
Albumin-EDTA-Vanadium Is a Powerful Anti-Proliferative Agent, Following Entrance into Glioma Cells via Caveolae-Mediated Endocytosis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Mercaptoalbumin
2.3. Preparation of EDTA-NH-(CH2)2-Maleimide (EDTA-Maleimide)
2.4. Preparation of HSA-S-MAL-EDTA
2.5. Characterization of HSA-S-MAL-(CH2)2-NHCO-EDTA by Reversed Phase Liquid Chromatography, Coupled to Mass Spectrometry (LC-MS) Analysis
2.6. Evaluating the Affinities of HSA-EDTA to Vanadium: Reversal of Vanadium-Evoked Inhibition of Acid Phosphatase
2.7. Preparation of HSA-EDTA-Vanadium
2.8. Preparation of Rhodamine-Labeled HSA-EDTA
2.9. Growth Inhibitory Effects of EDTA and Vanadium Containing Conjugates
2.10. Immunocytochemistry
2.11. Uptake of Rhodamine-HSA and Rhodamine-HSA-EDTA-VO++
2.12. Statistical Analysis
3. Results
3.1. Preparation of Monomodified HSA-EDTA Derivative
3.2. Characterization of HSA-S-MAL-(CH2)2-NHCO-EDTA by LC-MS
3.3. Association of Vanadium with HSA-EDTA: Comparison to EDTA and EDTA-Maleimide
3.4. Preparation of HSA-EDTA Vanadium Conjugates
3.5. Stability of HSA-EDTA-VO++ as a Function of pH
3.6. HSA-EDTA-Vanadium Conjugates Are Powerful Anti-Proliferative Agents
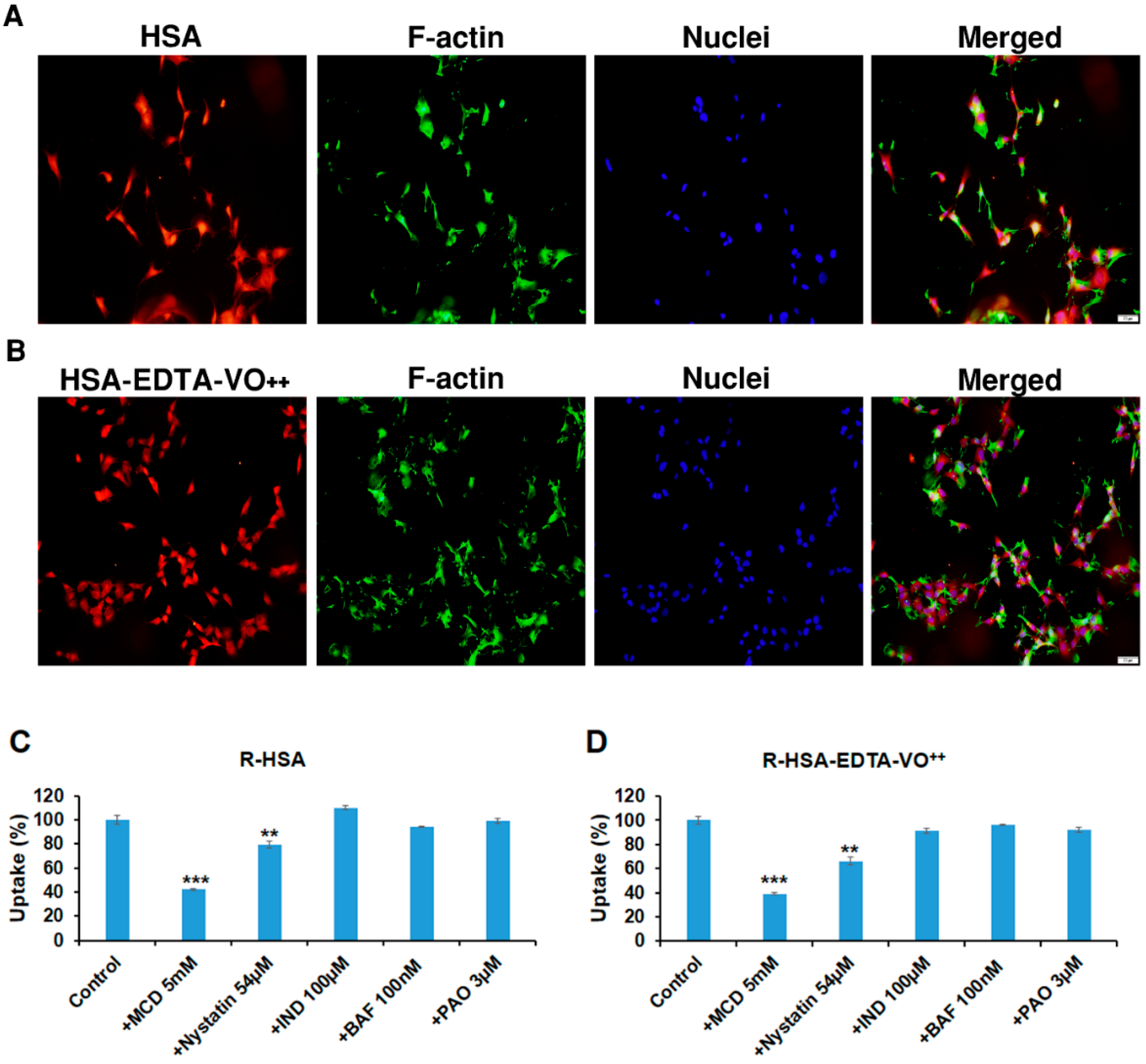
3.7. HSA-EDTA-VO++ Penetrates into CNS-1 Glioma Cell Line via Caveolae-Mediated Endocytosis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Elberg, G.; Li, J.; Shechter, Y. Vanadium activates or inhibits receptor and non-receptor protein tyrosine kinases in cell-free experiments, depending on its oxidation state. Possible role of endogenous vanadium in controlling cellular protein tyrosine kinase activity. J. Biol. Chem. 1994, 269, 9521–9527. [Google Scholar] [CrossRef]
- Meyerovitch, J.; Farfel, Z.; Sack, J.; Shechter, Y. Oral administration of vanadate normalizes blood glucose levels in streptozotocin-treated rats. Characterization and mode of action. J. Biol. Chem. 1987, 262, 6658–6662. [Google Scholar] [CrossRef]
- Shechter, Y.; Karlish, S.J. Insulin-like stimulation of glucose oxidation in rat adipocytes by vanadyl (IV) ions. Nature 1980, 284, 556–558. [Google Scholar] [CrossRef]
- Goldwaser, I.; Gefel, D.; Gershonov, E.; Fridkin, M.; Shechter, Y. Insulin-like effects of vanadium: Basic and clinical implications. J. Inorg. Biochem. 2000, 80, 21–25. [Google Scholar] [CrossRef]
- Sekar, N.; Li, J.; Shechter, Y. Vanadium salts as insulin substitutes: Mechanisms of action, a scientific and therapeutic tool in diabetes mellitus research. Crit. Rev. Biochem. Mol. Biol. 1996, 31, 339–359. [Google Scholar] [CrossRef]
- Jungwirth, U.; Kowol, C.R.; Keppler, B.K.; Hartinger, C.G.; Berger, W.; Heffeter, P. Anticancer activity of metal complexes: Involvement of redox processes. Antioxid. Redox Signal. 2011, 15, 1085–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tracey, A.S.; Willsky, G.R.; Takeuchi, E.S. Vanadium: Chemistry, Biochemistry, Pharmacological, and Practical Applications; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2007. [Google Scholar]
- Ścibior, A.; Pietrzyk, Ł.; Plewa, Z.; Skiba, A. Vanadium: Risks and possible benefits in the light of a comprehensive overview of its pharmacotoxicological mechanisms and multi-applications with a summary of further research trends. J. Trace Elem. Med. Biol. 2020, 61, 126508. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A.; Waghray, A.; Patel, M.A.; Chatterjee, M. Vanadium in the detection, prevention and treatment of cancer: The in vivo evidence. Cancer Lett. 2010, 294, 1–12. [Google Scholar] [CrossRef]
- Chakraborty, A.; Ghosh, R.; Roy, K.; Ghosh, S.; Chowdhury, P.; Chatterjee, M. Vanadium: A modifier of drug-metabolizing enzyme patterns and its critical role in cellular proliferation in transplantable murine lymphoma. Oncology 1995, 52, 310–314. [Google Scholar] [CrossRef]
- El-Naggar, M.M.; El-Waseef, A.M.; El-Halafawy, K.M.; El-Sayed, I.H. Antitumor activities of vanadium(IV), manganese(IV), iron(III), cobalt(II) and copper(II) complexes of 2-methylaminopyridine. Cancer Lett. 1998, 133, 71–76. [Google Scholar] [CrossRef]
- Evangelou, A.M. Vanadium in cancer treatment. Crit. Rev. Oncol. Hematol. 2002, 42, 249–265. [Google Scholar] [CrossRef]
- Kostova, I. Titanium and vanadium complexes as anticancer agents. Anticancer Agents Med. Chem. 2009, 9, 827–842. [Google Scholar] [CrossRef] [PubMed]
- Leon, I.E.; Cadavid-Vargas, J.F.; Di Virgilio, A.L.; Etcheverry, S.B. Vanadium, Ruthenium and Copper Compounds: A New Class of Nonplatinum Metallodrugs with Anticancer Activity. Curr. Med. Chem. 2017, 24, 112–148. [Google Scholar] [CrossRef] [PubMed]
- Neumann, E.; Frei, E.; Funk, D.; Becker, M.D.; Schrenk, H.-H.; Müller-Ladner, U.; Fiehn, C. Native albumin for targeted drug delivery. Expert Opin. Drug Deliv. 2010, 7, 915–925. [Google Scholar] [CrossRef]
- Larsen, M.T.; Kuhlmann, M.; Hvam, M.L.; Howard, K.A. Albumin-based drug delivery: Harnessing nature to cure disease. Mol. Cell. Ther. 2016, 4, 3. [Google Scholar] [CrossRef] [Green Version]
- Crans, D.C.; Bunch, R.L.; Theisen, L.A. Interaction of trace levels of vanadium(IV) and vanadium(V) in biological systems. J. Am. Chem. Soc. 1989, 111, 7597–7607. [Google Scholar] [CrossRef]
- Sogami, M.; Nagoka, S.; Era, S.; Honda, M.; Noguchi, K. Resolution of human mercapt- and nonmercaptalbumin by high-performance liquid chromatography. Int. J. Pept. Protein Res. 1984, 24, 96–103. [Google Scholar] [CrossRef]
- Hartley, R.W.; Peterson, E.A.; Sober, H.A. The relation of free sulfhydryl groups to chromatographic heterogeneity and polymerization of bovine plasma albumin. Biochemistry 1962, 1, 60–68. [Google Scholar] [CrossRef]
- Grassetti, D.R.; Murray, J.F. Determination of sulfhydryl groups with 2,2′- or 4,4′-dithiodipyridine. Arch. Biochem. Biophys. 1967, 119, 41–49. [Google Scholar] [CrossRef]
- Rozen, S.; Tieri, A.; Ridner, G.; Stark, A.-K.; Schmaler, T.; Ben-Nissan, G.; Dubiel, W.; Sharon, M. Exposing the subunit diversity within protein complexes: A mass spectrometry approach. Methods 2013, 59, 270–277. [Google Scholar] [CrossRef]
- Marty, M.T.; Baldwin, A.J.; Marklund, E.G.; Hochberg, G.K.A.; Benesch, J.L.P.; Robinson, C.V. Bayesian deconvolution of mass and ion mobility spectra: From binary interactions to polydisperse ensembles. Anal. Chem. 2015, 87, 4370–4376. [Google Scholar] [CrossRef] [Green Version]
- Kruse, C.A.; Molleston, M.C.; Parks, E.P.; Schiltz, P.M.; Kleinschmidt-DeMasters, B.K.; Hickey, W.F. A rat glioma model, CNS-1, with invasive characteristics similar to those of human gliomas: A comparison to 9L gliosarcoma. J. Neurooncol. 1994, 22, 191–200. [Google Scholar] [CrossRef]
- Cecchelli, R.; Aday, S.; Sevin, E.; Almeida, C.; Culot, M.; Dehouck, L.; Coisne, C.; Engelhardt, B.; Dehouck, M.-P.; Ferreira, L. A stable and reproducible human blood-brain barrier model derived from hematopoietic stem cells. PLoS ONE 2014, 9, e99733. [Google Scholar]
- Shelly, S.; Liraz Zaltsman, S.; Ben-Gal, O.; Dayan, A.; Ganmore, I.; Shemesh, C.; Atrakchi, D.; Garra, S.; Ravid, O.; Rand, D.; et al. Potential neurotoxicity of titanium implants: Prospective, in-vivo and in-vitro study. Biomaterials 2021, 276, 121039. [Google Scholar] [CrossRef]
- Israelov, H.; Ravid, O.; Atrakchi, D.; Rand, D.; Elhaik, S.; Bresler, Y.; Twitto-Greenberg, R.; Omesi, L.; Liraz-Zaltsman, S.; Gosselet, F.; et al. Caspase-1 has a critical role in blood-brain barrier injury and its inhibition contributes to multifaceted repair. J. Neuroinflamma 2020, 17, 267. [Google Scholar] [CrossRef]
- Cooper, I.; Fridkin, M.; Shechter, Y. Conjugation of Methotrexate-Amino Derivatives to Macromolecules through Carboxylate Moieties Is Superior Over Conventional Linkage to Amino Residues: Chemical, Cell-Free and In Vitro Characterizations. PLoS ONE 2016, 11, e0158352. [Google Scholar] [CrossRef]
- Hazum, E.; Shisheva, A.; Shechter, Y. Preparation and application of radioiodinated sulfhydryl reagents for the covalent labeling of SH-proteins present in minute quantities. J. Biochem. Biophys. Methods 1992, 24, 95–106. [Google Scholar] [CrossRef]
- Butler, P.J.; Harris, J.I.; Hartley, B.S.; Lebeman, R. The use of maleic anhydride for the reversible blocking of amino groups in polypeptide chains. Biochem. J. 1969, 112, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Visser, C.C.; Stevanović, S.; Heleen Voorwinden, L.; Gaillard, P.J.; Crommelin, D.J.A.; Danhof, M.; De Boer, A.G. Validation of the transferrin receptor for drug targeting to brain capillary endothelial cells in vitro. J. Drug Target 2004, 12, 145–150. [Google Scholar] [CrossRef]
- Shechter, Y.; Eldberg, G.; Shisheva, A.; Gefel, D.; Sekar, N.; Qian, S.; Bruck, R.; Gershonov, E.; Crans, D.C.; Goldwasser, Y.; et al. Insulin-like Effects of Vanadium; Reviewing In Vivo and In Vitro Studies and Mechanisms of Action. In Vanadium Compounds: Chemistry, Biochemistry, and Therapeutic Applications; Tracey, A.S., Crans, D.C., Eds.; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1998; Volume 711, pp. 308–315. [Google Scholar]
- Mukherjee, B.; Patra, B.; Mahapatra, S.; Banerjee, P.; Tiwari, A.; Chatterjee, M. Vanadium—An element of atypical biological significance. Toxicol. Lett. 2004, 150, 135–143. [Google Scholar] [CrossRef]
- Jain, R.K. Delivery of molecular and cellular medicine to solid tumors. J. Control. Release 1998, 53, 49–67. [Google Scholar] [CrossRef]
- Maeda, H.; Wu, J.; Sawa, T.; Matsumura, Y.; Hori, K. Tumor vascular permeability and the EPR effect in macromolecular therapeutics: A review. J. Control. Release 2000, 65, 271–284. [Google Scholar] [CrossRef]
- Carter, D.C.; Ho, J.X. Structure of serum albumin. In Lipoproteins, Apolipoproteins, and Lipases; Advances in Protein Chemistry; Elsevier: Amsterdam, The Netherlands, 1994; Volume 45, pp. 153–203. [Google Scholar]
- Kratz, F.; Warnecke, A.; Scheuermann, K.; Stockmar, C.; Schwab, J.; Lazar, P.; Drückes, P.; Esser, N.; Drevs, J.; Rognan, D.; et al. Probing the cysteine-34 position of endogenous serum albumin with thiol-binding doxorubicin derivatives. Improved efficacy of an acid-sensitive doxorubicin derivative with specific albumin-binding properties compared to that of the parent compound. J. Med. Chem. 2002, 45, 5523–5533. [Google Scholar] [CrossRef]
- Smith, S.W. The role of chelation in the treatment of other metal poisonings. J. Med. Toxicol. 2013, 9, 355–369. [Google Scholar] [CrossRef] [Green Version]
- Boden, G.; Chen, X.; Ruiz, J.; van Rossum, G.D.; Turco, S. Effects of vanadyl sulfate on carbohydrate and lipid metabolism in patients with non-insulin-dependent diabetes mellitus. Metab. Clin. Exp. 1996, 45, 1130–1135. [Google Scholar] [CrossRef]
- Cohen, N.; Halberstam, M.; Shlimovich, P.; Chang, C.J.; Shamoon, H.; Rossetti, L. Oral vanadyl sulfate improves hepatic and peripheral insulin sensitivity in patients with non-insulin-dependent diabetes mellitus. J. Clin. Investg. 1995, 95, 2501–2509. [Google Scholar] [CrossRef] [Green Version]
- Halberstam, M.; Cohen, N.; Shlimovich, P.; Rossetti, L.; Shamoon, H. Oral vanadyl sulfate improves insulin sensitivity in NIDDM but not in obese nondiabetic subjects. Diabetes 1996, 45, 659–666. [Google Scholar] [CrossRef]
- Heyliger, C.E.; Tahiliani, A.G.; McNeill, J.H. Effect of vanadate on elevated blood glucose and depressed cardiac performance of diabetic rats. Science 1985, 227, 1474–1477. [Google Scholar] [CrossRef]
- Meyerovitch, J.; Rothenberg, P.; Shechter, Y.; Bonner-Weir, S.; Kahn, C.R. Vanadate normalizes hyperglycemia in two mouse models of non-insulin-dependent diabetes mellitus. J. Clin. Investg. 1991, 87, 1286–1294. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.; Moshnikova, A.; Engelman, D.M.; Reshetnyak, Y.K.; Andreev, O.A. Probe for the measurement of cell surface pH in vivo and ex vivo. Proc. Natl. Acad. Sci. USA 2016, 113, 8177–8181. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Chen, Z.; Wang, B.; Jin, Z.; Hou, Y.; Ma, S.; Liu, X. The role of lysosome in cell death regulation. Tumour Biol. 2016, 37, 1427–1436. [Google Scholar] [CrossRef] [PubMed]
- Corbet, C.; Feron, O. Tumour acidosis: From the passenger to the driver’s seat. Nat. Rev. Cancer 2017, 17, 577–593. [Google Scholar] [CrossRef] [PubMed]
- Korenchan, D.E.; Flavell, R.R. Spatiotemporal ph heterogeneity as a promoter of cancer progression and therapeutic resistance. Cancers 2019, 11, 1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stehle, G.; Sinn, H.; Wunder, A.; Schrenk, H.H.; Schütt, S.; Maier-Borst, W.; Heene, D.L. The loading rate determines tumor targeting properties of methotrexate-albumin conjugates in rats. Anticancer Drugs 1997, 8, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Valitova, J.; Sulkarnayeva, A.; Kotlova, E.; Ponomareva, A.; Mukhitova, F.K.; Murtazina, L.; Ryzhkina, I.; Beckett, R.; Minibayeva, F. Sterol binding by methyl-β-cyclodextrin and nystatin--comparative analysis of biochemical and physiological consequences for plants. FEBS J. 2014, 281, 2051–2060. [Google Scholar] [CrossRef] [Green Version]
- Ikehata, M.; Yumoto, R.; Nakamura, K.; Nagai, J.; Takano, M. Comparison of albumin uptake in rat alveolar type II and type I-like epithelial cells in primary culture. Pharm. Res. 2008, 25, 913–922. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | IC50 (µM) 2 | % Dissociated 3 | Comments |
|---|---|---|---|
| 5.0 | 0.7 4 | 100 | Full dissociation |
| 5.4 | 1.0 | 70 | |
| 6.0 | 1.57 | 44 | |
| 7.0 | 3.7 | 19 | |
| 7.15 | 5.0 | 14 | |
| 7.31 | 8.1 4 | 9 | Relatively stable |
| Compound | IC50 (µM) 1 | Potentiation Efficacy (Fold) |
|---|---|---|
| Vanadate (NaVO3) | 5.3 | |
| Vanadyl (VOCl2) | 8.9 | |
| HSA | >20 | |
| EDTA | >20 | |
| HSA-EDTA | >10 | |
| EDTA·VO3− (1:1 complex) | 3.2 | |
| EDTA·VO+2 (1:1 complex) | 16.0 | |
| HSA-EDTA-VO3− | 0.27 | 19.6 |
| HSA-EDTA-VO+2 | 0.34 | 26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cooper, I.; Ravid, O.; Rand, D.; Atrakchi, D.; Shemesh, C.; Bresler, Y.; Ben-Nissan, G.; Sharon, M.; Fridkin, M.; Shechter, Y. Albumin-EDTA-Vanadium Is a Powerful Anti-Proliferative Agent, Following Entrance into Glioma Cells via Caveolae-Mediated Endocytosis. Pharmaceutics 2021, 13, 1557. https://doi.org/10.3390/pharmaceutics13101557
Cooper I, Ravid O, Rand D, Atrakchi D, Shemesh C, Bresler Y, Ben-Nissan G, Sharon M, Fridkin M, Shechter Y. Albumin-EDTA-Vanadium Is a Powerful Anti-Proliferative Agent, Following Entrance into Glioma Cells via Caveolae-Mediated Endocytosis. Pharmaceutics. 2021; 13(10):1557. https://doi.org/10.3390/pharmaceutics13101557
Chicago/Turabian StyleCooper, Itzik, Orly Ravid, Daniel Rand, Dana Atrakchi, Chen Shemesh, Yael Bresler, Gili Ben-Nissan, Michal Sharon, Mati Fridkin, and Yoram Shechter. 2021. "Albumin-EDTA-Vanadium Is a Powerful Anti-Proliferative Agent, Following Entrance into Glioma Cells via Caveolae-Mediated Endocytosis" Pharmaceutics 13, no. 10: 1557. https://doi.org/10.3390/pharmaceutics13101557
APA StyleCooper, I., Ravid, O., Rand, D., Atrakchi, D., Shemesh, C., Bresler, Y., Ben-Nissan, G., Sharon, M., Fridkin, M., & Shechter, Y. (2021). Albumin-EDTA-Vanadium Is a Powerful Anti-Proliferative Agent, Following Entrance into Glioma Cells via Caveolae-Mediated Endocytosis. Pharmaceutics, 13(10), 1557. https://doi.org/10.3390/pharmaceutics13101557