Curcumin, Curcumin Nanoparticles and Curcumin Nanospheres: A Review on Their Pharmacodynamics Based on Monogastric Farm Animal, Poultry and Fish Nutrition



Abstract
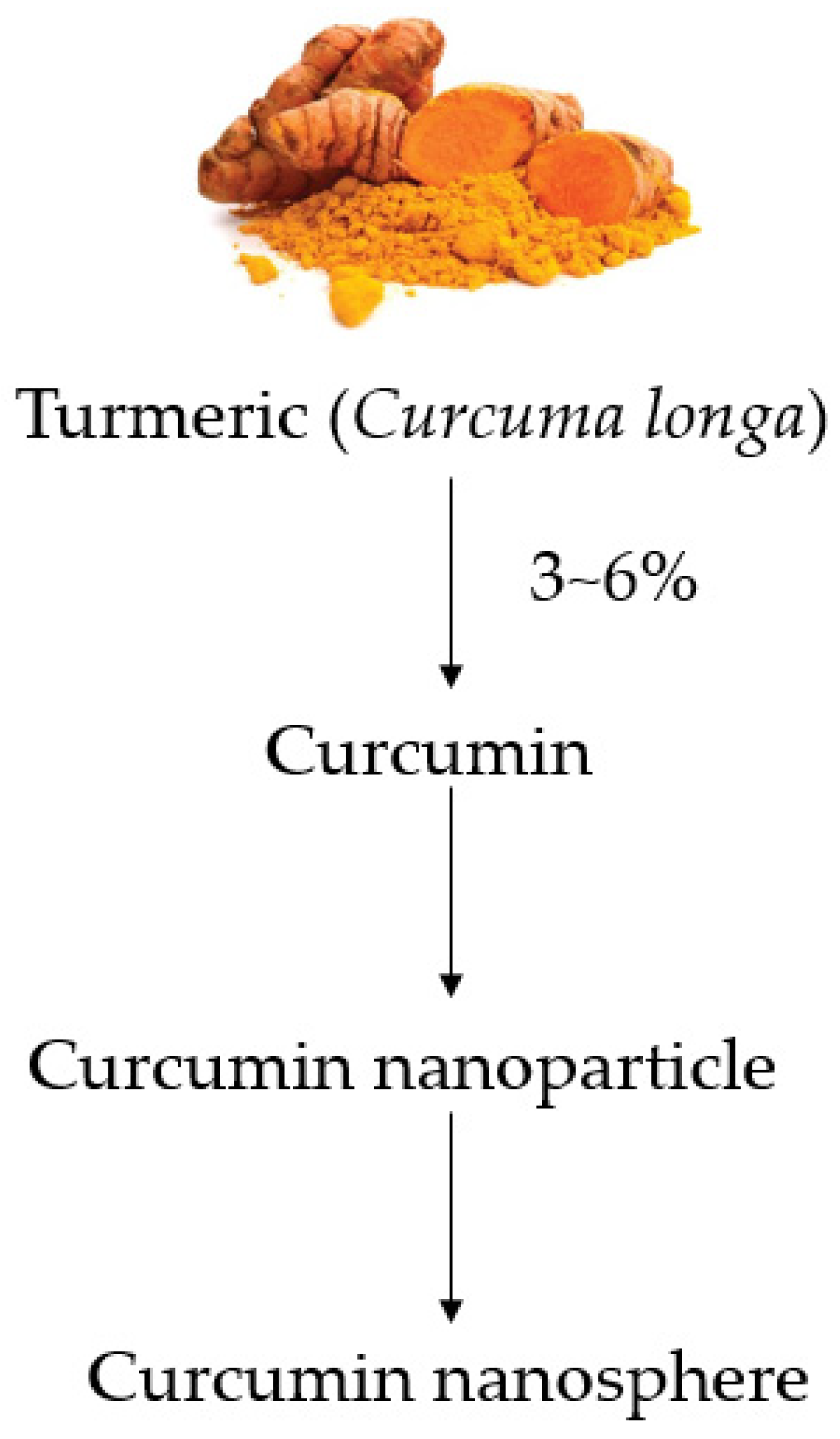
:1. Introduction
2. Curcumin or Nanocurcumin in Monogastric Farm Animal, Poultry and Fish Nutrition
2.1. Dietary Turmeric/Curcumin in Swine Nutrition
2.2. Dietary Turmeric/Curcumin in Poultry Nutrition
2.3. Dietary Turmeric and Curcumin in Rabbit Nutrition
2.4. Dietary Turmeric and Curcumin in Fish Nutrition
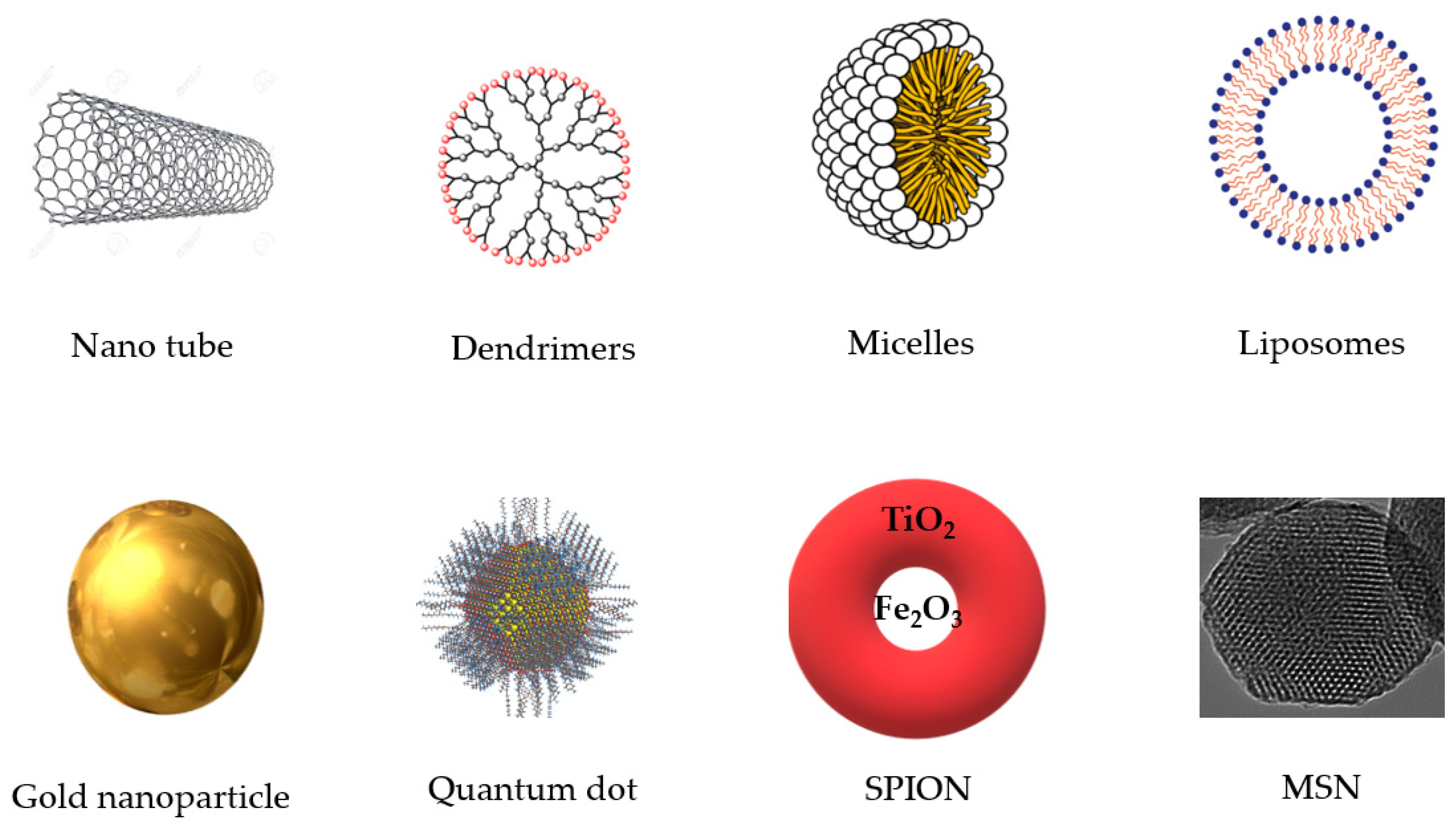
3. Curcumin Nanospheres in Monogastric Farm Animal, Poultry and Fish Nutrition
4. Conclusions and Future Outlook
Funding
Conflicts of Interest
References
- Toniguchi, N.; Arakawa, C.; Kobayashi, T. On the basic concept of nano-technology. In Proceedings of the International Conference on Production Engineering, Tokyo, Japan, 26–29 August 1974; Volume 2, pp. 18–23. [Google Scholar]
- Silva, G.A. Introduction to nanotechnology and its application to medicine. Surg. Neurol. 2004, 61, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Gopi, M.; Pearlin, B.; Kumar, R.D.; Shanmathy, M.; Prabakar, G. Role of nanoparticles in animal and poultry nutrition: Modes of action and applications in formulating feed additives and food processing. Int. J. Pharmacol. 2017, 13, 724–731. [Google Scholar] [CrossRef] [Green Version]
- Thulasi, A.; Rajendran, D.; Jash, S.; Selvaraju, S.; Jose, V.L.; Velusamy, S.; Mathivanan, S. Nanobiotechnology in Animal Nutrition. In Animal Nutrition and Reproductive Physiology (Recent Concepts), 1st ed.; Sampath, K.T., Ghosh, J., Eds.; Nanobiotechnology in Animal Nutrition; Satish Serial Publishing House: New Delhi, India, 2013; pp. 499–516. [Google Scholar]
- Uniyal, S.; Dutta, N.; Raza, M.; Jaiswal, S.K.; Sahoo, J.K.; Ashwin, K. Application of Nano Minerals in the Field of Animal Nutrition: A Review. Bull. Environ. Pharmacol. Life Sci. 2017, 6, 4–8. [Google Scholar]
- Sinha, R.; Kim, G.; Nie, S.; Shin, D. Nanotechnology in cancer therapeutics: Bioconjugated nanoparticles for drug delivery. Mol. Cancer Ther. 2006, 5, 1909–1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, S.A.; Srinivas, P.R.; Clifford, A.J.; Lee, S.C.; Philbert, M.A.; Hettich, R.L. New technologies for nutrition research. J. Nutr. 2004, 134, 681–685. [Google Scholar] [CrossRef] [Green Version]
- Schaffazick, S.R.; Pohlmann, A.R.; Mezzalira, G.; Guterres, S.S. Development of nanocapsule suspensions and nanocapsule spray-dried powders containing melatonin. J. Braz. Chem. Soc. 2006, 17, 562–569. [Google Scholar] [CrossRef]
- Bowman, K.; Leong, K.W. Chitosan nanoparticles for oral drug and gene delivery. Int. J. Nanomed. 2006, 1, 117–128. [Google Scholar] [CrossRef]
- Rahmani, M.; Golian, A.; Kermanshahi, H.; Bassami, M.R. Effects of curcumin and nanocurcumin on growth performance, blood gas indices and ascites mortalities of broiler chickens reared under normal and cold stress conditions. Italian J. Anim. Sci. 2017, 16, 438–446. [Google Scholar] [CrossRef] [Green Version]
- Johannah, N.M.; Joseph, A.; Maliakel, B.; Krishnakumar, I.M. Dietary addition of a standardized extract of turmeric (TurmaFEEDTM) improves growth performance and carcass quality of broilers. J. Anim. Sci. Tech. 2018, 60, 8. [Google Scholar]
- Marchiori, M.S.; Oliveira, R.C.; Souza, C.F.; Baldissera, M.D.; Ribeiro, Q.M.; Wagner, R.; Gundel, S.S.; Ourique, A.F.; Kirinus, J.K.; Stefani, L.M.; et al. Curcumin in the diet of quail in cold stress improves performance and egg quality. Anim. Feed Sci. Tech. 2019, 254, 144–157. [Google Scholar] [CrossRef]
- Al-Kassie, G.A.M.; Mohseen, A.M.; Abd-Al-Jaleel, R.A. Modification of productive performance and physiological aspects of broilers on the addition of a mixture of cumin and turmeric to the diet. Res. Opin. Anim. Vet. Sci. 2011, 1, 31–34. [Google Scholar]
- Galli, G.M.; Da Silva, A.S.; Biazus, A.H.; Reis, J.H.; Boiago, M.M.; Topazio, J.P.; Migliorini, M.J.; Guarda, N.S.; Moresco, R.N.; Ourique, A.F.; et al. Feed addition of curcumin to laying hens showed anticoccidial effect, and improved egg quality and animal health. Res. Vet. Sci. 2018, 118, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Gowda, N.K.; Ledoux, D.R.; Rottinghaus, G.E.; Bermudez, A.J.; Chen, Y.C. Antioxidant efficacy of curcuminoids from Turmeric (Curcuma longa L.) powder in broiler chickens fed diets containing aflatoxin B1. Br. J. Nutr. 2009, 102, 1629–1634. [Google Scholar] [CrossRef] [PubMed]
- Ruan, D.; Wang, W.C.; Lin, C.X.; Fouad, A.M.; Chen, W.; Xia, W.G.; Wang, S.; Luo, X.; Zhang, W.H.; Yan, S.J.; et al. Effects of curcumin on performance, antioxidation, intestinal barrier and mitochondrial function in ducks fed corn contaminated with ochratoxin A. Animal 2019, 13, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hu, Z.; Lu, C.; Bai, K.; Zhang, L.; Wang, T. Effect of various levels of dietary curcumin on meat quality and antioxidant profile of breast muscle in broilers. J. Agric. Food Chem. 2015, 63, 3880–3886. [Google Scholar] [CrossRef]
- Dar, A.H.; Rashid, N.; Majid, I.; Hussain, S.; Dar, M.A. Nanotechnology interventions in aquaculture and seafood preservation. Crit. Rev. Food Sci. Nutr. 2019, 27, 1–10. [Google Scholar] [CrossRef]
- Hill, E.K.; Li, J. Current and future prospects for nanotechnology in animal production. J. Anim. Sci. Biotech. 2017, 8, 26. [Google Scholar] [CrossRef] [Green Version]
- Yeh, Y.C.; Creran, B.; Rotello, V.M. Gold Nanoparticles: Preparation, Properties, and Applications in Bionanotechnology. Nanoscale 2012, 4, 1871–1880. [Google Scholar] [CrossRef]
- Jardón-Maximino, N.; Pérez-Alvarez, M.; Sierra-Ávila, R.; Carlos Alberto Ávila-Orta, C.A.; Jiménez-Regalado, E.; Bello, A.M.; González-Morones, P.; Cadenas-Pliego, G. Oxidation of Copper Nanoparticles Protected with Different Coatings and Stored under Ambient Conditions. J. Nannomat. 2018, 2018, 9512768. [Google Scholar] [CrossRef] [Green Version]
- Gunti, L.; Dass, R.S.; Kalagatur, N.K. Phytofabrication of Selenium Nanoparticles from Emblica officinalis Fruit Extract and Exploring Its Biopotential Applications: Antioxidant, Antimicrobial, and Biocompatibility. Front. Microbiol. 2019, 10, 931. [Google Scholar] [CrossRef] [Green Version]
- Phan, T.T.V.; Huynh, T.C.; Manivasagan, P.; Sudip Mondal, S.; Junghwan Oh, J. An Up-To-Date Review on Biomedical Applications of Palladium Nanoparticles. Nanomaterials 2020, 10, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Travan, A.; Pelillo, C.; Donati, I.; Marsich, E.; Benincasa, M.; Scarpa, T.; Semeraro, S.; Turco, G.; Gennaro, R.; Paoletti, S. Non-cytotoxic silver nanoparticle-polysaccharide nanocomposites with antimicrobial activity. Biomacromolecules 2009, 10, 1429–1435. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.J.; Oh, J.M.; Choy, J.H. Biocompatible nanoparticles intercalated with anticancer drug for target delivery: Pharmacokinetic and biodistribution study. J. Nanosci. Nanotechnol. 2010, 10, 2913–2916. [Google Scholar] [CrossRef] [PubMed]
- Shim, M.; Kam, N.W.S.; Chen, R.J.; Li, Y.; Dai, H. Functionalization of carbon nanotubes for biocompatibility and biomolecular recognition. Nano Lett. 2002, 2, 285–288. [Google Scholar] [CrossRef]
- Ichikawa, S.; Iwamoto, S.; Watanabe, J. Formation of biocompatible nanoparticles by self-assembly of enzymatic hydrolysates of chitosan and carboxymethyl cellulose. Biosci. Biotechnol. Biochem. 2005, 69, 1637–1642. [Google Scholar] [CrossRef] [Green Version]
- Taylor, T.M.; Davidson, P.M. Liposomal nanocapsules in food science and agriculture. Crit. Rev. Food Sci. Nutr. 2005, 45, 587–605. [Google Scholar] [CrossRef]
- Konkol, D.; Wojnarowski, K. The Use of Nanominerals in Animal Nutrition as a Way to Improve the Composition and Quality of Animal Products. J. Chem. 2018, 2018, 5927058. [Google Scholar] [CrossRef]
- Cervantes-Valencia, M.E.; Alcala-Canto, Y.; Salem, A.Z.M.; Kholif, A.E.; Ducoing-Watty, A.M.; Bernad-Bernad, M.J.; Gutiérrez-Olvera, C. Influence of curcumin (Curcuma longa) as a natural anticoccidial alternative in adult rabbits: First results. Ital. J. Anim. Sci. 2015, 14, 3838. [Google Scholar] [CrossRef]
- Xun, W.; Shi, L.; Zhou, H.; Hou, G.; Cao, T.; Zhao, C. Effects of curcumin on growth performance, jejunal mucosal membrane integrity, morphology and immune status in weaned piglets challenged with enterotoxigenic E. coli. Int. Immunopharmacol. 2015, 27, 46–52. [Google Scholar] [CrossRef]
- Sruthi, M.V.; Nair, A.B.; Arun, D.; Thushara, V.V.; Sheeja, C.C.; Vijayasree, A.S.; Oommen, O.V.; Divya, L. Dietary curcumin influences leptin, growth hormone and hepatic growth factors in Tilapia (Oreochromis mossambicus). Aquaculture 2018, 496, 105–111. [Google Scholar] [CrossRef]
- Grenni, P.; Ancona, V.; Caracciolo, A.B. Ecological effects of antibiotics on natural ecosystems: A review. Microchem. J. 2018, 136, 25–39. [Google Scholar] [CrossRef]
- EU. Ban on Antibiotics as Growth Promoters in Animal Feed Enters into Effect. Brussels. 2005. Available online: http://europa.eu/rapid/press-release_IP-05-1687_en.htm (accessed on 6 March 2020).
- FDA, Center for Veterinary Medicine. FDA Reminds Retail Establishments of Upcoming Changes to the Use of Antibiotics in Food Animals; FDA, Center for Veterinary Medicine: Rockville, Maryland, 2016. Available online: https://www.fda.gov/AnimalVeterinary/NewsEvents/CVMUpdates/ucm507355.htm (accessed on 6 March 2020).
- Samarasinghe, K.; Wenk, C.; Silva, K.F.S.T.; Gunasekera, J.M.D.M. Turmeric (Curcuma longa) Root Powder and Mannanoligosaccharides as Alternatives to Antibiotics in Broiler Chicken Diets. Asian-Aust. J. Anim. Sci. 2003, 16, 1495–1500. [Google Scholar] [CrossRef]
- Prasad, S.; Tyagi, A.K.; Aggarwal, B.B. Recent developments in delivery, bioavailability, absorption and metabolism of curcumin: The golden pigment from golden spice. Cancer Res. Treat. 2014, 46, 2–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, G.; Kapoor, I.P.S.; Singh, P.; de Heluani, C.S.; de Lampasona, M.P.; Catalan, C.A.N. Comparative study of chemical composition and antioxidant activity of fresh and dry rhizomes of turmeric (Curcuma Longa Linn). Food Chem. Toxicol. 2010, 48, 1026–1031. [Google Scholar] [CrossRef]
- Moghadamtousi, S.Z.; Kadir, H.A.; Hassandarvish, P.; Tajik, H.; Abubakar, S.; Zandi, K. A Review on Antibacterial, Antiviral, and Antifungal Activity of Curcumin. BioMed Res. Int. 2014, 2014, 186864. [Google Scholar]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A Review of Its’ Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef]
- Kharat, M.; McClements, D.J. Recent advances in colloidal delivery systems for nutraceuticals: A case study—Delivery by Design of curcumin. J. Coll. Inter. Sci. 2019, 557, 506–518. [Google Scholar] [CrossRef]
- Rai, M.; Pandit, R.; Gaikwad, S.; Yadav, A.; Gade, A. Potential applications of curcumin and curcumin nanoparticles: From traditional therapeutics to modern nanomedicine. Nanotech. Rev. 2015, 4, 161–172. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Bhatt, I.B.; Ichikawa, H.; Ahn, K.S.; Sethi, G.; Sandur, S.K.; Natarajan, C.; Seeram, N.; Shishodia, S. Curcumin biological and medicinal properties. In Turmeric: The Genus Curcuma; Ravindran, P.N., Nirmal Babu, K., Sivaraman, K., Eds.; CRC Press: New York, NY, USA, 2007; pp. 297–368. [Google Scholar]
- Zheng, B.; Peng, S.; Zhang, X.; McClements, D.J. Impact of Delivery System Type on Curcumin Bioaccessibility: Comparison of Curcumin-Loaded Nanoemulsions with Commercial Curcumin Supplements. J. Agric. Food Chem. 2018, 66, 10816–10826. [Google Scholar] [CrossRef]
- Gera, M.; Sharma, N.; Ghosh, M.; Huynh, D.L.; Lee, S.J.; Min, T.; Kwon, T.; Jeong, D.K. Nanoformulations of curcumin: An emerging paradigm for improved remedial application. Oncotarget 2017, 8, 66680–66698. [Google Scholar] [CrossRef] [Green Version]
- Wahlstrom, B.; Blennow, G. A study on the fate of curcumin in the rat. Acta Pharmacol. Toxicol. (Copenh) 1978, 43, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Patil, V.M.; Das, S.; Balasubramanian, K. Quantum Chemical and Docking Insights into Bioavailability Enhancement of Curcumin by Piperine in Pepper. J. Phys. Chem. A 2016, 120, 3643–3653. [Google Scholar] [CrossRef] [PubMed]
- Ghalandarlaki, N.; Alizadeh, A.M.; Ashkani-Esfahani, S. Nanotechnology-Applied Curcumin for Different Diseases Therapy. BioMed Res. Int. 2014, 2014, 394264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurita, T.; Makino, Y. Novel curcumin oral delivery systems. Anticancer Res. 2013, 33, 2807–2821. [Google Scholar]
- Hani, U.; Shivakumar, H.G. Solubility enhancement and delivery systems of curcumin a herbal medicine: A review. Curr. Drug Deliv. 2014, 11, 792–804. [Google Scholar] [CrossRef]
- Ma, Z.; Shayeganpour, A.; Brocks, D.R.; Lavasanifar, A.; Samuel, J. High-performance liquid chromatography analysis of curcuminin rat plasma: Application to pharmacokinetics of polymericmicellar formulation of curcumin. Biomed. Chromatogr. 2007, 21, 546–552. [Google Scholar] [CrossRef]
- Song, Z.; Feng, R.; Sun, M.; Guo, C.; Gao, Y.; Li, L.; Zhai, G. Curcumin-loaded PLGA-PEG-PLGA triblock copolymeric micelles: Preparation, pharmacokinetics and distribution in vivo. J. Colloid Interface Sci. 2011, 354, 116–123. [Google Scholar] [CrossRef]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The Essential Medicinal Chemistry of Curcumin. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef]
- Sobh, R.A.; Mohamed, W.S.; Moustafa, A.B.; Nasr, H.E. Encapsulation of Curcumin and Curcumin Derivative in Polymeric Nanospheres. Polymer-Plastics Tech. Eng. 2015, 54, 1457–1467. [Google Scholar] [CrossRef]
- Pignanelli, C.; Ma, D.; Noel, M.; Ropat, J.; Mansour, F.; Curran, C.; Pupulin, S.; Larocque, K.; Wu, J.; Liang, G.; et al. Selective Targeting of Cancer Cells by Oxidative Vulnerabilities with Novel Curcumin Analogs. Sci. Rep. 2017, 7, 1105. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, S.A.; El-Shishtawy, R.M.; Al-Bar, O.A.M.; Al-Najada, A.R. Chemical modification of curcumin: Solubility and antioxidant capacity. Int. J. Food Prop. 2017, 20, 718–724. [Google Scholar] [CrossRef]
- Kunwar, A.; Barik, A.; Pandey, R.; Priyadarsini, K.I. Transport of liposomal and albumin loaded curcumin to living cells: An absorption and fluorescence spectroscopic study. Biochim. Biophys. Acta (Gen.) 2006, 1760, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Priyadarsini, K.I. The chemistry of curcumin: From extraction to therapeutic agent. Molecules 2014, 19, 20091–20112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangwar, R.K.; Tomar, G.B.; Dhumale, V.A.; Zinjarde, S.; Sharma, R.B.; Datar, S. Curcumin conjugated silica nanoparticles for improving bioavailability and its anticancer applications. J. Agric. Food Chem. 2013, 61, 9632–9637. [Google Scholar] [CrossRef]
- Singh, S.P.; Sharma, M.; Gupta, P.K. Enhancement of phototoxicity of curcumin in human oral cancer cells using silica nanoparticles as delivery vehicle. Lasers Med. Sci. 2014, 29, 645–652. [Google Scholar] [CrossRef]
- Ma’Mani, L.; Nikzad, S.; Kheiri-Manjili, H.; Al-Musawi, S.; Saeedi, M.; Askaralou, S. Curcumin-loaded guanidine functionalized PEGylated mesoporous silica nanoparticles KIT-6: Practical strategy for the breast cancer therapy. Eur. J. Med. Chem. 2014, 83, 646–654. [Google Scholar] [CrossRef]
- Natarajan, S.K.; Selvaraj, S. Mesoporous silica nanoparticles: Importance of surface modifications and its role in drug delivery. RSC Adv. 2014, 4, 14328. [Google Scholar] [CrossRef]
- Baldissera, M.D.; Souza, C.F.; Zeppenfeld, C.C.; Descovi, S.N.; Machado, V.S.; Santos, R.C.V.; Baldisserotto, B. Efficacy of dietary curcumin supplementation as bactericidal for silver catfish against Streptococcus agalactiae. Microb. Pathog. 2018, 116, 237–240. [Google Scholar] [CrossRef]
- Mahmoud, H.K.; Al-Sagheera, A.A.; Redab, F.M.; Mahgoub, S.A.; Ayyata, M.S. Dietary curcumin supplement influence on growth, immunity, antioxidant status, and resistance to Aeromonas hydrophila in Oreochromis niloticus. Aquaculture 2017, 475, 16–23. [Google Scholar] [CrossRef]
- Xie, Z.; Shen, G.; Wang, Y.; Wu, C. Curcumin supplementation regulates lipid metabolism in broiler chickens. Poult. Sci. 2018, 98, 422–429. [Google Scholar] [CrossRef]
- Galli, G.M.; Gerbet, R.R.; Griss, L.G.; Fortuoso, B.F.; Petrolli, T.G.; Boiago, M.M.; Souza, C.F.; Matheus, D.; Baldissera, M.D.; Mesadri, J.; et al. Combination of herbal components (curcumin, carvacrol, thymol, cinnamaldehyde) in broiler chicken feed: Impacts on response parameters, performance, fatty acid profiles, meat quality and control of coccidia and bacteria. Microb. Pathol. 2020, 139, 103916. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, M.; da Rocha, B.A.; Francisco, C.R.L.; Miranda, C.G.; Santos, P.D.F.; de Araújo, P.H.H.; Sayer, C.; Leimann, F.V.; Goncalves, O.H.; Bersani-Amado, C.A. Evaluation of the in vivo acute antiinflammatory response of curcumin loaded nanoparticles. Food. Funct. 2018, 24, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; He, J.; Zhao, W.; Shen, M.; Zhang, L.; Zhong, X.; Wang, C.; Wang, T. Effect of Curcumin on Growth Performance, Inflammation, Insulin level, and Lipid Metabolism in Weaned Piglets with IUGR. Animals 2019, 9, 1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, E.; Zhang, J.; Han, H.; Wu, J.; Gan, Z.; Wei, C.; Zhang, L.; Wang, C.; Wang, T. Curcumin Alleviates IUGR Jejunum Damage by Increasing Antioxidant Capacity through Nrf2/Keap1 Pathway in Growing Pigs. Animals 2020, 10, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaguezeski, A.M.; Gündel, S.S.; Favarin, F.R.; Gündel, A.; Souza, C.F.; Baldissera, M.D.; Cazarotto, C.C.; Volpato, A.; Fortuoso, B.F.; Ourique, A.F. Low-dose curcumin-loaded Eudragit L-100-nanocapsules in the diet of dairy sheep increases antioxidant levels and reduces lipid peroxidation in milk. J. Food Eng. 2019, 43, e12942. [Google Scholar] [CrossRef]
- Baldissera, M.D.; Souza, C.F.; Zeppenfeld, C.C.; Velho, M.C.; Klein, B.; Abbad, L.B.; Ourique, A.F.; Wagner, R.; Da Silva, A.S.; Baldisserotto, B. Dietary supplementation with nerolidol nanospheres improves growth, antioxidant status and fillet fatty acid profiles in Nile tilapia: Benefits of nanotechnology for fish health and meat quality. Aquaculture 2020, 516, 734635. [Google Scholar] [CrossRef]
- Kiczorowska, B.; Samolińska, W.; Al-Yasiry, A.R.M.; Kiczorowski, P.; Winiarska-Mieczan, A. The natural feed additives as immunostimulants in monogastric animal nutrition—A review. Ann. Anim. Sci. 2017, 17, 605–625. [Google Scholar] [CrossRef] [Green Version]
- Guil-Guerrero, J.L.; Ramos, L.; Paredes, J.C.; Carlosama-Yépez, M.; Moreno, C.; Ruales, P. Effects of turmeric rhizome powder and curcumin on poultry production. A review. J. Anim. Feed Sci. 2017, 26, 293–302. [Google Scholar] [CrossRef]
- Seven, P.T.; Seven, I.; Baykalir, B.G.; Mutlu, S.I.; Salem, A.Z.M. Nanotechnology and nano-propolis in animal production and health: An overview. Italian J. Anim. Sci. 2018, 17, 921–930. [Google Scholar] [CrossRef] [Green Version]
- Shah, B.R.; Mraz, J. Advances in nanotechnology for sustainable aquaculture and fisheries. Rev. Aquacult. 2019, 1–18. [Google Scholar] [CrossRef]
- Epstein, J.; Sanderson, I.R.; MacDonald, T.T. Curcumin as a therapeutic agent: The evidence from in vitro, animal and human studies. British J. Nutr. 2010, 103, 1545–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, B.D.; Grace, D.; Sones, K. Current drivers and future directions of global livestock disease dynamics. Proc. Natl. Acad. Sci. USA 2013, 110, 20871–20877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.C.; Hansen, C.F.; Mullana, B.P.; Pluske, J.R. Nutrition and pathology of weaner pigs: Nutritional strategies to support barrier function in the gastrointestinal tract. Anim. Feed Sci. Technol. 2012, 173, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Wijtten, P.J.; van der Meulen, J.; Verstegen, M.W. Intestinal barrier function and absorption in pigs after weaning: A review. Br. J. Nutr. 2011, 105, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Pluske, J.R. Feed- and feed additives-related aspects of gut health and development in weanling pigs. J. Anim. Sci. Biotech. 2013, 4, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maneewan, C.; Yamauchi, K.E.; Mekbungwan, A.; Maneewan, B.; Siri, S. Effects of turmeric (Curcuma longa Linnaeus) on growth performance, nutrient digestibility, hematological values, and intestinal histology in nursery pigs. J. Swine Health Prod. 2012, 20, 231–240. [Google Scholar]
- Liu, Y.; Song, M.; Che, T.M.; Almeida, J.A.; Lee, J.J.; Bravo, D.; Maddox, C.W.; Pettigrew, J.E. Dietary plant extracts alleviate diarrhea and alter immune responses of weaned pigs experimentally infected with a pathogenic Escherichia coli. J. Anim. Sci. 2013, 91, 5294–5306. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Song, M.; Che, T.M.; Bravo, D.; Maddox, C.W.; Pettigrew, J.E. Effects of capsicum oleoresin, garlic botanical, and turmeric oleoresin on gene expression profile of ileal mucosa in weaned pigs. J. Anim. Sci. 2014, 92, 3426–3440. [Google Scholar] [CrossRef] [Green Version]
- Ilsley, S.E.; Miller, H.M.; Kamel, C. Effects of dietary quillaja saponin and curcumin on the performance and immune status of weaned piglets. J. Anim. Sci. 2005, 83, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Gan, Z.; Wei, W.; Li, Y.; Wu, J.; Zhao, Y.; Zhang, L.; Wang, T.; Zhong, X. Curcumin and Resveratrol Regulate Intestinal Bacteria and Alleviate Intestinal Inflammation in Weaned Piglets. Molecules 2019, 24, 1220. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Xu, H.; Xia, D.; Zhao, R. Curcumin attenuates the effects of transport stress on serum cortisol concentration, hippocampal NO production, and BDNF expression in the pig. Domestic Anim. Endocrinol. 2010, 39, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lillehoj, H.S.; Jang, S.I.; Lillehoj, E.P.; Min, W.; Bravo, D.M. Dietary supplementation of young broiler chickens with Capsicum and turmeric oleoresins increases resistance to necrotic enteritis. Br. J. Nutr. 2013, 110, 840–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salih, N.H. Effect of Turmeric (Curcuma longa) powder on growth performance, carcass traits, meat quality and serum biochemical parameters in broilers. J. Adv. Biomed. Path. Res. 2013, 3, 25–32. [Google Scholar]
- Nayaka, H.B.S.; Umakantha, B.; Ruban, S.W.; Murthy, H.N.N.; Narayanaswamy, H.D. Performance and hematological parameters of broilers fed neem, turmeric, vitamin e and their combinations. Emir. J. Food Agric. 2013, 25, 483–488. [Google Scholar] [CrossRef] [Green Version]
- Qasem, M.A.A.; Alhajj, M.S.; Ger El Nabi, A.R.; Al-Mufarrej, S.I. Effect of turmeric powder as a dietary supplement on performance indicators and immune responses in broiler chickens. J. Anim. Vet. Adv. 2015, 14, 30–35. [Google Scholar]
- Wang, D.; Huang, H.; Zhou, L.; Li, W.; Zhou, H.; Hou, G.; Liu, J.; Hu, L. Effects of dietary supplementation with turmericrhi zome extract on growth performance, carcass characteristics, antioxidant capability, and meat quality of Wenchang broiler chickens. Ital. J. Anim. Sci. 2015, 14, 344–349. [Google Scholar] [CrossRef]
- Akhavan-Salamat, H.; Ghasemi, H.A. Alleviation of chronic heat stress in broilers by dietary supplementation of betaine and turmeric rhizome powder: Dynamics of performance, leukocyte profile, humoral immunity, and antioxidant status. Trop. Anim. Health Prod. 2016, 48, 181–188. [Google Scholar] [CrossRef]
- Hassan, S.M. Effects of adding different dietary levels of turmeric (Curcuma longa Linn) powder on productive performance and egg quality of laying hens. Int. J. Poult. Sci. 2016, 15, 156–160. [Google Scholar] [CrossRef] [Green Version]
- Mirbod, M.; Mahdavi, A.H.; Samie, A.H.; Mehri, M. Effects of Curcuma longa rhizome powder on egg quality, performance, and some physiological indices of laying hens fed different levels of metabolizable energy. J. Sci. Food Agric. 2017, 97, 1286–1294. [Google Scholar] [CrossRef]
- Kilany, O.E.; Mahmoud, M.M. Turmeric and exogenous enzyme supplementation improve growth performance and immune status of Japanese quail. Worlds Vet. J. 2014, 4, 20–29. [Google Scholar] [CrossRef]
- Saraswati, T.R.; Tana, S. Physiological condition of first female and male offspring of Japanese quail (Coturnix japonica) whose parents were supplemented by turmeric powder. J. World Poult. Res. 2016, 6, 59–65. [Google Scholar]
- Kim, D.K.; Lillehoj, H.S.; Lee, S.H.; Jang, S.I.; Lillehoj, E.P.; Bravo, D. Dietary Curcuma longa enhances resistance against Eimeria maxima and Eimeria tenella infections in chickens. Poult. Sci. 2013, 92, 2635–2643. [Google Scholar] [CrossRef] [PubMed]
- Hang, T.T.T.; Molee, W.; Khempaka, S.; Paraksa, N. Supplementation with curcuminoids and tuna oil influenced skin yellowness, carcass composition, oxidation status, and meat fatty acids of slow-growing chickens. Poult. Sci. 2017, 97, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Rajput, N.; Muhammad, N.; Yan, R.; Zhong, X.; Wang, T. Effect of dietary supplementation of curcumin on growth performance, intestinal morphology and nutrients utilization of broiler chicks. J. Poult. Sci. 2013, 50, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Rajput, R.; Ali, S.; Naeem, M.; Khan, M.A.; Wang, T. The effect of dietary supplementation with the natural carotenoids curcumin and lutein on pigmentation, oxidative stability and quality of meat from broiler chickens affected by a coccidiosis challenge. Br. Poult. Sci. 2014, 55, 501–509. [Google Scholar] [CrossRef]
- Zhang, J.; Bai, K.; Su, W.; Wang, A.; Zhang, L.; Huang, K.; Wang, T. Curcumin attenuates heat-stress-induced oxidant damage by simultaneous activation of GSH-related antioxidant enzymes and Nrf2-mediated phase II detoxifying enzyme systems in broiler chickens. Poult. Sci. 2018, 97, 1209–1219. [Google Scholar] [CrossRef]
- Zhang, J.; Bai, K.W.; He, J.; Niu, Y.; Lu, Y.; Zhang, L.; Wang, T. Curcumin attenuates hepatic mitochondrial dysfunction through the maintenance of thiol pool, inhibition of mtDNA damage, and stimulation of the mitochondrial thioredoxin system in heat-stressed broilers. J. Anim. Sci. 2018, 96, 867–879. [Google Scholar] [CrossRef] [Green Version]
- Salah, A.S.; Mahmoud, M.A.; Ahmed-Farid, O.A.; El-Tarabany, M.S. Effects of dietary curcumin and acetylsalicylic acid supplements on performance, muscle amino acid and fatty acid profiles, antioxidant biomarkers and blood chemistry of heat-stressed broiler chickens. J. Therm. Biol. 2019, 84, 259–265. [Google Scholar] [CrossRef]
- Zhang, N.Y.; Qi, M.; Zhu, M.K.; Guo, J.; Liu, J.; Gu, C.Q.; Rajput, S.A.; Krumm, C.S.; Qi, D.S.; Sun, L.H. Curcumin Prevents Aflatoxin B1 Hepatoxicity by Inhibition of Cytochrome P450 Isozymes in Chick Liver. Toxins 2016, 8, 327. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, I.; Sun, X.; Wang, H.; Li, W.; Wang, X.; Cheng, P.; Li, S.; Zhang, X.; Hamid, S. Curcumin Successfully Inhibited the Computationally Identified CYP2A6 Enzyme-Mediated Bioactivation of Aflatoxin B1 in Arbor Acres broiler. Front. Pharmacol. 2017, 8, 143. [Google Scholar] [CrossRef] [Green Version]
- Nawab, A.; Li, G.; Liu, W.; Lan, R.; Wu, J.; Zhao, Y.; Kang, K.; Kieser, B.; Sun, C.; Tang, S.; et al. Effect of Dietary Curcumin on the Antioxidant Status of Laying Hens under High-Temperature Conditions. Braz. J. Poult. Sci. 2019, 21, 1–10. [Google Scholar] [CrossRef]
- Sahin, K.; Orhan, C.; Tuzcu, Z.; Tuzcu, M.; Sahin, N. Curcumin ameloriates heat stress via inhibition of oxidative stress and modulation of Nrf2/HO-1 pathway in quail. Food Chem. Toxicol. 2012, 50, 4035–4041. [Google Scholar] [CrossRef] [PubMed]
- Ruan, D.; Zhu, Y.W.; Fouad, A.M.; Yan, S.J.; Chen, W.; Zhang, Y.N.; Xia, W.G.; Wang, S.; Jiang, S.Q.; Yang, L.; et al. Dietary curcumin enhances intestinal antioxidant capacity in ducklings via altering gene expression of antioxidant and key detoxification enzymes. Poult. Sci. 2019, 98, 3705–3714. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, M.; Golian, A.; Kermanshahi, H.; Bassami, M.R. Effects of curcumin or nanocurcumin on blood biochemical parameters, intestinal morphology and microbial population of broiler chickens reared under normal and cold stress conditions. J. Appl. Anim. Res. 2018, 46, 200–209. [Google Scholar] [CrossRef] [Green Version]
- Basavaraj, M.; Nagabhushana, V.; Prakash, N.; Mallikarjunappa, S.; Appannavar, M.M.; Wagmare, P. Effect of dietary supplementation of Pulvis Curcuma longa on the voluntary feed intake, nutrient digestibility and growth performance of broiler rabbits under summer stress. Vet. World 2010, 3, 369–372. [Google Scholar]
- Guerreiro, I.; Oliva-Teles, A.; Enes, P. Prebiotics as functional ingredients: Focus on Mediterranean fish aquaculture. Rev. Aquacult. 2018, 10, 800–832. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Khanongnuch, C.; Kanpiengjai, A.; Unban, K.; Kim, V.V.; Srichaiyo, S. Host-associated probiotics boosted mucosal and serum immunity, disease resistance and growth performance of Nile tilapia (Oreochromis niloticus). Aquaculture 2018, 491, 94–100. [Google Scholar] [CrossRef]
- Rajabiesterabadi, H.; Hoseini, S.M.; Fazelan, Z.; Hoseinifar, S.H.; Doan, H.V. Effects of dietary turmeric administration on stress, immune, antioxidant and inflammatory responses of common carp (Cyprinus carpio) during copper exposure. Aquacult. Nutr. 2020, 1–11. [Google Scholar] [CrossRef]
- Yonar, M.E.; Yonar, S.M.; İspir, U.; Ural, M.S. Effects of curcumin on haematological values, immunity, antioxidant status and resistance of rainbow trout (Oncorhynchus mykiss) against Aeromonas salmonicida subsp. Achromogenes. Fish. Shell. Immun. 2019, 89, 83–90. [Google Scholar] [CrossRef]
- Elgendy, M.; Hakim, A.; Ibrahim, T.; Soliman, W.; Ali, S. Immunomodulatory effects of curcumin on nile tilapia, Oreochromis niloticus and its antimicrobial properties against Vibrio alginolyticus. J. Fish. Aquat. Sci. 2016, 11, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Liu, B.; Ge, X.; Xieb, J.; Xub, P.; Miao, L.; Sun, S.; Liaoa, Y.; Chen, R.; Ren, M.; et al. Effects of dietary curcumin on growth performance, biochemical parameters, HSP70 gene expression and resistance to Streptococcus iniae of juvenile Gift Tilapia, Oreochromis niloticus. Isr. J. Aquacult. Bamidgeh 2017, 66, 986–996. [Google Scholar]
- El-Barbary, M.I. Detoxification and antioxidant effects of garlic and curcumin in Oreochromis niloticus injected with aflatoxin B1 with reference to gene expression of glutathione peroxidase (GPx) by RT-PCR. Fish. Physiol. Biochem. 2016, 42, 617–629. [Google Scholar] [CrossRef] [PubMed]
- El-Barbary, M.I. Impact of garlic and curcumin on the hepatic histology and cytochrome P450 gene expression of aflatoxicosis Oreochromis niloticus using RT-PCR. Turk. J. Fish. Aquat. Sci. 2018, 18, 405–415. [Google Scholar]
- Manju, M.; Akbarsha, M.A.; Oommen, O.V. In vivo protective effect of dietary curcumin in fish Anabas testudineus (Bloch). Fish Physiol. Biochem. 2012, 38, 309–318. [Google Scholar] [CrossRef]
- Manju, M.; Vijayasree, A.S.; Akbarsha, M.A.; Oommen, O.V. Protective effect of dietary curcumin in Anabas testudineus (Bloch) with a special note on DNA fragmentation assay on hepatocytes and micronucleus assay on erythrocytes in vivo. Fish. Physiol. Biochem. 2013, 39, 1323–1330. [Google Scholar] [CrossRef]
- Manju, M.; Sherin, T.G.; Rajasekharan, K.N.; Oommen, O.V. Curcumin analogue inhibits lipid peroxidation in a freshwater teleost, Anabas testudineus (Bloch)-an in vitro and in vivo study. Fish. Physiol. Biochem. 2009, 35, 413–420. [Google Scholar] [CrossRef]
- Cao, L.; Ding, W.; Du, J.; Jia, R.; Liu, Y.; Zhao, C.; Shen, Y.; Yin, G. Effects of curcumin on antioxidative activities and cytokine production in Jian carp (Cyprinus carpio var. Jian) with CCl 4-induced liver damage. Fish. Shell. Immunol. 2015, 43, 150–157. [Google Scholar] [CrossRef]
- Jiang, J.; Wu, X.; Zhou, X.; Feng, L.; Liu, Y.; Jiang, W.; Wu, P.; Zhao, Y. Effects of dietary curcumin supplementation on growth performance, intestinal digestive enzyme activities and antioxidant capacity of crucian carp Carassius auratus. Aquaculture 2016, 463, 174–180. [Google Scholar] [CrossRef]
- Erdagi, S.I.; Uyanik, C. Biological evaluation of bioavailable amphiphilic polymeric conjugate based-on natural products: Diosgenin and curcumin. Int. J. Poly. Mat. Poly. Biol. 2020, 69, 73–84. [Google Scholar] [CrossRef]
- Vaiserman, A.; Koliada, A.; Zayachkivska, A.; Lushchak, O. Nanodelivery of Natural Antioxidants: An Anti-aging Perspective. Front. Bioeng. Biotechnol. 2020, 7, 447. [Google Scholar] [CrossRef] [Green Version]
- Faehnrich, B.; Lukas, B.; Humer, E.; Zebeli, Q. Phytogenic pigments in animal nutrition: Potentials and risks. J. Sci. Food Agric. 2016, 96, 1420–1430. [Google Scholar] [CrossRef] [PubMed]
- Hossen, S.; Hossain, M.K.; Basher, M.K.; Mia, M.N.H.; Rahman, M.T.; Uddin, M.J. Smart nanocarrier-based drug delivery systems for cancer therapy and toxicity studies: A review. J. Adv. Res. 2019, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Lee, Y.M.; Kim, D.W.; Min, T.S.; Lee, S.J. Nanosphere Loaded with Curcumin Inhibits the Gastrointestinal Cell Death Signaling Pathway Induced by the Foodborne Pathogen Vibrio vulnificus. Cells 2020, 9, 631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suwannateep, N.; Banlunara, W.; Wanichwecharungruang, S.P.; Chiablaem, K.; Lirdprapamongkol, K.; Svasti, J. Mucoadhesive curcumin nanospheres: Biological activity, adhesion to stomach mucosa and release of curcumin into the circulation. J. Contr. Rel. 2011, 151, 176–182. [Google Scholar] [CrossRef]
- Balasubramanian, K. Molecular Orbital Basis for Yellow Curry Spice Curcumin’s Prevention of Alzheimer’s Disease. J. Agric. Food Chem. 2006, 54, 3512–3520. [Google Scholar] [CrossRef]
- Balasubramanian, K. Quantum chemical insights into Alzheimer’s disease: Curcumin’s chelation with Cu(II), Zn(II), and Pd(II) as a mechanism for its prevention. Int. J. Quant. Chem. 2016, 116, 1107–1119. [Google Scholar] [CrossRef]
- Yang, R.; Zhang, S.; Kong, D.; Gao, X.; Zhao, Y.; Wang, Z. Biodegradable Polymer-Curcumin Conjugate Micelles Enhance the Loading and Delivery of Low-Potency Curcumin. Pharm. Res. 2012, 29, 3512–3525. [Google Scholar] [CrossRef]
- Liu, C.H.; Chang, F.Y. Development and Characterization of Eucalyptol Microemulsions for Topic Delivery of Curcumin. Chem. Pharm. Bull. 2011, 59, 172–178. [Google Scholar] [CrossRef] [Green Version]
- Huo, X.; Zhang, Y.; Jin, X.; Yongang Li, Y.; Li, Z. A novel synthesis of selenium nanoparticles encapsulated PLGA nanospheres with curcumin molecules for the inhibition of amyloid β aggregation in Alzheimer’s disease. J. Photochem. Photobiol. B Biol. 2019, 190, 98–102. [Google Scholar] [CrossRef]
- Yavarpour-Bali, H.; Ghasemi-Kasman, M.; Pirzadeh, M. Curcumin-loaded nanoparticles: A novel therapeutic strategy in treatment of central nervous system disorders. Int. J. Nanomed. 2019, 14, 4449–4460. [Google Scholar] [CrossRef] [Green Version]
- Abdelghany, S.; Tekko, I.A.; Vora, L.; Larraneta, E.; Permana, A.D.; Donnelly, R.F. Nanosuspension-Based Dissolving Microneedle Arrays for Intradermal Delivery of Curcumin. Pharmaceutics 2019, 11, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dende, C.; Meena, J.; Nagarajan, P.; Nagaraj, V.A.; Panda, A.K.; Padmanaban, G. Nanocurcumin is superior to native curcumin in preventing degenerative changes in Experimental Cerebral Malaria. Sci. Rep. 2017, 7, 10062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldissera, M.D.; Souza, C.F.; da Silva, A.S.; Velho, M.C.; Ourique, A.F.; Baldisserotto, B. Benefits of nanotechnology: Dietary supplementation with nerolidol-loaded nanospheres increases survival rates, reduces bacterial loads and prevents oxidative damage in brains of Nile tilapia experimentally infected by Streptococcus agalactiae. Microb. Pathol. 2020, 141, 103989. [Google Scholar] [CrossRef] [PubMed]
- Baldissera, M.D.; Souza, C.F.; Velho, M.C.; Bassotto, V.A.; Ourique, A.F.; da Silva, A.S.; Baldisserotto, B. Nanospheres as a technological alternative to suppress hepatic cellular damage and impaired bioenergetics caused by nerolidol in Nile tilapia (Oreochromis niloticus). Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 751–759. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Types | Animal Category and Duration of Exposure | Diet Preparation and Experimental Design | Experimental Findings | Source |
|---|---|---|---|---|
| Turmeric | 50- to 60 -days-old weaned, crossbred pigs (Large white × Landrace × Duroc), and duration was 85–90 days of age for Experiment 1 and 175 to 185 days of age for Experiment 2. | Total 48 nursery pigs (~15 kg) with four treatments fed 0.00%, 0.05%, 0.10% and 0.20% turmeric in feed. Experiment 1, visceral organ and epithelial cell morphology on villus was observed by SEM at 30 kg BW. Experiment 2, blood was collected at 40 and 80 kg BW, growth compared at 90 kg BW | ↔ Growth, carcass and visceral organ weight did not differ. ↑ nutrient digestibility at 0.10%, ↓ hematocrit at 0.10% turmeric, ↑ WBC at 0.10% and 0.20%, ↑ cell mitosis in jejunum and ileum at 0.10% and 0.20% diets | [81] |
| 21-days-old weaned piglets (G-Performer × Fertilium 25)/11 days of trial | Total 64 weaned piglets (~6 kg) in 2 × 4 factorial design with or without F-18 Escherichia coli challenged; four diets were basal or 10 ppm of capsicum oleoresin, garlic botanical, or turmeric oleoresin added | ↑ average daily weight gain, ↓ ileal macrophages, diarrhea score and frequency of diarrhea, ↔ growth in diets with plant extract and challenged, F-18 E. coli | [82] | |
| 21-days-old weaned piglets (G-Performer × Fertilium 25)/9 days of trial | Total 64 weaned piglets (~6 kg) divided into 4 treatments; diets were: a nursery basal diet, basal diet supplemented with 10 mg/kg of capsicum oleoresin, garlic botanical, or turmeric oleoresin | ↑ expression of immunity-related genes in ileal mucosa of pigs treated with plant extracts which enhanced intestinal health status and immunity in pigs | [83] | |
| Curcumin | 21-days-old weaned piglets (Duroc × Landrace × Yorkshire)/21 days of trial | Total 50 weaned piglets (~6 kg) divided into 5 dietary treatments: control (basal diet), and the basal diet supplemented with 50 mg/kg quinocetone, or 200 mg/kg, 300 mg/kg or 400 mg/kg curcumin | ↓ FCR, crypt depth, IELs, mRNA levels of IL-1β and TLR4, and TNF-α as well as ↑ villus height, goblet cells, mRNA level of IL-10 in pigs fed with 50 mg/kg quinocetone, or 300 or 400 mg/kg curcumin | [31] |
| 26-days-old weaned piglets (Landrace × Yorkshire)/24 days of trial | Total 40 weaned piglets (~6 kg) divided into 4 dietary treatments: NBW with control diet, NC (NBW with 400 mg/kg curcumin in control diet), IUGR with control diet, and IC (IUGR with 400 mg/kg curcumin in control diet) | ↓ growth and ↑ serum pro-inflammatory cytokines, insulin resistance, hepatic lipid contents in pigs with IUGR and vice-versa in curcumin fed piglets; ↑ hepatic insulin-signaling and lopogenic pathway by IUGR but positive attenuation by IC | [68] | |
| 26-days-old weaned female crossbred piglets (Duroc × Landrace × Large White)/26 to 115 days of age | Total 24 weaned piglets divided into 3 dietary treatments: IUGR group, NBW group, and IUGR + CUR group, which were fed diets containing 0 mg/kg (NBW), 0 mg/kg (IUGR) and 200 mg/kg (IUGR + CUR) curcumin | ↑ SOD, ↓ MDA in jejunum of IUGR pigs fed CUR diet; ↑ Nrf2, GCLC, SOD1, GCLM, and NQO1 mRNA gene expressions in IUGR+CUR diet; ↓ TNFα, IL-6, IFNγ, caspase3, bax, bcl2, hsp70 ↑ IL-2, ocln expression in IUGR + CUR | [69] | |
| 29-days-old weaned piglets (Large white × Landrace × Duroc)/20 days of trial | Total 192 weaned piglets (~6 kg) divided into 4 dietary levels: diets with or without (as-fed basis) quillaja saponin (750 mg/kg during week 1, 300 mg/kg during weeks 2 to 3) and with or without dietary curcumin (200 mg/kg) | ↔ Growth in all treatments; ↓ ADFI, FE in quillaja saponin fed diets (days 15–20); ↑ IgG and CRP in saponin fed pigs; ↔ villus and crypt values in small intestine of all treatments; | [84] | |
| 28-days-old weaned piglets (Duroc × Large white × Landrace)/28 days of trial | Total 180 (90 ♂ + 90 ♀) weaned piglets (~8 kg) divided into 6 dietary treatments: control diet (CON group) or supplemented diet (300 mg/kg of antibiotics, ANT group; 300 mg/kg of RES and CUR, respectively, HRC group; 100 mg/kg of RES and CUR, respectively, LRC group; 300 mg/kg of RES; 300 mg/kg CUR) | ↓ IL-1β, TNF-α expressions and down-regulated TLR4 signaling pathway, ↑ secretion of IgA, IgG in RES + CUR or HRC fed diet in weaned piglets | [85] | |
| 60-days-old male crossbred piglets (Pietrain × Erhualian)/21 days of trial | Total 18 piglets (~16 kg) divided into 3 dietary treatments: control (not treated with curcumin and not subjected to transport (CON), transport control (not treated with curcumin and subjected to 2 h transport together with the curcumin treated group, T-CON), and curcumin (treated with curcumin (8 mg/kg, p.o.) for 21 days and subjected to 2 h of transport thereafter, CUR) groups | ↑ serum cortisol, hippocampal NO content, ↓ BDNF expression in pigs not treated with CUR; ↓ stress response, cNOS, total NOS, iNOS in CUR treated pigs; | [86] |
| Types | Animal Category and Duration of Exposure | Diet Preparation and Experimental Design | Experimental Findings | Source |
|---|---|---|---|---|
| Turmeric | 1-day-old broiler chickens (Ross/Ross)/20-days of trial | Total 45 chickens divided into three groups; diets are: control diet, control diet supplemented with 4 mg capsicum oleoresin and 4 mg turmeric oleoresin/kg diet (XT) and uninfected or orally challenged with Eimeria maxima oocysts at 14 days and Clostridium perfringens at 18 days of age | ↑ BW, ↓ gut lesion score, ↓ serum α-toxin, intestinal IL-8, LITAF, IL-17A and IL-17F mRNA levels, ↑ cytokine /chemokine in splenocytes in the XT-group compared with the birds fed the control diet | [87] |
| 1-day-old broiler chickens (Ross)/ 6-weeks trial | Total 240 chicks allotted into 4 groups; diets are: basal, turmeric powder (TP) supplemented at 5, 7, 9 g/kg of basal | ↑ BW, liver and gizzard index, ↓ serum Cho, TG at 7 g/kg TP compared with basal diet | [88] | |
| 288-days-old broiler chicks (Raja II)/ 42-days of trial | Total 288 broilers; basal diet with neem (8 g/kg), turmeric (2 g/kg) and vitamin E (0.2 g/kg) individually and in combination to form 8 test diet groups | ↑ hematological parameters in neem fed diet; ↓ Hb in turmeric fed diet; ↑ PCV in neem, turmeric and vit E fed than basal diet | [89] | |
| 1-day-old broiler chickens (Ross 308)/ 42-days of trial | Total 288 broilers; control diet with turmeric powder, TP at 10, 12, 14, 16, 18 and 20 g/kg, and vaccinated (positive control) or unvaccinated (negative control) to form 8 test diets | ↓ BW and DFI in 20 g/kg TP fed, ↔ NDV or IBV in chicks fed TP diets compared with positive control diet; TP did not improve growth and immunity of chicks | [90] | |
| 1-day-old Wenchang broiler chickens/ 12-weeks of trial | Total 300 broilers; basal diet with turmeric rhizome extract, TRE at 0, 100, 200 and 300 mg/kg to from 4 diets | ↑ growth performance, antioxidant activity, breast muscle ↓ abdominal fat in TRE fed diets | [91] | |
| 1-day-old Ross male chicks/ 42-days of trial | Total 625 chicks divided into 5 dietary treatment groups: thermoneutral control (TN-CON), heat stress (HS-CON), heat stressed supplemented with betaine (HS-Bet), HS with TRP (HS-TRP), and HS-BT (fed Bet and TRP) | ↑ heterophil, total and IgM antibody in HS-TRP and HS-BT than HS-CON group; ↓ MDA in supplemented group than HS diet; ↑ GPx, SOD supplemented group than to TN and HS-CON groups | [92] | |
| 52-weeks-old Hisex laying hens/8-w trial | Total 150 laying hens; control diet with turmeric powder, TP at 0, 2 and 4% to form three dietary treatment groups | ↔ BW, egg weight or production at 4% TP diet; 2% TP had little effect on production, egg quality | [93] | |
| 37-weeks-old Leghorn laying hens/ 70-days of trial | Total 160 laying hens assigned to 8 diets in 2 × 4 factorial design at 2 levels of AME (11.51 and 12.35 MJ/kg) and 4 levels of CRP (0, 2, 4 and 6 g/kg) | ↑ egg shell thickness and hardness but ↓ yolk cholesterol as ↑ CRP; ↓ TG, ALT, AST and ↑ villus length, goblet cells at 2 g/kg CRP of diet | [94] | |
| 1-week-old Japanese quails/4-weeks trial | Total 150 quails allotted to 5 diets: FM -CON (T1), SBM-CON (T2), SBM with 0.5% TP (T3), SBM with 0.5% TP and 0.1% phytase (T4), SBM with 0.5% TP and 0.1% panzyme (T5) | ↑ BW, FE, total protein, albumin, SOD, CAT, GSH, GSH-Px but ↓ MDA at T3 to T5 diets compared with control diets | [95] | |
| 1st stage: 15-days-old Japanese quails/1-month trial 2nd stage: 1-month-old Japanese quails/3-month trial | 1st stage: 45 female quails divided into 3 groups like control (P0), control with 54 mg (P1) and 108 mg (P2) of TP/quail/day; 2nd stage: 3 groups like K0: offspring from parents of P0, K1: offspring from parents of P1 and K1: offspring from parents of P2 diets | After 3 months or 2nd stage: ↑ vitellogenin, HDL, vit-B12, vit-A, white egg protein, LA, ARA but ↓ Cho, LDL, total fat in eggs; ↑ carcass weight, SGPT, Cho, TG in serum, Cho in liver, but ↓ LDL and SGOT serum in female offspring | [96] | |
| Curcumin | 1-day-old chicks/ 42-days of trial | Total 180 chicks in three diet groups: control diet, control diet with lipophilic turmeric extract containing curcumin and turmerones, TF-36 at 0.5% and 1% to from 3 treatment groups | ↑ BW and antioxidant activity, ↓ lipid peroxidation, ↔ meat color, serum creatinine, total protein, liver enzyme activity in 1% TF-36 fed diet than control | [11] |
| 1-day-old broiler (Ross×Ross)/ 14-days of trial | Two diet groups- one standard diet (control) with no curcumin and another is control with 35 mg/kg of a freeze dried Curcuma longa extract; At 14 day posthatch, noninfected or infected by oral gavage with 2.0 × 104 oocysts of Eimeria maxima or E. tenella | ↑ BW and ↓ fecal oocyst, gut lesion as well as ↑ serum antibodies, cellular immunity, in C. longa fed diets; dietary C. longa showed coccidiosis resistance against Eimeria maxima or E. tenella | [97] | |
| 30-weeks-old laying hens (Hy-Line Brown)/ 21-days of trial | Total 60 laying hens in 3 diet groups: control (T0), control diet with curcumin at 30 (T30) and 50 (T50) mg/kg of curcumin, respectively | ↑ TAC, specific gravity and yolk index of eggs, and ↓ yolk color, TBARS in eggs and ↓ oocysts in feces in the T30 and T50 fed diets | [14] | |
| 1-day-old male chicks (Cobb 500)/ 44-days of trial | Total 225 male chicks in 5 diet groups: NC-negative control feed; PC-positive control; CU-with 50 mg/kg of curcumin, PHY-100 mg/kg phytogenic; and PHY + CU, a combination of both additives at 50 mg/kg (curcumin) and 100 mg/kg (phytogenic). | ↑ total protein, globulin, ↓ uric acid, Cho, TG, oocysts, bacteria in PHY + CU diet; ↓ SFA and ↑ MUFA or PUFA in CU or PHY+CU fed groups; ↑ crypt/ villus ratio, intestinal health in PHY + CU fed diets | [66] | |
| 1-day-old male broiler chicks (Ross × Ross)/ 21-days of trial | Total 180 chicks in 6 dietary groups: basal diet, basal diet with total curcuminoids (TCMN) at 444 mg/kg, basal diet with AFB1 at 1.0 mg/kg, basal diet with 74 mg/kg TCMN and 1.0 mg/kg AFB1, basal diet with 222 mg/kg TCMN and 1.0 mg/kg AFB1, basal diet with 444 mg/kg TCMN and 1.0 mg/kg AFB1 | ↑ BW, FE in 74 and 222 mg/kg TCMN at AFB1 diets; ↑ total protein, albumin, antioxidant function, γ-glutamyl transferase activity in 222 mg/kg TCMN at AFB1 diet | [15] | |
| 21-days-old mixed sex slow-growing chickens (Thai indigenous crossbred)/63-days of trial | Total 480 chicks in six diet groups: basal diet with 4% tuna oil as positive control, basal diet added with curcumin removed turmeric oleoresin at 20, 40, 60 or 80 mg/kg curcuminoids (CUR20, CUR40, CUR60, CUR80, respectively) or dl-α-tocopheryl acetate at 200 ppm as negative control (E-200) | ↑ FCR, breast fillet, yellowness of skin in curcuminoids fed diets; ↑ LA but ↓ DHA of breast meat in CUR20 and CUR40 fed diets; ↓ TBARS in chicken meat in CUR60 fed diet; suitable level of curcuminoids in slow-growing chickens was 60 mg/kg diet | [98] | |
| 1-day-old Arbor Acres broiler chickens/ 42-days of trial | Total 400 chicks in four diet groups: control (CRM0), basal diet added with 100 (CRM100), 150 (CRM150) and 200 (CRM200) mg/kg curcumin | ↑ BW, FE in CRM200 fed diet; ↑ APE and ↓ abdominal fat in CRM150 and CRM200 fed diets; ↑ villus length and width in intestine up to CRM200 fed diets | [99] | |
| 1-day-old Arbor Acres broiler chickens/ 21-days of trial | Total 200 chicks in four diet groups: basal diet without carotenoid (control), basal diet added with 300 curcumin (CRM), basal diet added with 300 lutein (LTN) or with a combination (C + L) of 150 mg/kg curcumin and 150 mg/kg lutein; All chickens were challenged with Eimeria maxima at 21 d old | ↑ redness and yellowness of fresh meat in C + L fed diet; ↓ MDA and carbonyl in CRM and C + L fed diets but ↑ sulfhydryl in C + L birds; ↑ myosin chain in carotenoid fed diets; CRM or C + L are efficient natural antioxidant to preserve meat quality and resistant against coccidiosis | [100] | |
| 21-days-old male Arbor Acres broiler chickens/ 42-days of trial | Total 320 chicks in four diet groups: basal diet (C1), basal diet added with 50, 100 or 200 mg/kg curcumin (C2, C3, C4, respectively) | ↑ redness value of meat, CAT, ABTS radical scavenging activity and ↓ drip loss at 48 h in curcumin supplemented groups | [17] | |
| 21-days-old male Arbor Acres broiler chickens/ 20-days of trial | Total 400 chicks in five diet groups: basal diet + 22 °C (CON), 34 °C for 8 h (0900-1700) + basal diet supplemented with 0, 50, 100 or 200 mg/kg curcumin (HS, CMN1, CMN2, and CMN3 treatments, respectively) | ↓ FCR in CMN1 & CMN2 diets; ↑ liver GSH in CMN1 & CMN2; ↑ γ-GCLm, GSH-Px, GST in in curcumin fed diets; ↑ Nrf2, HO-1, γ-GCLc expressions in curcumin fed diets; ↑ Cu/ZnSOD, CAT in CMN2 than the HS treatment; ↓ MDA, AST, ALT and ↑ MnSOD, mtDNA, ATP in curcumin groups; | [101,102] | |
| 1-day-old male broiler chickens (Ross 308)/ 49-days of trial | Total 1200 chicks in four diet groups: control diet, control diet supplied with 500, 1000 and 2000 mg/kg curcumin | ↓ BW, ADG, liver weight in 1000 and 2000 mg/kg CUR groups; ↓ plasma LDL and hepatic TG in 2000 mg/kg curcumin group; ↑ hepatic glycogen and hepatic lipase activities in 1000 and 2000 mg/kg curcumin groups; ↓ FAS, SREBP-1c gene expression in all curcumin group; ↑ PPARα, CPT-I expressions in 1000 and 2000 mg/kg CUR groups | [65] | |
| 160-days-old male chicks (Ross 308)/ 42-days of trial | Total 160 chicks in four diet groups: basal diet in thermoneutral condition (23 °C), basal diet in 8 h thermal stress (34 °C), basal diet with 100 mg/kg curcumin (CR) at thermal stress (34 °C), basal diet with 1 g/kg acetylsalicylic acid (ASA) at thermal stress (34 °C) | ↑ ADFI, ADG, FBW in CR added than the other diets; ↓ MDA in CR and ASA diets; ɔ PUFA, ALA, DHA in breast muscle of broiler with CR supplied diet at thermal stress (34 °C) | [103] | |
| 1-day-old male broiler chickens/ 12-weeks of trial | Total 120 chicks in four diet groups: a 2 × 2 factorial design was used where the main factors included adding aflatoxin B1, AFB1 (< 5 vs. 100 µg/kg) and curcumin, CM (0 vs. 150 mg/kg) in a corn/soybean-based diet | ↑ liver injury, ALT, AST, MDA, but ↓ albumin, total protein, CAT, GSH, GSH-Px and induced AFBO-DNA in AFB1 fed; these attributes are lowered, prevented or protected by CM added diets | [104] | |
| 1-day-old male Arbor Acres broiler chickens/ 28-days of trial | Total 120 chicks in six diet groups: control group, curcumin alone-treated group (450 mg/kg feed), the group fed AFB1-contaminated feed (5 mg/kg feed) plus the low (150 mg), medium (300 mg) or high (450 mg) of curcumin, and the group fed AFB1-contaminated diet alone (5 mg/kg feed) | ↓ liver weight and toxicity, ↑ body weight in curcumin treated groups; ↑ mRNA, protein expressions and CYP2A6 enzyme activity in AFB1-fed group; however, ↓ mRNA, protein expressions and CYP2A6 enzyme activity on dose dependent manner in curcumin fed | [105] | |
| 22-weeks-old Roman laying hens/ 21-days of trial | Total 336 laying hens in 3 diet groups: first group as a thermoneutral control (25 °C), second group at high temp (32 °C, 6 h/day), given a basal diet, third group was five treatment groups (100, 150, 200, 250, 300 mg/kg curcumin) (H1, H2, H3, H4, H5, respectively) fed a basal diet under high temp conditions (32 °C, 6 h/day) | ↑ SOD at H2 and H3 fed diets; ↑ total antioxidant capacity at H2, H3 and H5 fed diets; ↑ CAT and GSH-Px at H3 diet; ↓ MDA in curcumin added diets; ↑ CAT, SOD, GSH-Px and T-AOC in liver, heart and lungs of curcumin treated groups compared with heat stressed control group | [106] | |
| 10-days-old Japanese quails/ 42-days of trial | Total 180 birds reared at either 22 °C (thermoneutral) or 34 °C (heat stress) for 8 h/day (0900-1700) until the age of 42 days. Birds in both environments were randomly fed 1 of 3 diets: basal diet and basal diet added with 0, 200 or 400 mg of curcumin per kg of diet. | ↑ BW, FI, and ↓ FE, MDA, nuclear factor, HSP70 in response to increasing supplemental curcumin level in the diets | [107] | |
| 1-day-old White Pekin ducklings/ 21-days of trial | Total 540 mixed-sex birds in three dietary treatments: controls (fed only the basal diet), a group fed an OTA-contaminated diet (2 mg/kg feed), and a group fed the same OTA-feed plus 400 mg/kg of curcumin | ↑ BW, ADG, and no enterotoxicity in curcumin fed diet compared to OTA diet; ↓ interleukin-1β, tumor necrosis factor-α, MDA, apoptotic gene expression, mt-transcription factors and ↑ GSH, jejunal mucosa, tight junction protein in curcumin fed diets than OTA diet | [16] | |
| 1-day-old male Cherry Valley Pekin ducklings/ 21-days of trial | Total 720 male ducklings in four dietary treatments: control group were fed a basal diet and the remainder were fed the basal diet supplemented with 200, 400, or 800 mg/kg curcumin | ↑ jejunal and hepatic curcumin contents with 400 and 800 mg/kg; ↑ SOD1, CAT, GST, GPX1, HO-1, MRP6, CYP1A4, CYP2D17, Nrf2 transcript, ABCB1 and ↓ CYP1B1, CYP2A6 expressions in jejunal mucosa in curcumin fed diets; | [108] | |
| Nanocurcumin | 30-days-old Japanese quails/ 21-days of trial | Total 60 birds in four diet groups: control group (T0 - without CUR), free CUR (T30 - 30 mg/kg) and two doses of CUR in nanocapsules (T3 and T10 - nanocapsules containing 3 and 10 mg of curcumin /kg of feed, respectively) | ↓ FCR and ↑ egg production in T30 and T10 fed diets; ↓TBARS in egg yolk and ↑ antioxidant capacity against peroxyl radicals from T30, T3, and T10 fed diets compared to T0 diet; ↓ SFA and PUFA in egg yolk of T10 fed diet; ↑ MUFA in egg yolk of T10 and T30 fed diets | [12] |
| 1-day-old male broiler chickens (Ross 308)/ 42-days of trial | Total 500 chicks in five diet groups divided into two identical houses: diets were (1) control; (2) and (3) Control + 200 or 400 mg/kg curcumin; (4) and (5) Control + 200 or 400 mg/kg nanocurcumin, respectively under recommended temp up to 14 days, when the temp was dropped in one house from 28.5 to 13–15 °C and maintained at this level to induce ascites until 42 days. Whereas, in the second house the temperature was maintained according to the hybrid production guidelines | ↓ WG and ↑ FCR in birds reared in cold temp than the normal temp; ↑ blood pCO2, HTC, and ↓ pO2, O2 saturation in cold stress at 42 d of age which is alleviated by curcumin/nanocurcumin added diets; ↑ BW, villus surface in intestine and ↓ MDA, liver enzymes, caecal E. coli population in curcumin or nanocurcumin fed diets | [10,109] |
| Types | Animal Category and Duration of Exposure | Diet Preparation and Experimental Design | Experimental Findings | Source |
|---|---|---|---|---|
| Turmeric | 84-weeks-old weaned broiler rabbits/ 8 weeks of trial | Three treatment groups: basal diet, basal diet added with turmeric rhizome powder, turmeric rhizome powder (TRP)at 0.15% or 0.30% in the diets | no beneficial effect of dietary TRP on blood biochemical and meat characteristics of broiler rabbits reared under summer stress | [110] |
| Curcumin | 8-months-old New Zealand white rabbits/ 42 days of trial | Total 24 rabbits in four groups: control diet, control diet with 10, 25 and 40 mg aqueous extract of curcumin /kg body weight | curcumin decreased Eimeria spp. oocysts excretion efficiently at a dose of 40 mg/kg BW with 80.1%, 63.7% and 64.9% for days 28, 35 and 42, respectively, with reducing concentration of eggs per gram of feces with about 20.1, 15.6 and 17.8 for days 14, 21 and 35, respectively | [30] |
| Types | Animal Category and Duration of Exposure | Diet Preparation and Experimental Design | Experimental Findings | Source |
|---|---|---|---|---|
| Turmeric | Common carp, Cyprinus carpio fingerlings (average body weight 42 g)/3 weeks for Experiment 1 and 3 weeks for Experiment 2 | Experiment 1, total 240 fingerlings in four groups: diets supplemented with 0, 5, 10 and 20 g/kg turmeric for 3 weeks, then fish were exposed to lethal 3.5 mg/L copper for 24 h; Experiment 2, total 300 fish with same dietary groups simultaneously exposed to sub-lethal 0.25 mg/L ambient copper for 3 weeks | Experiment 1, mortality was 65.3%, 41.8%, 22.7% and 20.6% according to dietary treatments; Experiment 2, ↓ plasma cortisol, glucose, MDA, ALT, AST, and ↑ plasma T3, T4, lysozyme, ACH50, bactericidal activities, SOD, CAT, GSH-Px, RBC, Hb, and ↓ TNF-α, IL-1β, ↑ IL10 genes in turmeric fed at 10 g/kg diet | [113] |
| Curcumin | Rainbow trout (Oncorhynchus mykiss) juveniles (average body weight 31 g)/ 8 weeks of trial | Total 300 juveniles in four groups: control was fed with the basal diet without CUR, the remaining groups were fed with 1% (E1), 2% (E2), 4% (E3) CUR; fish were challenged by Aeromonas salmonicida subsp. achromogenes | ↑ WG, SGR, survival in E2 group; ↓ FCR, RBC, Hb, Ht in CU-fed fish; ↑ SOD, CAT, GSH-Px but ↓ MDA in liver, head kidney and spleen in CU-fed fish; ↑ hematological, immunological, antioxidant activities in fish of E2 group | [114] |
| Silver catfish (Rhamdia quelen) juveniles (average body weight 205 g)/ 14 days of trial | Total 40 fingerlings in four groups: A-uninfected and non-supplemented fish (negative control), B-uninfected fish with 150 mg/kg CUR, C- infected & non-supplemented fish (positive control), and D- infected & added fish with 150 mg/kg CUR | 100% disease resistance against Streptococcus agalactiae and ↓ disease sign, erratic swimming, corneal opacity, skin lesions in fin and tail, ↑ appetite in fish fed the CUR-supplemented diet; | [63] | |
| Tilapia Oreochromis. niloticus, juveniles (average body weight 2.5 g)/ 84 days of trial | Total 300 juveniles in five groups: supplemented with 0 (basal diet), 50, 100, 150 or 200 mg CUR/kg diet (CUR50, CUR100, CUR150, CUR200, respectively) | ↑ WG, ADG, SGR, lysozyme, IgG, IgM in CUR50 group; ↑ FCR, PER, CP, CL in CUR100 and CUR150; ↑ CAT, GSH, ↓ MDA and survival challenged with A. hydrophila in all CUR fed | [64] | |
| Tilapia O. niloticus, fingerlings (average body weight 3 g)/ 100 days of trial | Total 180 juveniles in three groups: supplemented with 0% (basal group), 0.5%, or 1% CUR in the basal diets for juvenile tilapia | ↑ growth, FE and GH, IGF-1, IGF-2 in fish fed the CUR supplemented diets; ↓ blood glucose, FI, leptin genes, ↑ glycogenesis in CUR fed diets | [32] | |
| Nile tilapia O. niloticus, fingerlings (average body weight 45 g)/30 days of trial | Total 60 fingerlings in two groups: commercial diets supplemented with 0% (control) and 2% CUR (weight/weight) at 2% body weight | ↑ peroxidase, serum bactericidal activity, serum protein in CUR fed diet; 100% survivability in CUR fed diet challenged with V. alginolyticus | [115] | |
| Gift tilapia O. niloticus, juveniles (average body weight 13 g)/8 weeks of trial | Total 375 juveniles in five groups: basal diet was supplemented with 0 (control), 50, 100, 150 and 200 mg CUR/kg diets | ↑ WG, FBW, SGR, ↓ CF, FCR, MDA in 150 mg/kg CUR fed diet; ↑ Serum total protein, liver HSP70 in fish fed 100-200 mg/kg CUR diets | [116] | |
| Tilapia O. niloticus, fingerlings (average body weight 40 g)/ 14 days of trial | Total 180 fingerlings in ten groups: T1, negative control; T2, injected IP with AFB1 at 6 mg/kg in basal diet; T3-T6 were fed with garlic (T3 and T4) and CUR (T5 and T6) at 10 and 20 g/kg diet, respectively; T7-T10 injected IP with AFB1 and fed both garlic and CUR at 10 and 20 g/kg diet | ↓ HSI ↑ hepatic lesion in AFB1 group; ↑ HSI ↓ hepatic lesion in garlic + CUR fed groups at 10 mg/kg diet; ↑ CYP1A expression in CUR fed diet; ↓ toxicity of AFB1 in garlic fed diet | [117,118] | |
| Anabas testudineus, juveniles (average body weight 40 g)/ 8 weeks of trial | Total 72 juveniles in three groups: diets supplemented with 0% (basal group), 0.5%, or 1% CUR in 40% protein feeds | ↓ lipid peroxidation, TBARS, but ↑ GSH and normal hepatocytes, hepatopancreas, melanocytes in fish fed the CUR fed diets | [119,120] | |
| Anabas testudineus, juveniles/ 60 days of trial | Salicylcurcumin (0.5%) as CUR analog was supplemented along with the basal diet | ↓ TBARS, GSH, CAT, GSH-Px, ↑ SOD, protein content CUR fed fish | [121] | |
| Jian carp (Cyprinus carpio var. Jian), juveniles (average body weight 30 g)/ 60 days of trial | Juvenile carp were fed 0.1%, 0.5%, or 1.0% curcumin for 60 days, then injected IP with 30% CCl4 solution | ↓ AST, ALT, hepatocyte degeneration, MDA ↑ SOD, antioxidant capacity, GSH in liver and ↓ TNF-α, IL-1β in fish fed 0.5% and 1.0% curcumin | [122] | |
| Crucian carp (Carasius auratus), juveniles (average body weight 76 g)/ 105-days of trial | Total 585 juveniles in three groups: diets supplemented with 0 (basal group), 1, or 5 g/kg CUR | ↑ FW, PWG, FE, in 5 g/kg CUR diet; ↑ hepatopancreas weight, protein content, trypsin and lipase activities, AKP, γ-GT, CK activities in intestine, but in ↓ MDA, protein carbonyl content in 5 g/kg CUR diet; ↑ SOD, CAT, GR, GSH, GSH-Px, GST in intestine of CUR fed diets; ↑ trypsin, lipase, NKA, AKP, γGT, CK, SOD1, CAT, GSH-Px, GST, and GR gene expressions in intestine of fish fed the 5 g/kg CUR diet | [123] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moniruzzaman, M.; Min, T. Curcumin, Curcumin Nanoparticles and Curcumin Nanospheres: A Review on Their Pharmacodynamics Based on Monogastric Farm Animal, Poultry and Fish Nutrition. Pharmaceutics 2020, 12, 447. https://doi.org/10.3390/pharmaceutics12050447
Moniruzzaman M, Min T. Curcumin, Curcumin Nanoparticles and Curcumin Nanospheres: A Review on Their Pharmacodynamics Based on Monogastric Farm Animal, Poultry and Fish Nutrition. Pharmaceutics. 2020; 12(5):447. https://doi.org/10.3390/pharmaceutics12050447
Chicago/Turabian StyleMoniruzzaman, Mohammad, and Taesun Min. 2020. "Curcumin, Curcumin Nanoparticles and Curcumin Nanospheres: A Review on Their Pharmacodynamics Based on Monogastric Farm Animal, Poultry and Fish Nutrition" Pharmaceutics 12, no. 5: 447. https://doi.org/10.3390/pharmaceutics12050447
APA StyleMoniruzzaman, M., & Min, T. (2020). Curcumin, Curcumin Nanoparticles and Curcumin Nanospheres: A Review on Their Pharmacodynamics Based on Monogastric Farm Animal, Poultry and Fish Nutrition. Pharmaceutics, 12(5), 447. https://doi.org/10.3390/pharmaceutics12050447