
Oral Gene Therapy of HFD-Obesity via Nonpathogenic Yeast Microcapsules Mediated shRNA Delivery

Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of IL-1β shRNA Vectors
2.2. Adipocyte Culture
2.3. Function of IL-1β shRNA in Lipid Metabolism
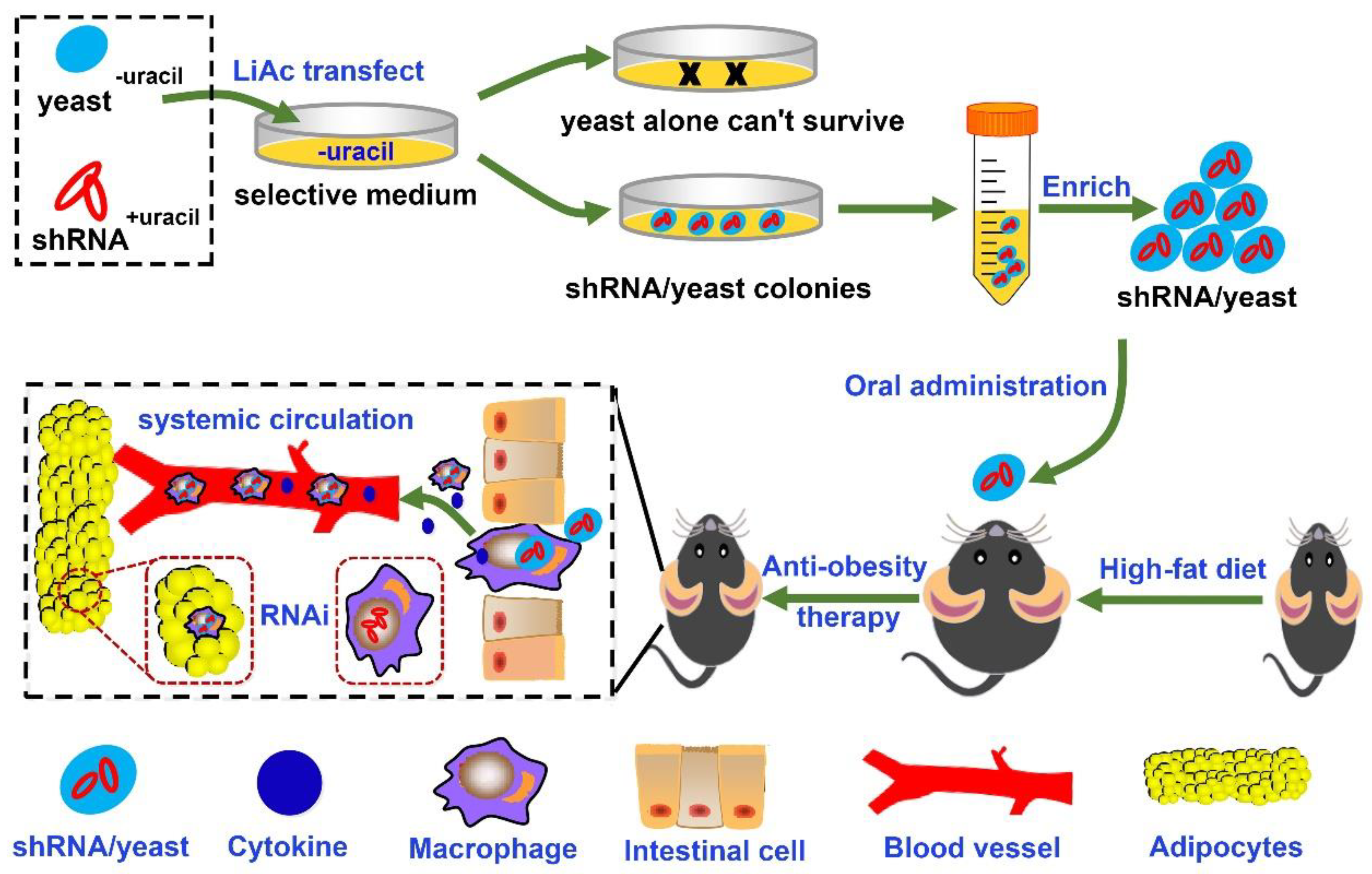
2.4. Preparation of IL-1β shRNA/Yeast Microcapsules
2.5. Function Detect of shRNA/Yeast in Macrophage
2.6. Mice and Diets
2.7. Glucose Tolerance Test
2.8. Small Intestinal Macrophage Isolation
2.9. RT-qPCR and ELISA Assay
2.10. Histological Staining
2.11. Statistical Analysi
3. Results
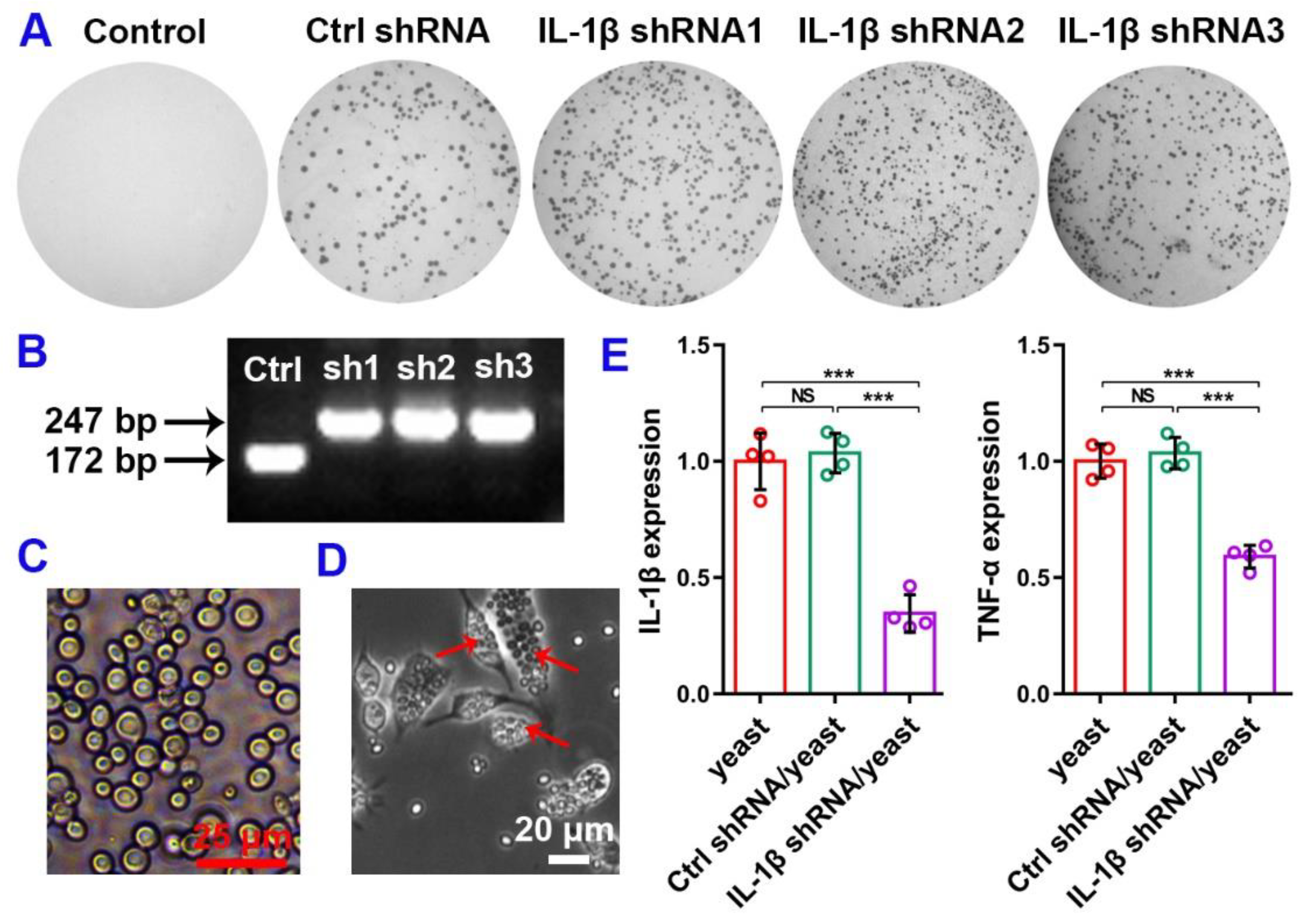
3.1. IL-1β shRNA Knock Down IL-1β Expression In Vitro
3.2. IL-1β shRNA Increased Lipid Metabolism in Adipocytes
3.3. Construction of IL-1β shRNA/Yeast Microcapsule and Its Functional Verification on Macrophage
3.4. Yeast Microcapsule Protected shRNA from the Degradation in Gastrointestinal Tract
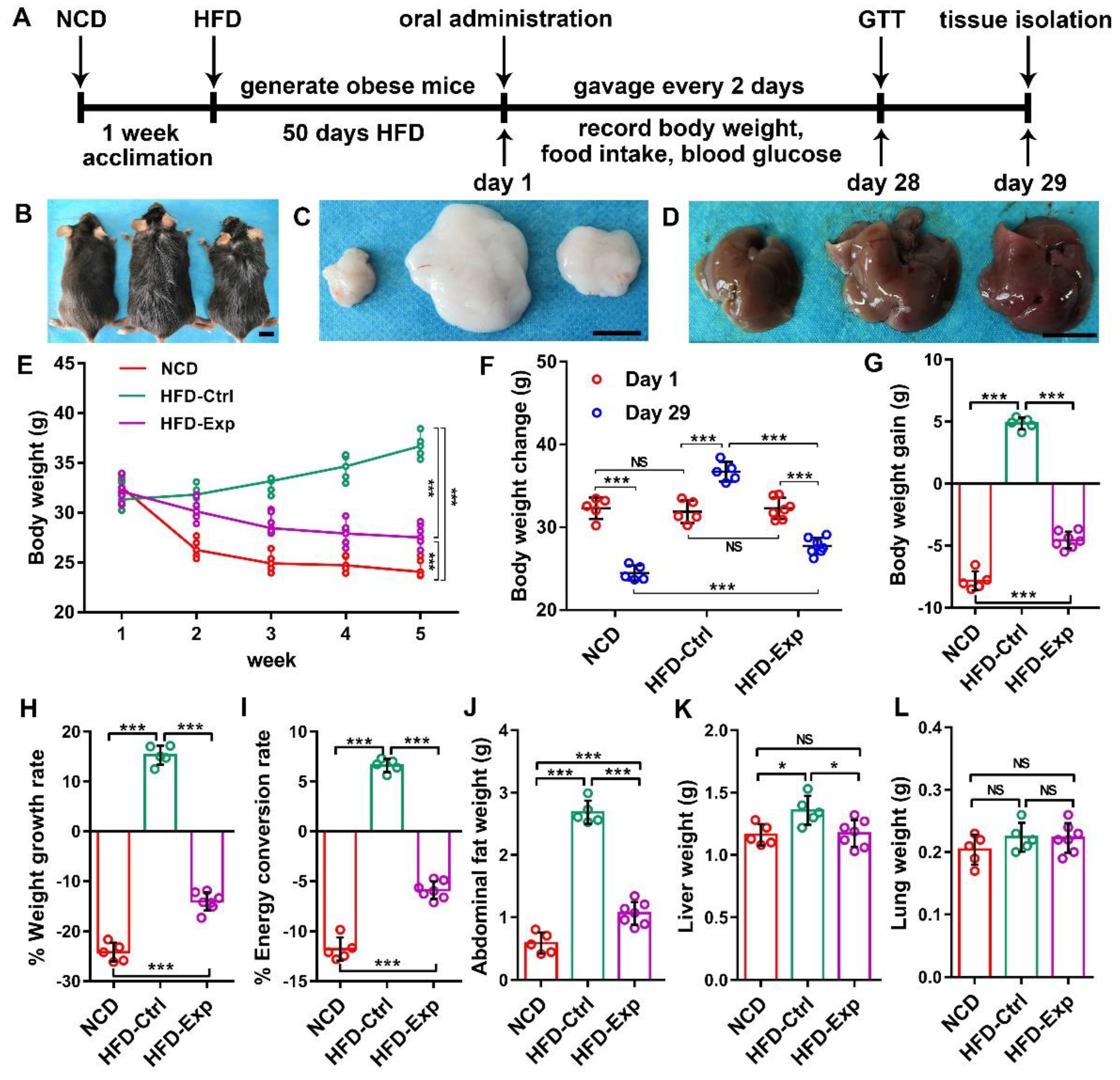
3.5. IL-1β shRNA/Yeast Regulated Body Weight via Oral Route In Vivo
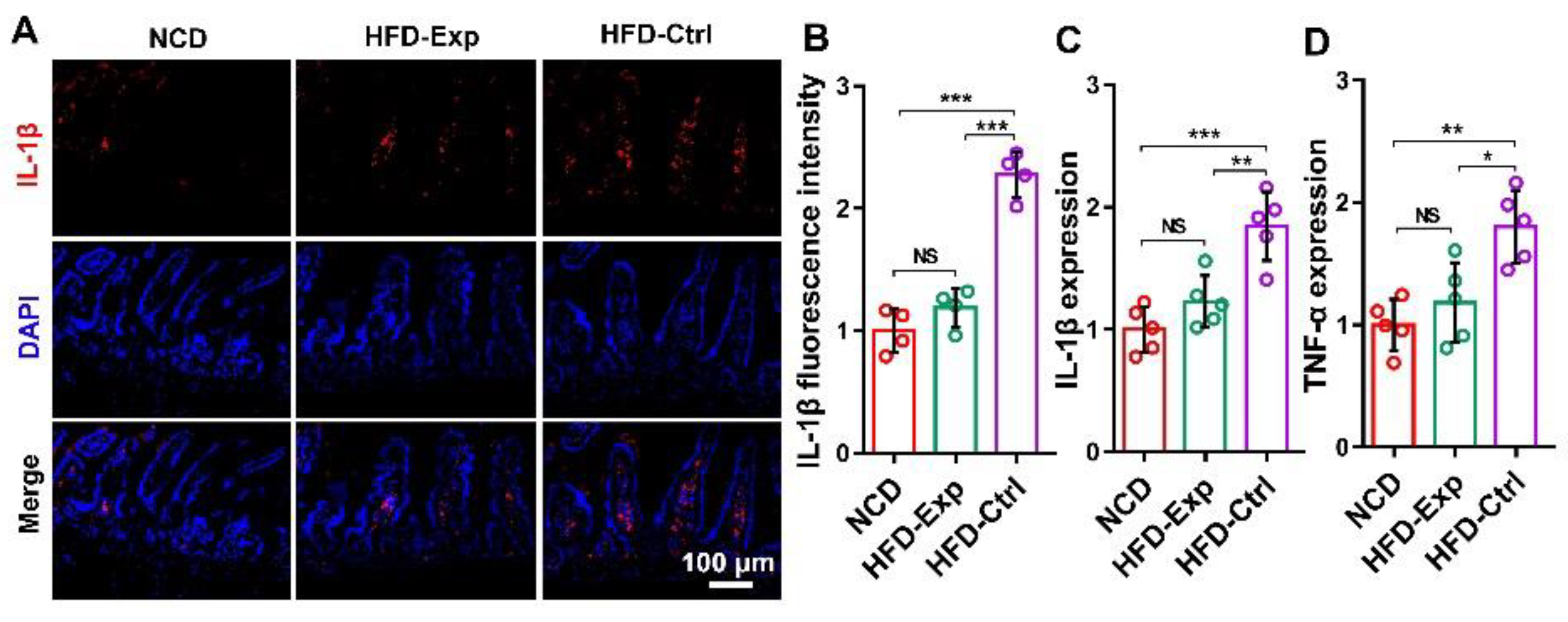
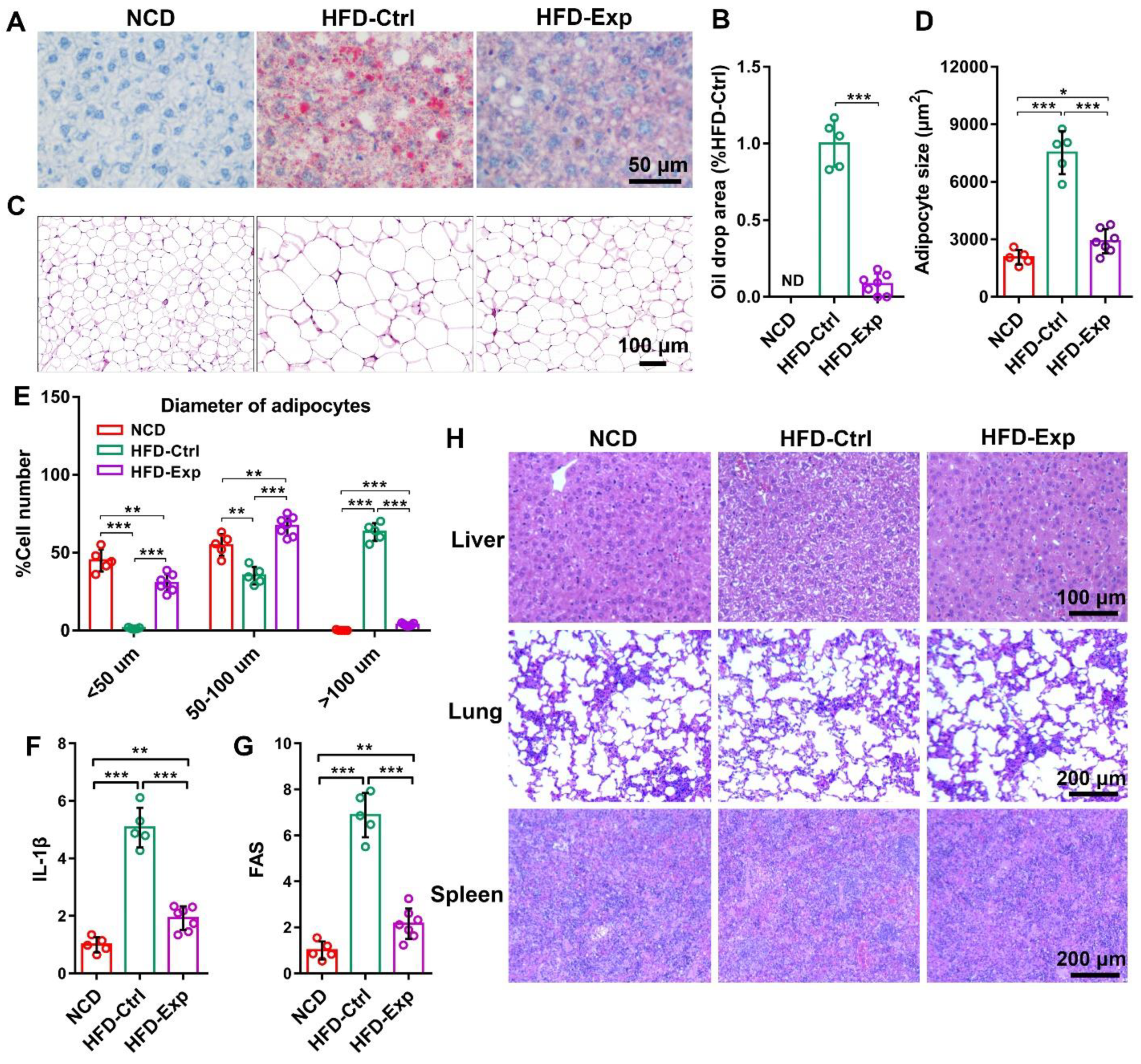
3.6. Histological Staining Proved the Effect of IL-1β shRNA/Yeast in Anti-Obesity
3.7. Biological Safety of shRNA/Yeast Microcapsule In Vivo
3.8. IL-1β shRNA/Yeast Regulated Blood Glucose Concentration and Cytokines Expression in Serum
3.9. Weight Loss Alleviates Articular Cartilage Injury
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Padwal, R.S.; Majumdar, S.R. Drug treatments for obesity: Orlistat, sibutramine, and rimonabant. Lancet 2007, 369, 71–77. [Google Scholar] [CrossRef]
- Bray, G.A.; Kim, K.K.; Wilding, J.P.H. World Obesity Federation. Obesity: A chronic relapsing progressive disease process. A position statement of the World Obesity Federation. Obes. Rev. 2017, 18, 715–723. [Google Scholar] [CrossRef] [Green Version]
- Elagizi, A.; Kachur, S.; Lavie, C.J.; Carbone, S.; Pandey, A.; Ortega, F.B.; Milani, R.V. An Overview and Update on Obesity and the Obesity Paradox in Cardiovascular Diseases. Prog. Cardiovasc. Dis. 2018, 61, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Sonnenburg, J.L.; Bäckhed, F. Diet–microbiota interactions as moderators of human metabolism. Nat. Cell Biol. 2016, 535, 56–64. [Google Scholar] [CrossRef]
- Maedler, K.; Dharmadhikari, G.; Schumann, D.; Størling, J. Interleukin-1 beta targeted therapy for type 2 diabetes. Expert Opin. Biol. Ther. 2009, 9, 1177–1188. [Google Scholar] [CrossRef] [PubMed]
- Dror, E.; Dalmas, E.; Zeman-Meier, D.; Wueest, S.; Thévenet, J.; Thienel, C.; Timper, K.; Nordmann, T.M.; Traub, S.; Schulze, F.; et al. Postprandial macrophage-derived IL-1β stimulates insulin, and both synergistically promote glucose disposal and inflammation. Nat. Immunol. 2017, 18, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Andrade-Oliveira, V.; Câmara, N.O.S.; Moraes-Vieira, P.M. Adipokines as Drug Targets in Diabetes and Underlying Disturbances. J. Diabetes Res. 2015, 2015, 681612. [Google Scholar] [CrossRef]
- High, K.A.; Roncarolo, M.G. Gene therapy. N. Engl. J. Med. 2019, 381, 455–464. [Google Scholar] [CrossRef]
- Gu, J.; Fang, X.; Hao, J.; Sha, X. Reversal of P-glycoprotein-mediated multidrug resistance by CD44 antibody-targeted nanocomplexes for short hairpin RNA-encoding plasmid DNA delivery. Biomaterials 2015, 45, 99–114. [Google Scholar] [CrossRef]
- Escribano-Viana, R.; Portu, J.; Garijo, P.; López, R.; Santamaría, P.; López-Alfaro, I.; Gutiérrez, A.R.; González-Arenzana, L. Effect of the Sequential Inoculation of Non-Saccharomyces/Saccharomyces on the Anthocyans and Stilbenes Composition of Tempranillo Wines. Front. Microbiol. 2019, 10, 773. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhang, T.; Wang, L.; Shao, S.; Chen, Z.; Zhang, Z. In vivo targeted delivery of CD40 shRNA to mouse intestinal dendritic cells by oral administration of recombinant Sacchromyces cerevisiae. Gene Ther. 2014, 21, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, X.; Han, S.; Dou, Y.; Liu, M.; Zhang, L.; Guo, J.; Shi, Q.; Gong, G.; Wang, R.; et al. Yeast microcapsule-mediated targeted delivery of diverse nanoparticles for imaging and therapy via the oral route. Nano Lett. 2017, 17, 1056–1064. [Google Scholar] [CrossRef]
- Sabu, C.; Mufeedha, P.; Pramod, K. Yeast-inspired drug delivery: Biotechnology meets bioengineering and synthetic biology. Expert Opin. Drug Deliv. 2019, 16, 27–41. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, X.; Chen, Y.; Dou, Y.; Zhou, X.; Li, L.; Li, C.; An, H.; Tao, H.; Hu, H.; et al. Bioinspired yeast microcapsules loaded with self-assembled nanotherapies for targeted treatment of cardiovascular disease. Mater. Today 2017, 20, 301–313. [Google Scholar] [CrossRef]
- Aouadi, M.; Tesz, G.J.; Nicoloro, S.M.; Wang, M.; Chouinard, M.; Soto, E.; Ostroff, G.R.; Czech, M.P. Orally delivered siRNA targeting macrophage Map4k4 suppresses systemic inflammation. Nat. Cell Biol. 2009, 458, 1180–1184. [Google Scholar] [CrossRef] [Green Version]
- Trevaskis, N.; Kaminskas, L.; Porter, C. From sewer to saviour—Targeting the lymphatic system to promote drug exposure and activity. Nat. Rev. Drug Discov. 2015, 14, 781–803. [Google Scholar] [CrossRef]
- Pabst, O.; Mowat, A.M. Oral tolerance to food protein. Mucosal Immunol. 2012, 5, 232–239. [Google Scholar] [CrossRef]
- Zhang, L.; Peng, H.; Feng, M.; Zhang, W.; Li, Y. Yeast microcapsule-mediated oral delivery of IL-1β shRNA for post-traumatic osteoarthritis therapy. Mol. Ther. Nucleic Acids 2021, 23, 336–346. [Google Scholar] [CrossRef]
- Zhang, L.; Peng, H.; Zhang, W.; Li, Y.; Liu, L.; Leng, T. Yeast Cell wall Particle mediated Nanotube-RNA delivery system loaded with miR365 Antagomir for Post-traumatic Osteoarthritis Therapy via Oral Route. Theranostics 2020, 10, 8479–8493. [Google Scholar] [CrossRef]
- Zhao, J.; Luo, D.; Zhang, Z.; Fan, N.; Wang, Y.; Nie, H.; Rong, J. Celastrol-loaded PEG-PCL nanomicelles ameliorate inflammation, lipid accumulation, insulin resistance and gastrointestinal injury in diet-induced obese mice. J. Control. Release 2019, 310, 188–197. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, M.; Chen, M.; Niu, W.; Liu, W.; Leng, T.; Ji, W.; Lei, B. A safe and efficient bioactive citrate-lysine/miRNA33 agonist nanosystem for high fat diet-induced obesity therapy. Chem. Eng. J. 2021, 408, 127304. [Google Scholar] [CrossRef]
- Mobasheri, A.; Rayman, M.P.; Gualillo, O.; Sellam, J.; van der Kraan, P.; Fearon, U. The role of metabolism in the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2017, 13, 302–311. [Google Scholar] [CrossRef]
- Lopes, E.B.P.; Filiberti, A.; Husain, S.A.; Humphrey, M.B. Immune Contributions to Osteoarthritis. Curr. Osteoporos. Rep. 2017, 15, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Li, X.; Yu, X.; Liu, X.; Xu, X.; He, J.; Gu, H.; Liu, L. Oral Administration of Resveratrol Alleviates Osteoarthritis Pathology in C57BL/6J Mice Model Induced by a High-Fat Diet. Mediat. Inflamm. 2017, 2017, 7659023. [Google Scholar] [CrossRef] [PubMed]
- Datta, P.; Zhang, Y.; Parousis, A.; Sharma, A.; Rossomacha, E.; Endisha, H.; Wu, B.; Kacprzak, I.; Mahomed, N.N.; Gandhi, R.; et al. High-fat diet-induced acceleration of osteoarthritis is associated with a distinct and sustained plasma metabolite signature. Sci. Rep. 2017, 7, 8205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbett, J.A.; Wang, J.L.; Sweetland, M.A.; Lancaster, J.R., Jr.; McDaniel, M.L. Interleukin 1 beta induces the formation of nitric oxide by beta-cells purified from rodent islets of Langerhans. Evidence for the beta-cell as a source and site of action of nitric oxide. J. Clin. Investig. 1992, 90, 2384–2391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzusoff, A.; Duke, R.C.; King, T.H.; Lu, Y.; Rodell, T.C. Yeasts encoding tumour antigens in cancer immunotherapy. Expert Opin. Biol. Ther. 2005, 5, 565–575. [Google Scholar] [CrossRef]
- Cohn, A.; Morse, M.A.; O’Neil, B.; Bellgrau, D.; Duke, R.C.; Franzusoff, A.J.; Munson, S.; Ferraro, J.; Rodell, T.C. Treatment of Ras mutation-bearing solid tumors using whole recombinant S. cerevisiae yeast expressing mutated Ras: Preliminary safety and immunogenicity results from a phase 1 trial. J. Clin. Oncol. 2005, 23, 2571. [Google Scholar] [CrossRef]
- Baldassano, S.; Accardi, G.; Vasto, S. Beta-glucans and cancer: The influence of inflammation and gut peptide. Eur. J. Med. Chem. 2017, 142, 486–492. [Google Scholar] [CrossRef]
- Walachowski, S.; Tabouret, G.; Fabre, M.; Foucras, G. Molecular Analysis of a Short-term Model of β-Glucans-Trained Immunity Highlights the Accessory Contribution of GM-CSF in Priming Mouse Macrophages Response. Front. Immunol. 2017, 8, 1089. [Google Scholar] [CrossRef] [Green Version]
- Price, N.L.; Singh, A.K.; Rotllan, N.; Goedeke, L.; Wing, A.; Canfrán-Duque, A.; Diaz-Ruiz, A.; Araldi, E.; Baldán, Á.; Camporez, J.-P.; et al. Genetic ablation of miR-33 increases food intake, enhances adipose tissue expansion, and promotes obesity and insulin resistance. Cell Rep. 2018, 22, 2133–2145. [Google Scholar] [CrossRef] [Green Version]
- Könner, A.C.; Brüning, J.C. Selective insulin and leptin resistance in metabolic disorders. Cell Metab. 2012, 16, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Myers, M.G., Jr.; Leibel, R.L.; Seeley, R.J.; Schwartz, M.W. Obesity and leptin resistance: Distinguishing cause from effect. Trends Endocrinol. Metab. 2010, 21, 643–651. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, T.; Kadowaki, T. Adiponectin Receptor as a Key Player in Healthy Longevity and Obesity-Related Diseases. Cell Metab. 2013, 17, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Wojdasiewicz, P.; Poniatowski, Ł.A.; Szukiewicz, D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediat. Inflamm. 2014, 2, 561459. [Google Scholar] [CrossRef] [Green Version]
- Che, C.; Liu, J.; Ma, L.; Xu, H.; Bai, N.; Zhang, Q. LOX-1 is involved in IL-1β production and extracellular matrix breakdown in dental peri-implantitis. Int. Immunopharmacol. 2017, 52, 127–135. [Google Scholar] [CrossRef]
- Chen, J.; Wu, W.; Zhang, M.; Chen, C. Taraxasterol suppresses inflammation in IL-1β-induced rheumatoid arthritis fibroblast-like synoviocytes and rheumatoid arthritis progression in mice. Int. Immunopharmacol. 2019, 70, 274–283. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′ to 3′) |
|---|---|
| shDec-F | AGAATCGTTGCCTGCACATC |
| shDec-R | GAGATAGCAAGGTATTCAG |
| IL-1β-F | AACCTGCTGGTGTGTGACGTTC |
| IL-1β-R | CAGCACGAGGCTTTTTTGTTGT |
| 18S-F | CGGCTACCACATCCAAGGAA |
| 18S-R | GCTGGAATTACCGCGGCT |
| FAS-F | CTGAGATCCCAGCACTTCTTGA |
| FAS-R | GCCTCCGAAGCCAAATGAG |
| TNF-α-F | ACCCTCACACTCAGATCATCTTC |
| TNF-α-R | TGGTGGTTTGCTACGACGT |
| MHC-F | GGAAGACCGCAAGAATGTTCTC |
| MHC-R | CAGGTTGACATTGGATTGTTCCT |
| MSTN-F | CAAAACACGAGGTACTCCAGAATAG |
| MSTN-R | CCGTGGTAGCGTGATAATCGT |
| Col II-F | AGGACCTGGTAGTGCGAGTGAC |
| Col II-R | CTGGGCGATAGTGGAATACAACT |
| Col X-F | GATGCCGCTTGTCAGTGCT |
| Col X-R | CCAGATCTTGGGTCGTAATGC |
| MMP13-F | GCCCCTTCCCTATGGTGATG |
| MMP13-R | TTGCATTTCTCGGAGCCTGT |
| Week 1 | Week 2 | Week 3 | Week 4 | Week 5 | |
|---|---|---|---|---|---|
| NCD (g) | 32.30 ± 1.29 | 26.39 ± 0.93 | 25.02 ± 0.94 | 24.87 ± 0.77 | 24.47 ± 0.91 |
| HFD-Ctrl (g) | 31.87 ± 1.36 | 31.86 ± 0.92 | 32.80 ± 0.76 | 34.54 ± 1.24 | 36.72 ± 1.18 |
| HFD-Exp (g) | 32.82 ± 1.28 | 30.37 ± 1.09 | 28.86 ± 0.98 | 27.83 ± 1.10 | 27.73 ± 1.01 |
| Total Intake (g) | Average Net Intake Per Mouse (g) | Net Gain Per Mouse (g) | Energy Conversion Rate (%) | |
|---|---|---|---|---|
| NCD (n = 5) | 332.12 | 66.42 | −7.82 ± 0.77 | −11.78 ± 1.16 |
| HFD-Ctrl (n = 5) | 367.21 | 73.44 | 4.85 ± 0.48 | 6.60 ± 0.65 |
| HFD-Exp (n = 7) | 538.83 | 76.98 | −4.55 ± 0.67 | −5.91 ± 0.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Zhang, W.; Peng, H.; Li, Y.; Leng, T.; Xie, C.; Zhang, L. Oral Gene Therapy of HFD-Obesity via Nonpathogenic Yeast Microcapsules Mediated shRNA Delivery. Pharmaceutics 2021, 13, 1536. https://doi.org/10.3390/pharmaceutics13101536
Zhang L, Zhang W, Peng H, Li Y, Leng T, Xie C, Zhang L. Oral Gene Therapy of HFD-Obesity via Nonpathogenic Yeast Microcapsules Mediated shRNA Delivery. Pharmaceutics. 2021; 13(10):1536. https://doi.org/10.3390/pharmaceutics13101536
Chicago/Turabian StyleZhang, Li, Wan Zhang, Hang Peng, Yankun Li, Tongtong Leng, Chenxi Xie, and Long Zhang. 2021. "Oral Gene Therapy of HFD-Obesity via Nonpathogenic Yeast Microcapsules Mediated shRNA Delivery" Pharmaceutics 13, no. 10: 1536. https://doi.org/10.3390/pharmaceutics13101536
APA StyleZhang, L., Zhang, W., Peng, H., Li, Y., Leng, T., Xie, C., & Zhang, L. (2021). Oral Gene Therapy of HFD-Obesity via Nonpathogenic Yeast Microcapsules Mediated shRNA Delivery. Pharmaceutics, 13(10), 1536. https://doi.org/10.3390/pharmaceutics13101536