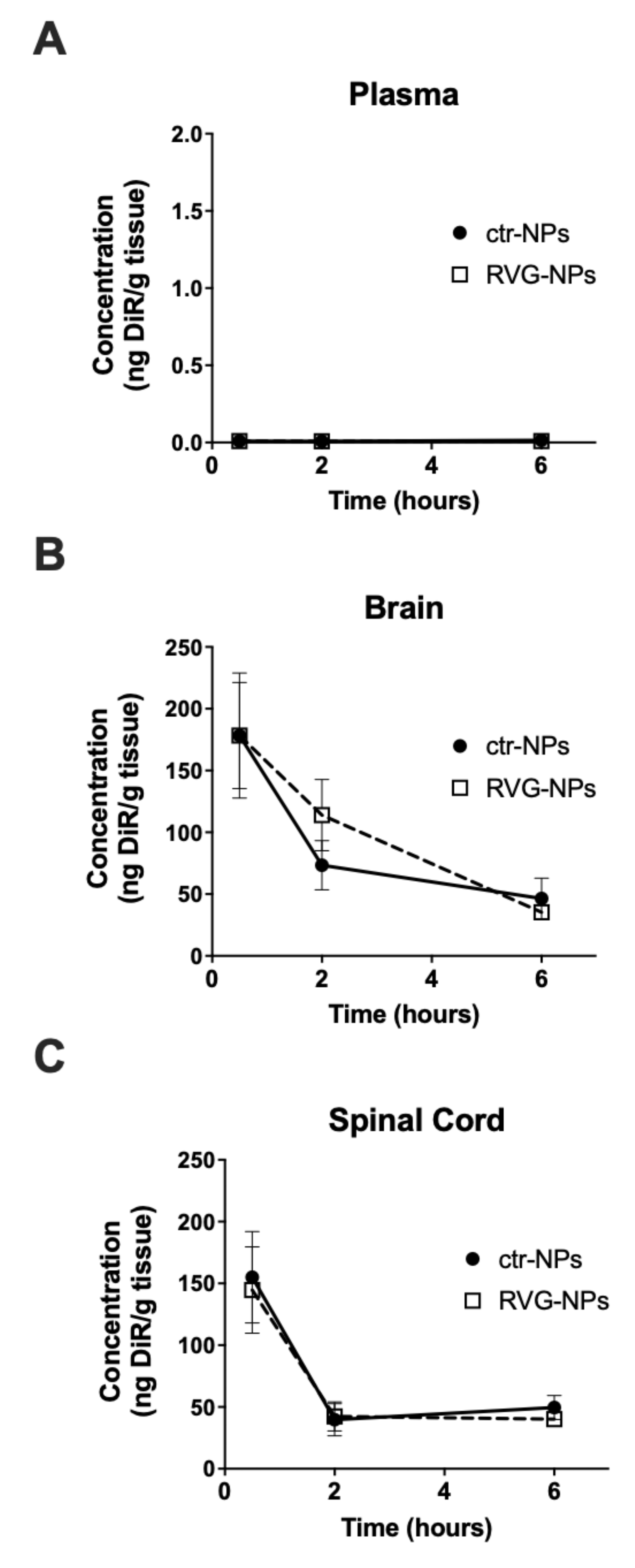
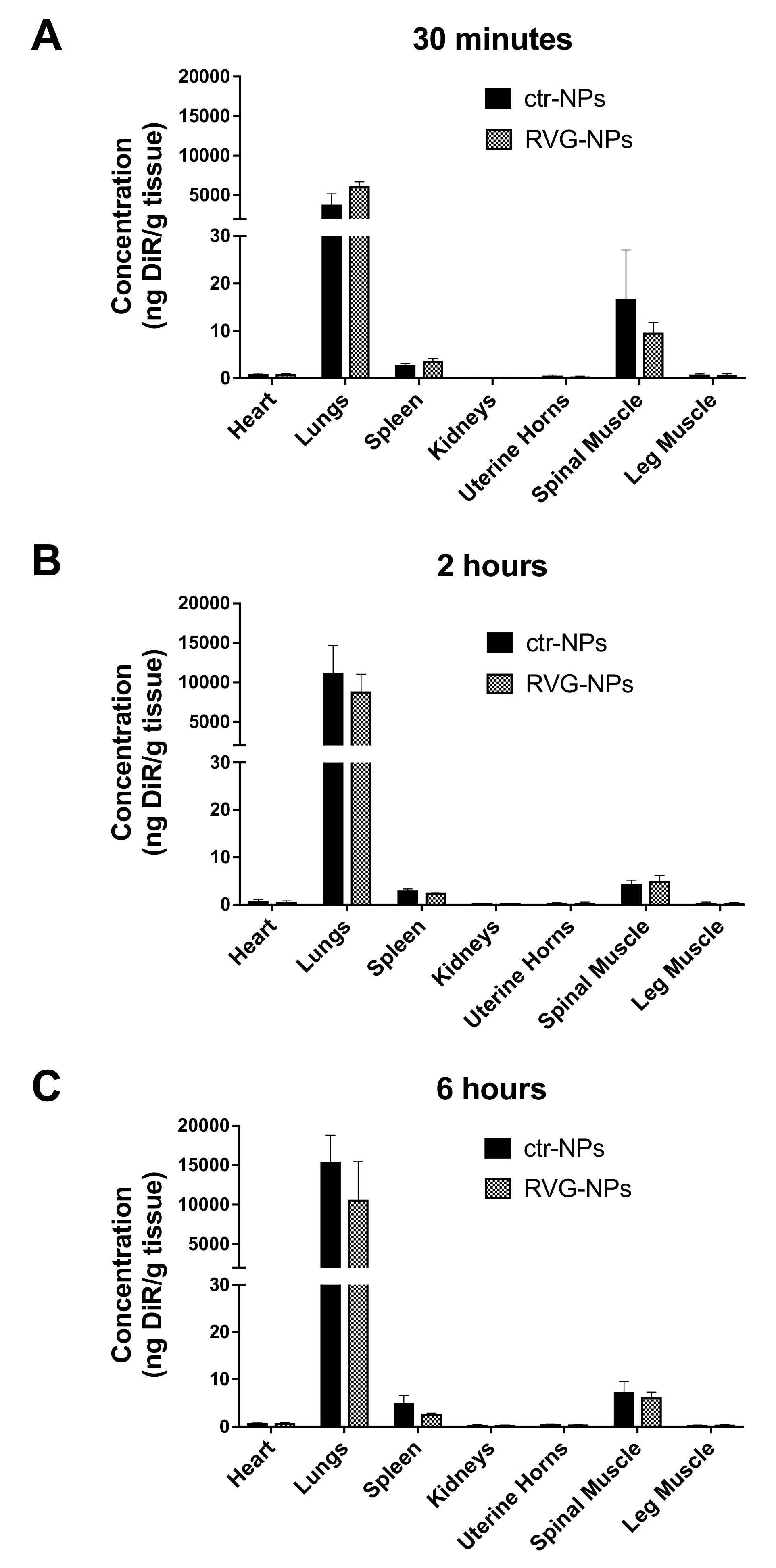
3.2. Whole-Organ Biodistribution
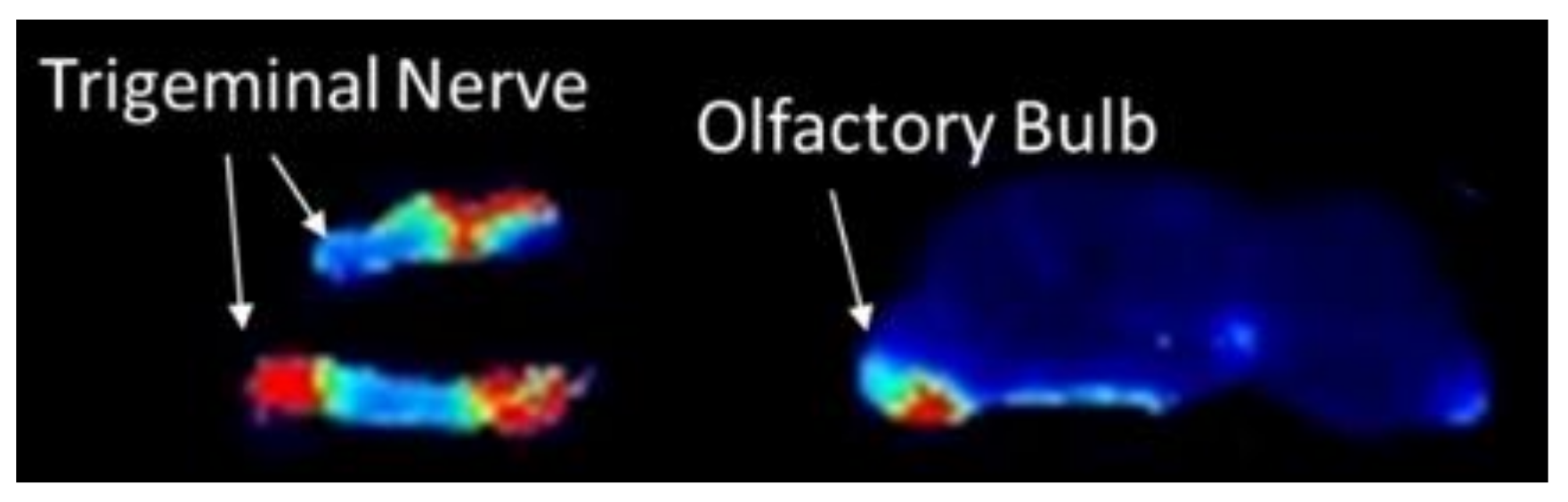
3.3. Spatial Biodistribution Within the CNS
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, X.-Q.; Fawcett, J.R.; Rahman, Y.-E.; Ala, T.A.; Frey, W.H., II. Delivery of nerve growth factor to the brain via the olfactory pathway. J. Alzheimers Dis. 1998, 1, 35–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorne, R.G.; Pronk, G.J.; Padmanabhan, V.; Frey, W.H., II. Delivery of insulin-like growth factor-I to the rat brain and spinal cord along olfactory and trigeminal pathways following intranasal administration. Neuroscience 2004, 127, 481–496. [Google Scholar] [CrossRef] [PubMed]
- Ross, T.M.; Martinez, P.M.; Renner, J.C.; Thorne, R.G.; Hanson, L.R.; Frey, W.H. Intranasal administration of interferon beta bypasses the blood-brain barrier to target the central nervous system and cervical lymph nodes: A non-invasive treatment strategy for multiple sclerosis. J. Neuroimmunol. 2004, 151, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.-P.; Xu, Q.-Q.; Zhang, Q.; Zhang, W.-P.; Zhang, L.-H.; Wei, E.-Q. Intranasal recombinant human erythropoietin protects rats against focal cerebral ischemia. Neurosci. Lett. 2005, 387, 5–10. [Google Scholar] [CrossRef]
- Han, I.-K.; Kim, M.Y.; Byun, H.-M.; Hwang, T.S.; Kim, J.M.; Hwang, K.W.; Park, T.G.; Jung, W.W.; Chun, T.; Jeong, G.J.; et al. Enhanced brain targeting efficiency of intranasally administered plasmid DNA: An alternative route for brain gene therapy. J. Mol. Med. Berl. Ger. 2007, 85, 75–83. [Google Scholar] [CrossRef]
- Wang, D.; Gao, Y.; Yun, L. Study on brain targeting of raltitrexed following intranasal administration in rats. Cancer Chemother. Pharmacol. 2006, 57, 97–104. [Google Scholar] [CrossRef]
- Barakat, N.S.; Omar, S.A.; Ahmed, A. Carbamazepine uptake into rat brain following intra-olfactory transport. J. Pharm. Pharmacol. 2006, 58, 63–72. [Google Scholar] [CrossRef]
- Upadhyay, S.; Parikh, A.; Joshi, P.; Upadhyay, U.M. Intranasal drug delivery system-A glimpse to become maestro. J. Appl. Pharm. Sci. 2011, 11, 34–44. [Google Scholar]
- Lochhead, J.J.; Thorne, R.G. Intranasal delivery of biologics to the central nervous system. Adv. Drug Deliv. Rev. 2012, 64, 614–628. [Google Scholar] [CrossRef]
- Ghori, M.U.; Mahdi, M.H.; Smith, A.M.; Conway, B.R. Nasal drug delivery systems: An overview. Am. J. Pharm. Sci. 2015, 3, 110–119. [Google Scholar]
- Seju, U.; Kumar, A.; Sawant, K.K. Development and evaluation of olanzapine-loaded PLGA nanoparticles for nose-to-brain delivery: In vitro and in vivo studies. Acta Biomater. 2011, 7, 4169–4176. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Sharma, R.K.; Sharma, N.; Gabrani, R.; Sharma, S.K.; Ali, J.; Dang, S. Nose-to-brain delivery of PLGA-diazepam nanoparticles. AAPS PharmSciTech 2015, 16, 1108–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vyas, S.P.; Sihorkar, V. Endogenous carriers and ligands in non-immunogenic site-specific drug delivery. Adv. Drug Deliv. Rev. 2000, 43, 101–164. [Google Scholar] [CrossRef]
- Huey, R.; Hawthorne, S.; McCarron, P. The potential use of rabies virus glycoprotein-derived peptides to facilitate drug delivery into the central nervous system: A mini review. J. Drug Target. 2017, 25, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Cook, R.L.; Householder, K.T.; Chung, E.P.; Prakapenka, A.V.; DiPerna, D.M.; Sirianni, R.W. A critical evaluation of drug delivery from ligand modified nanoparticles: Confounding small molecule distribution and efficacy in the central nervous system. J. Control. Release 2015, 220, 89–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storer, R.J.; Akerman, S.; Goadsby, P.J. GABA receptors modulate trigeminovascular nociceptive neurotransmission in the trigeminocervical complex. Br. J. Pharmacol. 2001, 134, 896–904. [Google Scholar] [CrossRef] [Green Version]
- Henry, M.A.; Westrum, L.E.; Bothwell, M.; Johnson, L.R. Nerve growth factor receptor (p75)-immunoreactivity in the normal adult feline trigeminal system and following retrogasserian rhizotomy. J. Comp. Neurol. 1993, 335, 425–436. [Google Scholar] [CrossRef]
- Alimohammadi, H.; Silver, W.L. Evidence for nicotinic acetylcholine receptors on nasal trigeminal nerve endings of the rat. Chem. Senses 2000, 25, 61–66. [Google Scholar] [CrossRef]
- Pardeshi, C.V.; Belgamwar, V.S. Direct nose to brain drug delivery via integrated nerve pathways bypassing the blood-brain barrier: An excellent platform for brain targeting. Expert Opin. Drug Deliv. 2013, 10, 957–972. [Google Scholar] [CrossRef]
- McCall, R.L.; Sirianni, R.W. PLGA nanoparticles formed by single- or double-emulsion with vitamin E-TPGS. J. Vis. Exp. JoVE 2013, 82, 51015. [Google Scholar] [CrossRef] [Green Version]
- Sirianni, R.W.; Zheng, M.Q.; Patel, T.R.; Shafbauer, T.; Zhou, J.; Saltzman, W.M.; Carson, R.E.; Huang, Y. Radiolabeling of poly(lactic-co-glycolic acid) (PLGA) nanoparticles with biotinylated F-18 prosthetic groups and imaging of their delivery to the brain with positron emission tomography. Bioconj. Chem. 2014, 25, 2157–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Southam, D.; Dolovich, M.; O’Byrne, P.; Inman, M. Distribution of intranasal instillations in mice: Effects of volume, time, body position, and anesthesia. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 282, L833–L839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiPerna, D.M.; Prakapenka, A.V.; Chung, E.P.; Sirianni, R.W. Non-enzymatic tissue homogenization for biodistribution analysis. Methods Mol. Biol. 2018, 1830, 191–199. [Google Scholar]
- Lü, J.M.; Wang, X.; Marin-Muller, C.; Wang, H.; Lin, P.H.; Yao, Q.; Chen, C. Current advances in research and clinical applications of PLGA-based nanotechnology. Expert Rev. Mol. Diagn. 2009, 9, 325–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonvico, F.; Clementino, A.; Buttini, F.; Colombo, G.; Pescina, S.; Stanisçuaski Guterres, S.; Raffin Pohlmann, A.; Nicoli, S. Surface-modified nanocarriers for nose-to-brain delivery: From bioadhesion to targeting. Pharmaceutics 2018, 10, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Wu, B.; Zhang, Q.; Chen, J.; Zhu, J.; Zhang, W.; Rong, Z.; Chen, H.; Jiang, X. Brain delivery of vasoactive intestinal peptide enhanced with the nanoparticles conjugated with wheat germ agglutinin following intranasal administration. J. Control. Release 2007, 121, 156–167. [Google Scholar] [CrossRef]
- Hoekman, J.D.; Srivastava, P.; Ho, R.J. Aerosol-stable peptide-coated liposome nanoparticles: A proof-of-concept study with opioid fentanyl in enhancing analgesic effects and reducing plasma drug exposure. J. Pharm. Sci. 2014, 103, 2231–2239. [Google Scholar] [CrossRef] [Green Version]
- Bi, C.; Wang, A.; Chu, Y.; Liu, S.; Mu, H.; Liu, W.; Wu, Z.; Sun, K.; Li, Y. Intranasal delivery of rotigotine to the brain with lactoferrin-modified PEG-PLGA nanoparticles for Parkinson’s disease treatment. Int. J. Nanomed. 2016, 11, 6547–6559. [Google Scholar] [CrossRef] [Green Version]
- Taki, H.; Kanazawa, T.; Akiyama, F.; Takashima, Y.; Okada, H. Intranasal delivery of camptothecin-loaded tat-modified nanomicells for treatment of intracranial brain tumors. Pharmaceuticals 2012, 15, 1092–1102. [Google Scholar] [CrossRef]
- Householder, K.T.; Dharmaraj, S.; Sandberg, D.I.; Wechsler-Reya, R.J.; Sirianni, R.W. Fate of nanoparticles in the central nervous system after intrathecal injection in healthy mice. Sci. Rep. 2019, 9, 12587. [Google Scholar] [CrossRef] [Green Version]
- Johnson, N.J.; Hanson, L.R.; Frey, W.H. Trigeminal pathways delivery a low molecular weight drug from the nose to the brain and orofacial structures. Mol. Pharm. 2010, 7, 884–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittal, D.; Ali, A.; Baboota, S.; Sahni, J.K.; Ali, J. Insights into direct nose to brain delivery: Current status and future perspective. Drug Deliv. 2014, 21, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Honig, M.G.; Hume, R.I. Fluorescent carbocyanine dyes allow living neurons of identified origin to be studied in long-term cultures. J. Cell Biol. 1986, 103, 171–187. [Google Scholar] [CrossRef] [PubMed]
- Dogru, S.; Van Daele, D.; Hansen, M.R. Retrograde labeling of the rat facial nerve with carbocyanine dyes to enhance intraoperative identification. Ann. Otol. Rhinol. Laryngol. 2008, 117, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Hadaczek, P.; Yamashita, Y.; Mirek, H.; Tamas, L.; Bohn, M.C.; Noble, C.; Park, J.W.; Bankiewicz, K. The “perivascular pump“ driven by arterial pulsation is a powerful mechanism for the distribution of therapeutic molecules within the brain. Mol. Ther. 2006, 14, 69–78. [Google Scholar] [CrossRef]
- Djupesland, P.G.; Messina, J.C.; Mahmoud, R.A. The nasal approach to delivering treatment for brain diseases: An anatomic, physiologic, and delivery technology overview. Ther. Deliv. 2014, 5, 709–733. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Sanz, M.; Villegas-Pérez, M.P.; Bray, G.M.; Aguayo, A.J. Persistent retrograde labeling of adult rat retinal ganglion cells with the carbocyanine dye dil. Exp. Neurol. 1988, 102, 92–101. [Google Scholar] [CrossRef]
- Phukan, K.; Nandy, M.; Sharma, R.B.; Sharma, H.K. Nanosized drug delivery systems for direct nose to brain targeting: A review. Recent Pat. Drug Deliv. Formul. 2016, 10, 156–164. [Google Scholar] [CrossRef]
- Lafon, M. Rabies virus receptors. J. Neurovirol. 2005, 11, 82–87. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, R.; Han, L.; Ke, W.; Shao, K.; Ye, L.; Lou, J.; Jiang, C. Brain-targeting gene delivery and cellular internalization mechanisms for modified rabies virus glycoprotein RVG29 nanoparticles. Biomaterials 2009, 30, 4195–4202. [Google Scholar] [CrossRef]
- Lafay, F.; Coulon, P.; Astic, L.; Saucier, D.; Riche, D.; Holley, A.; Flamand, A. Spread of the CVS strain of rabies virus and of the avirulent mutant AvO1 along the olfactory pathways of the mouse after intranasal inoculation. Virology 1991, 183, 320–330. [Google Scholar] [CrossRef]
- Babic, N.; Mettenleiter, T.C.; Ugolini, G.; Flamand, A.; Coulon, P. Propagation of pseudorabies virus in the nervous system of the mouse after intranasal inoculation. Virology 1994, 204, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Rassu, G.; Soddu, E.; Posadino, A.M.; Pintus, G.; Sarmento, B.; Giunchedi, P.; Gavini, E. Nose-to-brain delivery of BACE1 siRNA loaded in solid lipid nanoparticles for Alzheimer’s therapy. Colloids Surf. B Biointerfaces 2017, 152, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Medina, D.X.; Householder, K.T.; Ceton, R.; Kovalik, T.; Heffernan, J.M.; Shankar, R.V.; Bowser, R.P.; Wechsler-Reya, R.J.; Sirianni, R.W. Optical barcoding of PLGA for multispectral analysis of nanoparticle fate in vivo. J. Control. Release 2017, 253, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, C.; Zong, S.; Qi, J.; Dong, X.; Zhao, W.; Wu, W.; Fu, Q.; Lu, Y.; Chen, Z. The trigeminal pathway dominates the nose-to-brain transportation of intact polymeric nanoparticles: Evidence from aggregation-caused quenching probes. J. Biomed. Nanotechnol. 2019, 15, 686–702. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, E.; Feng, Y.; Qi, J.; Fan, W.; Ma, Y.; He, H.; Xia, F.; Dong, X.; Zhao, W.; Lu, Y.; et al. Evidence of nose-to-brain delivery of nanoemulsions: Cargoes but not vehicles. Nanoscale 2017, 9, 1174–1183. [Google Scholar] [CrossRef]
- Lai, S.K.; Wang, Y.Y.; Hanes, J. Mucus-penetrating nanoparticles for drug and gene delivery to mucosal tissues. Adv. Drug Deliv. Rev. 2009, 61, 158–171. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | SEM | DLS | |||||
|---|---|---|---|---|---|---|---|
| Loading (%) | EE1 (%) | Size (nm) | PD2 (nm) | Size (nm) | PD (nm) | Zeta Potential (mV) | |
| RVG-NPs | 0.26 | 38.1 | 129 | 36 | 188 | 44 | 0.36 ± 1.76 |
| NPs | - | - | 141 | 31 | 238 | 56 | 1.69 ± 0.95 |
| Time | CNS Region | Ctr-NPs1 | RVG-NPs1 | Targeting2 |
|---|---|---|---|---|
| 0.5 h | Brain | |||
| Olfactory Bulb | 627.8 ± 507.6 | 325.7 ± 189.8 | −48% | |
| Cortex | 120.7 ± 95.5 | 95.5 ± 93.3 | −21% | |
| Striatum | 241.8 ±130.9 | 234.4 ± 217.1 | −3% | |
| Midbrain | 260.3 ± 98.4 | 271.2 ± 135.6 | +4% | |
| Hippocampus | 148.5 ± 115.6 | 164.3 ± 127.7 | +11% | |
| Cerebellum | 234.4 ± 78.6 | 223.8 ± 128.1 | −5% | |
| Brain Stem | 205.6 ± 89.5 | 235.7 ± 174.6 | +15% | |
| Spinal Cord3 | ||||
| SC-C | 278.1 ± 98.6 | 263.7 ± 155.3 | −5% | |
| SC-T | 122.1 ± 76.4 | 111.8 ± 77.2 | −8% | |
| SC-L | 85.2 ± 72.4 | 99.2 ± 74.4 | +16% | |
| SC-S | 126.8 ± 107.3 | 121.7 ± 87.7 | −4% | |
| 2 h | Brain | |||
| Olfactory Bulb | 227.8 ± 257.1 | 359.9 ± 244.4 | +58% | |
| Cortex | 73.7 ± 101.4 | 46.3 ± 25.9 | −37% | |
| Striatum* | 43.4 ± 19.4 | 160.4 ± 93.6 | +270% | |
| Midbrain* | 51.2 ± 20.1 | 154.7 ± 108.8 | +202% | |
| Hippocampus | 41.5 ± 21.9 | 87.4 ± 83.6 | +111% | |
| Cerebellum | 68.4 ± 79.6 | 108.9 ± 84.5 | +59% | |
| Brain Stem* | 70.0 ± 40.8 | 186.1 ± 139.7 | +166% | |
| Spinal Cord | ||||
| SC-C | 40.1 ± 35.7 | 114.3 ± 106.3 | +185% | |
| SC-T | 40.7 ± 20.4 | 42.5 ± 12.6 | +5% | |
| SC-L | 38.2 ± 29.1 | 39.0 ± 3.7 | +2% | |
| SC-S | 69.2 ± 107.6 | 29.8 ± 10.0 | −57% | |
| 6 h | Brain | |||
| Olfactory Bulb | 133.5 ± 222.6 | 63.0 ± 27.7 | −53% | |
| Cortex | 33.8 ± 26.9 | 27.6 ± 10.5 | −18% | |
| Striatum | 51.6 ± 40.0 | 38.1 ± 10.5 | −26% | |
| Midbrain | 50.6 ± 37.2 | 39.3 ± 27.8 | −22% | |
| Hippocampus | 41.1 ± 35.3 | 28.2 ± 9.6 | −31% | |
| Cerebellum | 47.3 ± 37.8 | 32.0 ± 11.4 | −32% | |
| Brain Stem | 46.2 ± 26.3 | 51.8 ± 29.7 | +12% | |
| Spinal Cord | ||||
| SC-C | 96.2 ± 74.1 | 70.7 ± 31.8 | −26% | |
| SC-T | 37.6 ± 9.1 | 31.0 ± 10.2 | −18% | |
| SC-L | 30.2 ± 17.4 | 27.8 ± 9.4 | −8% | |
| SC-S | 37.9 ± 8.8 | 43.3 ± 22.5 | +14% | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chung, E.P.; Cotter, J.D.; Prakapenka, A.V.; Cook, R.L.; DiPerna, D.M.; Sirianni, R.W. Targeting Small Molecule Delivery to the Brain and Spinal Cord via Intranasal Administration of Rabies Virus Glycoprotein (RVG29)-Modified PLGA Nanoparticles. Pharmaceutics 2020, 12, 93. https://doi.org/10.3390/pharmaceutics12020093
Chung EP, Cotter JD, Prakapenka AV, Cook RL, DiPerna DM, Sirianni RW. Targeting Small Molecule Delivery to the Brain and Spinal Cord via Intranasal Administration of Rabies Virus Glycoprotein (RVG29)-Modified PLGA Nanoparticles. Pharmaceutics. 2020; 12(2):93. https://doi.org/10.3390/pharmaceutics12020093
Chicago/Turabian StyleChung, Eugene P., Jennifer D. Cotter, Alesia V. Prakapenka, Rebecca L. Cook, Danielle M. DiPerna, and Rachael W. Sirianni. 2020. "Targeting Small Molecule Delivery to the Brain and Spinal Cord via Intranasal Administration of Rabies Virus Glycoprotein (RVG29)-Modified PLGA Nanoparticles" Pharmaceutics 12, no. 2: 93. https://doi.org/10.3390/pharmaceutics12020093
APA StyleChung, E. P., Cotter, J. D., Prakapenka, A. V., Cook, R. L., DiPerna, D. M., & Sirianni, R. W. (2020). Targeting Small Molecule Delivery to the Brain and Spinal Cord via Intranasal Administration of Rabies Virus Glycoprotein (RVG29)-Modified PLGA Nanoparticles. Pharmaceutics, 12(2), 93. https://doi.org/10.3390/pharmaceutics12020093