
Enhanced Cellular Uptake of H-Chain Human Ferritin Containing Gold Nanoparticles

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Construction
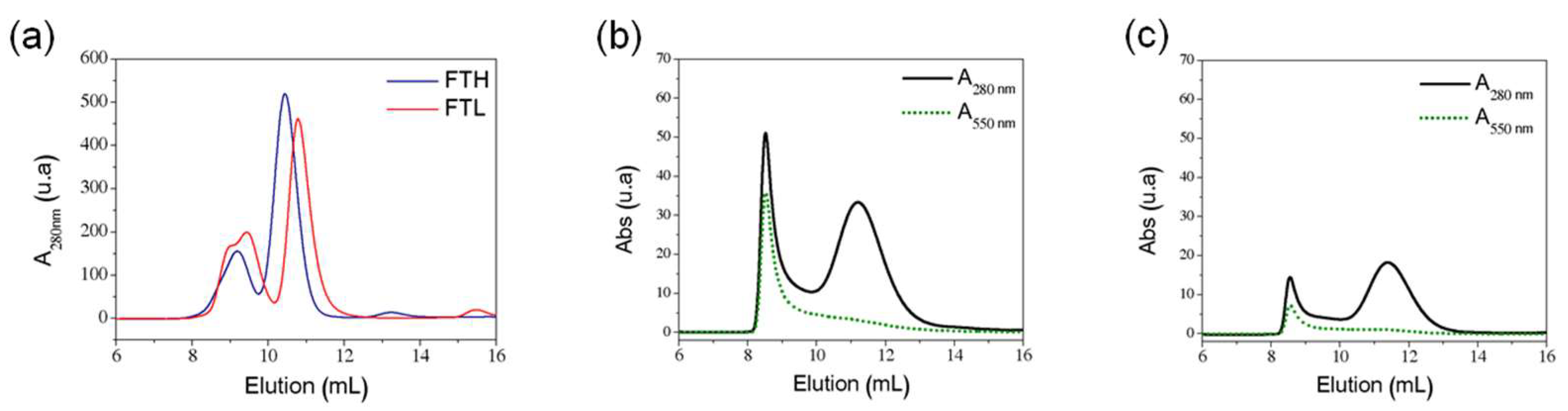
2.2. Ferritin Expression and Purification
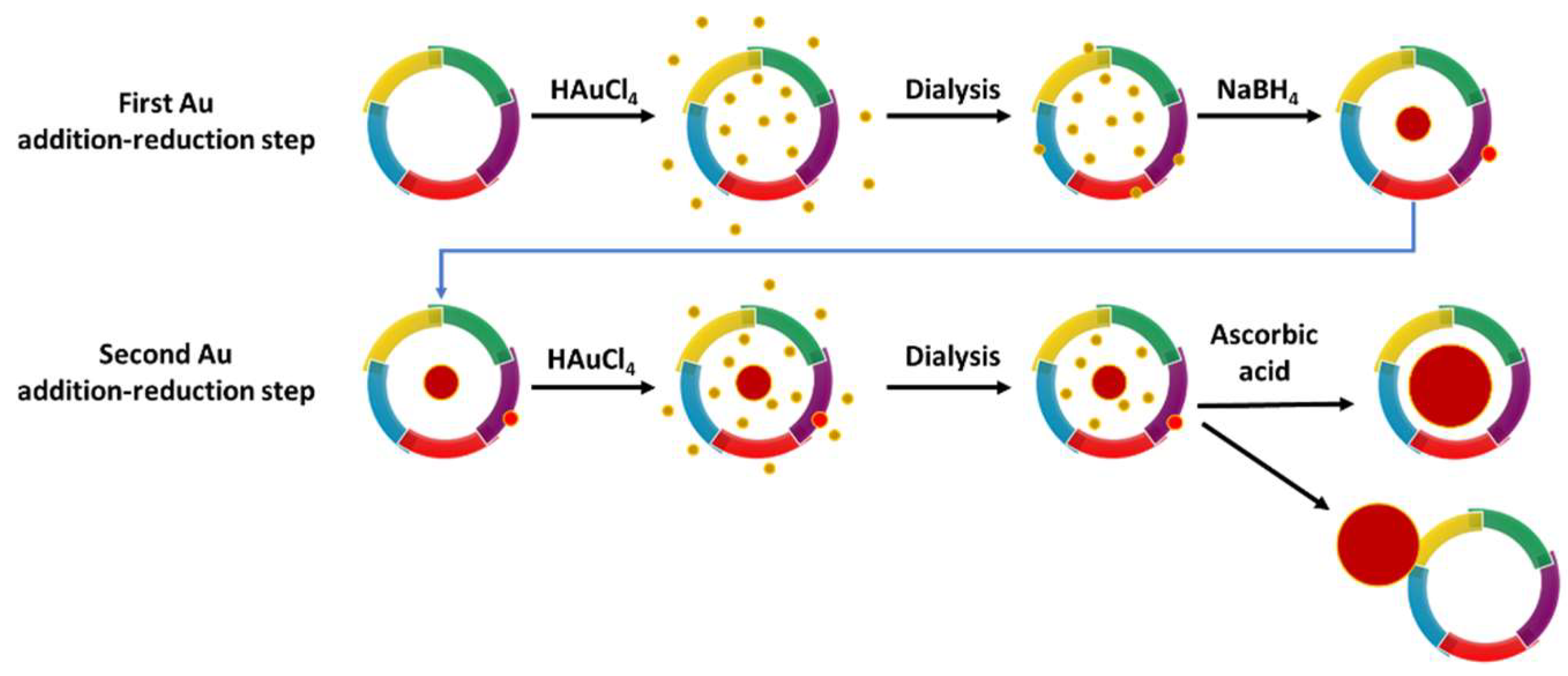
2.3. Gold Nanoparticle Synthesis in Apoferritin
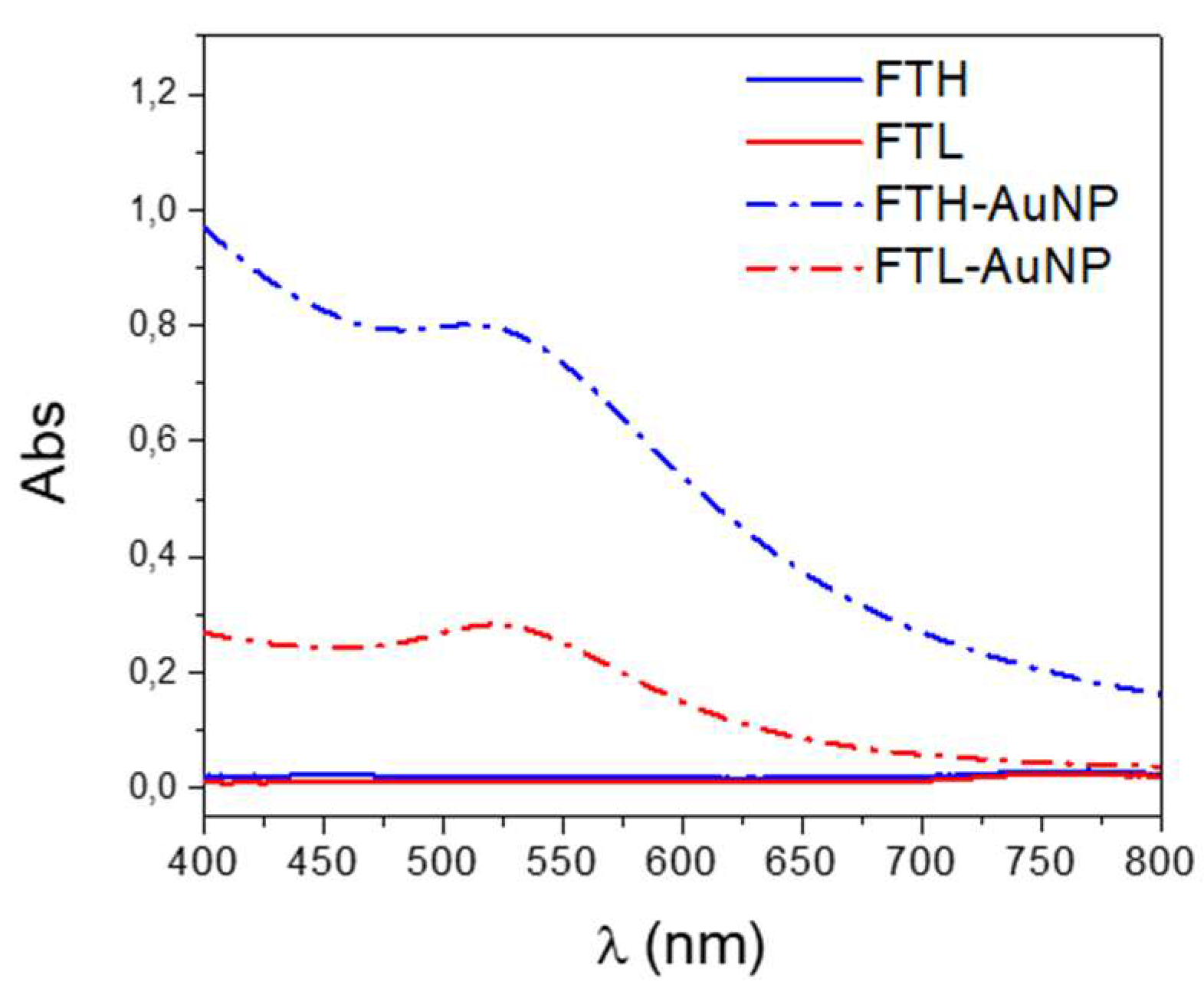
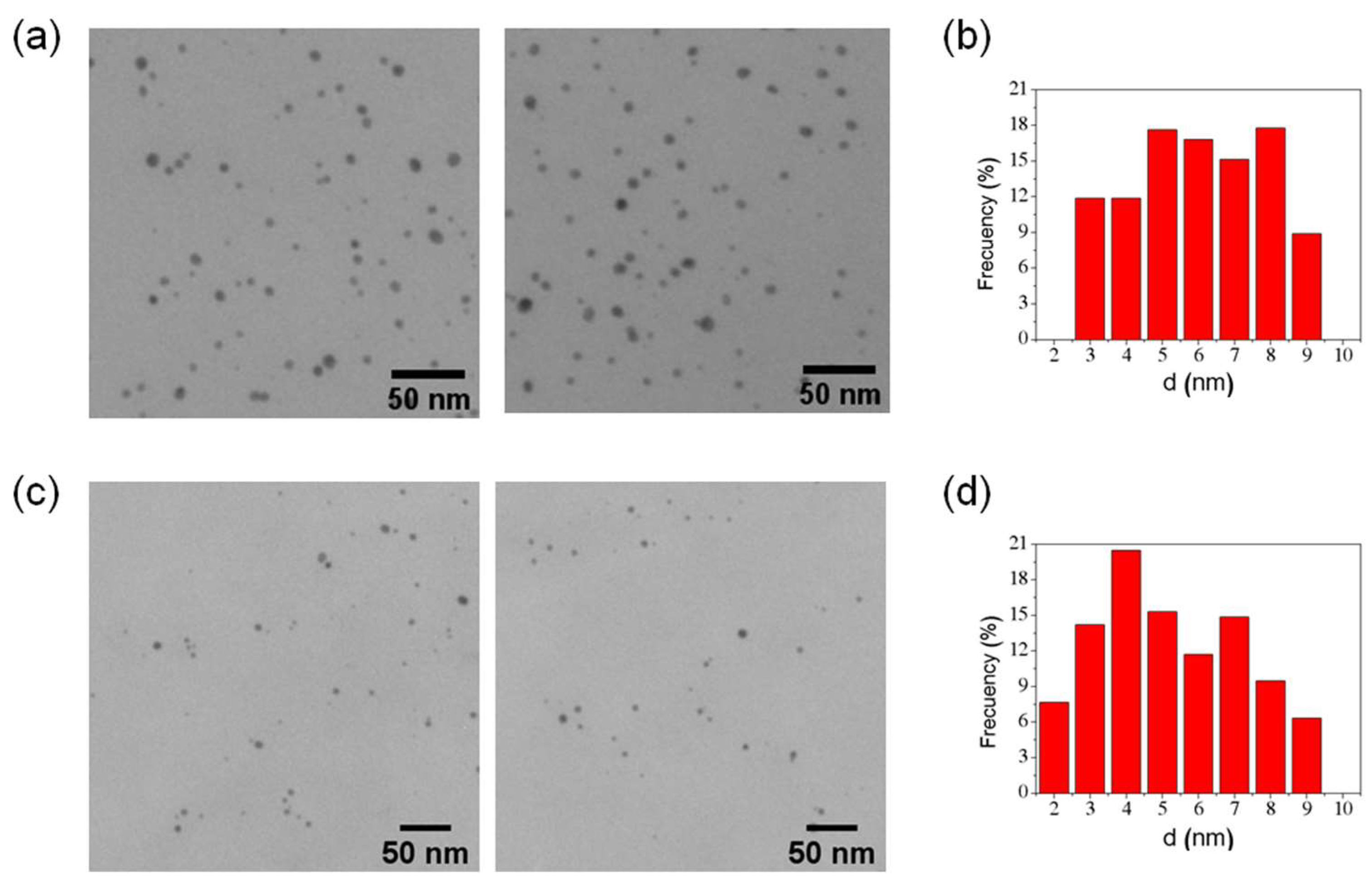
2.4. Characterization of Ferritin AuNP
2.5. Cell Culture and Incubation with Ferritin AuNP
2.6. Viability Assays
2.7. Fixation and Confocal Microscopy
3. Results and Discussion
3.1. Gold Nanoparticles Synthesis and Characterization
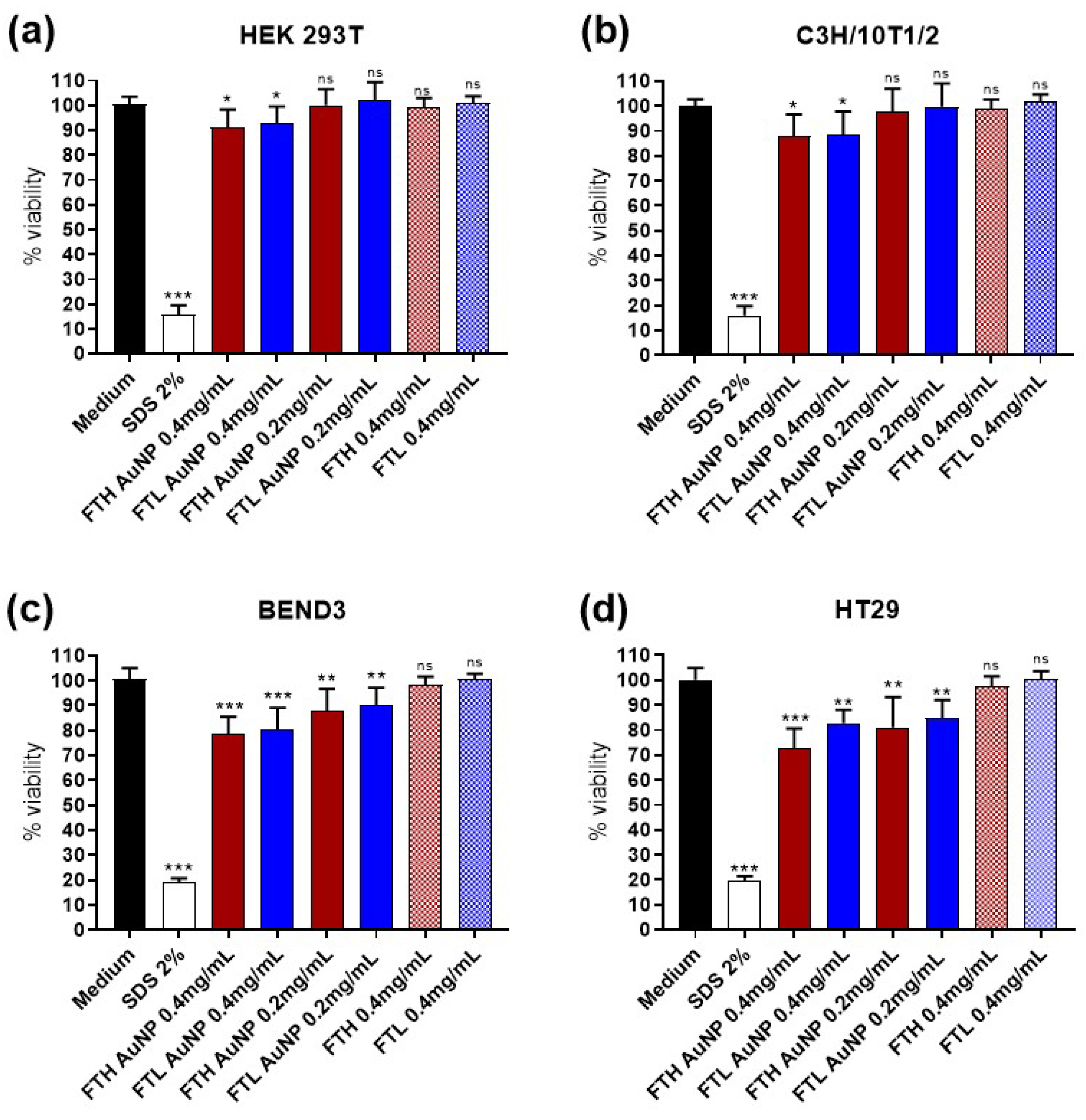
3.2. Cell Viability and FT-AuNP Toxicity
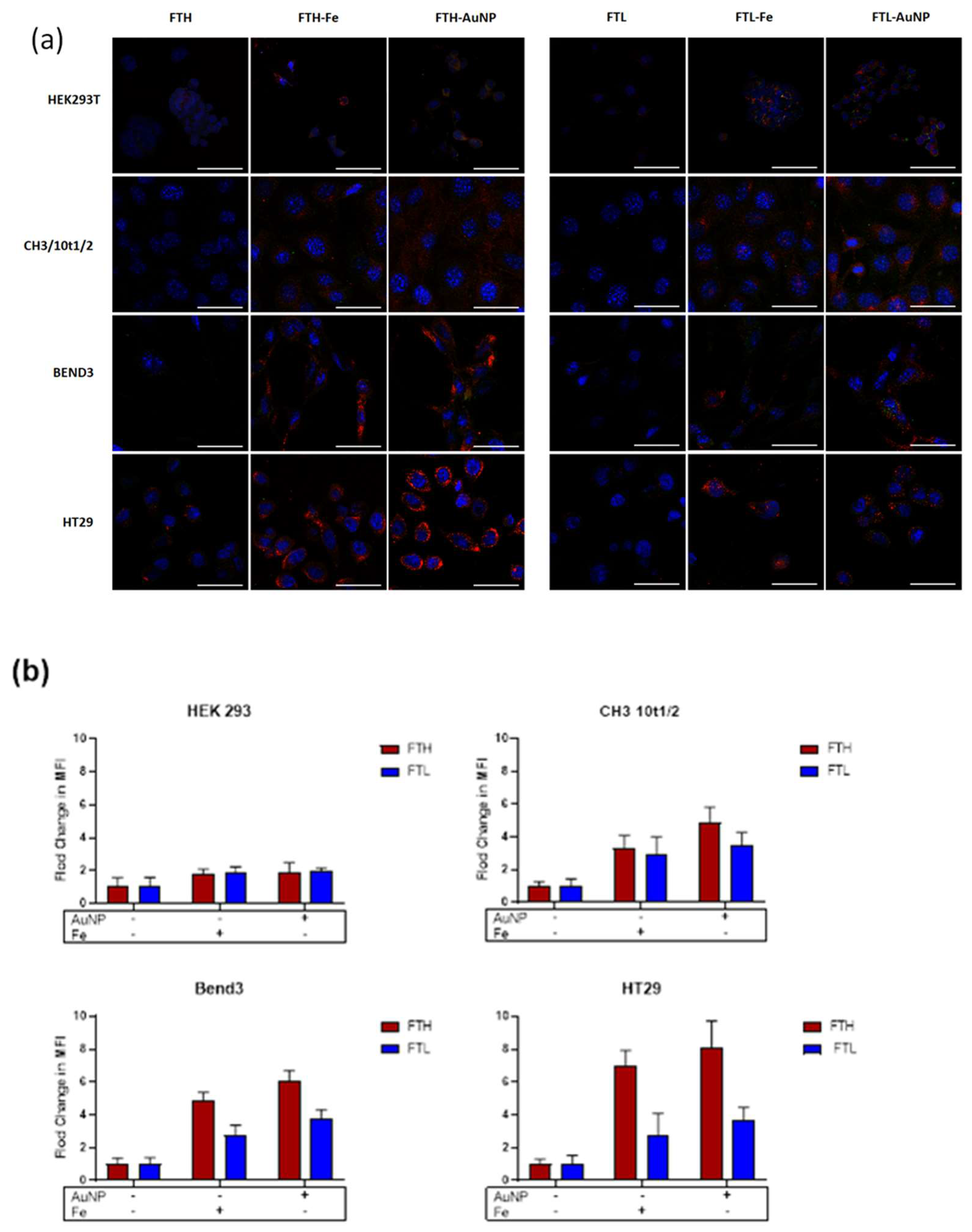
3.3. Cell Uptake of FTH and FTL with Gold Nanoparticles
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barbero, F.; Russo, L.; Vitali, M.; Piella, J.; Salvo, I.; Borrajo, M.L.; Busquets-Fité, M.; Grandori, R.; Bastús, N.G.; Casals, E.; et al. Formation of the Protein Corona: The Interface between Nanoparticles and the Immune System. Semin. Immunol. 2017, 34, 52–60. [Google Scholar] [CrossRef]
- Nie, S. Understanding and Overcoming Major Barriers in Cancer Nanomedicine. Nanomedicine 2010, 5, 523–528. [Google Scholar] [CrossRef] [Green Version]
- Wuelfing, P.; Gross, S.M.; Miles, D.T.; Murray, R.W. Nanometer Gold Clusters Protected by Surface-Bound Monolayers of Thiolated Poly(ethylene Glycol) Polymer Electrolyte. J. Am. Chem. Soc. 1998, 120, 12696–12697. [Google Scholar] [CrossRef]
- Luthuli, S.D.; Chili, M.M.; Revaprasadu, N.; Shonhai, A. Cysteine-Capped Gold Nanoparticles Suppress Aggregation of Proteins Exposed to Heat Stress. IUBMB Life 2013, 65, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Saw, W.S.; Ujihara, M.; Chong, W.Y.; Voon, S.H.; Imae, T.; Kiew, L.V.; Lee, H.B.; Sim, K.S.; Chung, L.Y. Size-Dependent Effect of Cystine/citric Acid-Capped Confeito-like Gold Nanoparticles on Cellular Uptake and Photothermal Cancer Therapy. Colloids Surf. B Biointerfaces 2018, 161, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Harper, B.; Sinche, F.; Wu, R.H.; Gowrishankar, M.; Marquart, G.; Mackiewicz, M.; Harper, S.L. The Impact of Surface Ligands and Synthesis Method on the Toxicity of Glutathione-Coated Gold Nanoparticles. Nanomaterials 2014, 4, 355–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, A.A.; Morgan, J.T.; Brown, P.H.; Adams, A.; Jayasekara, M.P.S.; Zhang, G.; Ackerson, C.J.; Kruhlak, M.J.; Leapman, R.D. Synthesis, Characterization, and Direct Intracellular Imaging of Ultrasmall and Uniform Glutathione-Coated Gold Nanoparticles. Small 2012, 8, 2277–2286. [Google Scholar] [CrossRef] [Green Version]
- Slocik, J.M.; Stone, M.O.; Naik, R.R. Synthesis of Gold Nanoparticles Using Multifunctional Peptides. Small 2005, 1, 1048–1052. [Google Scholar] [CrossRef]
- Murawala, P.; Phadnis, S.M.; Bhonde, R.R.; Prasad, B.L.V. In Situ Synthesis of Water Dispersible Bovine Serum Albumin Capped Gold and Silver Nanoparticles and Their Cytocompatibility Studies. Colloids Surf. B Biointerfaces 2009, 73, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, L.; Kora, A.J.; Arunachalam, J. Highly Stable, Protein Capped Gold Nanoparticles as Effective Drug Delivery Vehicles for Amino-Glycosidic Antibiotics. Mater. Sci. Eng. C Mater. Biol. Appl. 2012, 32, 1571–1577. [Google Scholar] [CrossRef] [PubMed]
- Lillo, C.R.; Calienni, M.N.; Rivas Aiello, B.; Prieto, M.J.; Rodriguez Sartori, D.; Tuninetti, J.; Toledo, P.; Alonso, S.D.V.; Moya, S.; Gonzalez, M.C.; et al. BSA-Capped Gold Nanoclusters as Potential Theragnostic for Skin Diseases: Photoactivation, Skin Penetration, in Vitro, and in Vivo Toxicity. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 112, 110891. [Google Scholar] [CrossRef] [PubMed]
- Khullar, P.; Singh, V.; Mahal, A.; Dave, P.N.; Thakur, S.; Kaur, G.; Singh, J.; Singh Kamboj, S.; Singh Bakshi, M. Bovine Serum Albumin Bioconjugated Gold Nanoparticles: Synthesis, Hemolysis, and Cytotoxicity toward Cancer Cell Lines. J. Phys. Chem. C 2012, 116, 8834–8843. [Google Scholar] [CrossRef]
- Nghiem, T.H.L.; Nguyen, T.T.; Fort, E.; Nguyen, T.P.; Hoang, T.M.N.; Nguyen, T.Q.; Tran, H.N. Capping and in Vivo Toxicity Studies of Gold Nanoparticles. Adv. Nat. Sci: Nanosci. Nanotechnol. 2012, 3, 015002. [Google Scholar]
- Shin, Y.; Dohnalkova, A.; Lin, Y. Preparation of Homogeneous Gold−Silver Alloy Nanoparticles Using the Apoferritin Cavity as a Nanoreactor. J. Phys. Chem. C Nanomater. Interfaces 2010, 114, 5985–5989. [Google Scholar] [CrossRef]
- Fan, R.; Chew, S.W.; Cheong, V.V.; Orner, B.P. Fabrication of Gold Nanoparticles inside Unmodified Horse Spleen Apoferritin. Small 2010, 6, 1483–1487. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Yang, H.; Yuan, Y.; Tian, X.; Wang, L.; Guo, Y.; Xu, L.; Lei, J.; Gao, N.; Anderson, G.J.; et al. Controlling Assembly of Paired Gold Clusters within Apoferritin Nanoreactor for in Vivo Kidney Targeting and Biomedical Imaging. J. Am. Chem. Soc. 2011, 133, 8617–8624. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Yuan, Y.; Xu, Z.; Ji, T.; Tian, Y.; Wu, S.; Lei, J.; Li, J.; Gao, N.; Nie, G. Fine-Tuned H-Ferritin Nanocage with Multiple Gold Clusters as near-Infrared Kidney Specific Targeting Nanoprobe. Bioconjug. Chem. 2015, 26, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Monti, D.M.; Ferraro, G.; Petruk, G.; Maiore, L.; Pane, F.; Amoresano, A.; Cinellu, M.A.; Merlino, A. Ferritin Nanocages Loaded with Gold Ions Induce Oxidative Stress and Apoptosis in MCF-7 Human Breast Cancer Cells. Dalton Trans. 2017, 46, 15354–15362. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Gao, H.; Zhang, Y.; Liu, G.; Niu, G.; Chen, X. Functional Ferritin Nanoparticles for Biomedical Applications. Front. Chem. Sci. Eng. 2017, 11, 633–646. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhang, X.; Zhao, G. Ferritin Nanocage: A Versatile Nanocarrier Utilized in the Field of Food, Nutrition, and Medicine. Nanomaterials 2020, 10, 1894. [Google Scholar] [CrossRef]
- Domínguez-Vera, J.M.; Colacio, E. Nanoparticles of Prussian Blue Ferritin: A New Route for Obtaining Nanomaterials. Inorg. Chem. 2003, 42, 6983–6985. [Google Scholar] [CrossRef]
- Moglia, I.; Santiago, M.; Soler, M.; Olivera-Nappa, A. Silver Nanoparticle Synthesis in Human Ferritin by Photochemical Reduction. J. Inorg. Biochem. 2020, 206, 111016. [Google Scholar] [CrossRef]
- Kasyutich, O.; Ilari, A.; Fiorillo, A.; Tatchev, D.; Hoell, A.; Ceci, P. Silver Ion Incorporation and Nanoparticle Formation inside the Cavity of Pyrococcus Furiosus Ferritin: Structural and Size-Distribution Analyses. J. Am. Chem. Soc. 2010, 132, 3621–3627. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Huang, P.; Jacobson, O.; Wang, Z.; Liu, Y.; Lin, L.; Lin, J.; Lu, N.; Zhang, H.; Tian, R.; et al. Biomineralization-Inspired Synthesis of Copper Sulfide-Ferritin Nanocages as Cancer Theranostics. ACS Nano 2016, 10, 3453–3460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.; Yin, J.-J.; Ning, B.; Wu, X.; Hu, Y.; Ferrari, M.; Anderson, G.J.; Wei, J.; Zhao, Y.; Nie, G. Direct Evidence for Catalase and Peroxidase Activities of Ferritin–platinum Nanoparticles. Biomaterials 2011, 32, 1611–1618. [Google Scholar] [CrossRef] [PubMed]
- Pulsipher, K.W.; Honig, S.; Deng, S.; Dmochowski, I.J. Controlling Gold Nanoparticle Seeded Growth in Thermophilic Ferritin Protein Templates. J. Inorg. Biochem. 2017, 174, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Mosca, M.; Hong, J.; Hadeler, E.; Hakimi, M.; Brownstone, N.; Liao, W.; Bhutani, T. Psoriasis and Cardiometabolic Comorbidities: An Evaluation of the Impact of Systemic Treatments in Randomized Clinical Trials. Dermatol. Ther. 2021, 11, 1497–1520. [Google Scholar] [CrossRef]
- Maity, B.; Abe, S.; Ueno, T. Observation of Gold Sub-Nanocluster Nucleation within a Crystalline Protein Cage. Nat. Commun. 2017, 8, 14820. [Google Scholar] [CrossRef] [Green Version]
- Mosca, L.; Falvo, E.; Ceci, P.; Poser, E.; Genovese, I.; Guarguaglini, G.; Colotti, G. Use of Ferritin-Based Metal-Encapsulated Nanocarriers as Anticancer Agents. NATO Adv. Sci. Inst. Ser. E Appl. Sci. 2017, 7, 101. [Google Scholar] [CrossRef] [Green Version]
- Butts, C.A.; Swift, J.; Kang, S.-G.; Di Costanzo, L.; Christianson, D.W.; Saven, J.G.; Dmochowski, I.J. Directing Noble Metal Ion Chemistry within a Designed Ferritin Protein. Biochemistry 2008, 47, 12729–12739. [Google Scholar] [CrossRef]
- Li, L.; Fang, C.J.; Ryan, J.C.; Niemi, E.C.; Lebrón, J.A.; Björkman, P.J.; Arase, H.; Torti, F.M.; Torti, S.V.; Nakamura, M.C.; et al. Binding and Uptake of H-Ferritin Are Mediated by Human Transferrin Receptor-1. Proc. Natl. Acad. Sci. USA 2010, 107, 3505–3510. [Google Scholar] [CrossRef] [Green Version]
- Daniels, T.R.; Delgado, T.; Rodriguez, J.A.; Helguera, G.; Penichet, M.L. The Transferrin Receptor Part I: Biology and Targeting with Cytotoxic Antibodies for the Treatment of Cancer. Clin. Immunol. 2006, 121, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Tortorella, S.; Karagiannis, T.C. Transferrin Receptor-Mediated Endocytosis: A Useful Target for Cancer Therapy. J. Membr. Biol. 2014, 247, 291–307. [Google Scholar] [CrossRef] [PubMed]
- Todorich, B.; Zhang, X.; Slagle-Webb, B. Tim-2 Is the Receptor for H-ferritin on Oligodendrocytes. J. Neurochem. 2008, 107, 1495–1505. [Google Scholar] [CrossRef]
- Chen, T.T.; Li, L.; Chung, D.-H.; Allen, C.D.C.; Torti, S.V.; Torti, F.M.; Cyster, J.G.; Chen, C.-Y.; Brodsky, F.M.; Niemi, E.C.; et al. TIM-2 Is Expressed on B Cells and in Liver and Kidney and Is a Receptor for H-Ferritin Endocytosis. J. Exp. Med. 2005, 202, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Seaman, W.E.; Di, X.; Wang, W.; Willingham, M.; Torti, F.M.; Torti, S.V. Iron Uptake Mediated by Binding of H-Ferritin to the TIM-2 Receptor in Mouse Cells. PLoS ONE 2011, 6, e23800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.Y.; Paragas, N.; Ned, R.M.; Qiu, A.; Viltard, M.; Leete, T.; Drexler, I.R.; Chen, X.; Sanna-Cherchi, S.; Mohammed, F.; et al. Scara5 Is a Ferritin Receptor Mediating Non-Transferrin Iron Delivery. Dev. Cell 2009, 16, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Geninatti Crich, S.; Cadenazzi, M.; Lanzardo, S.; Conti, L.; Ruiu, R.; Alberti, D.; Cavallo, F.; Cutrin, J.C.; Aime, S. Targeting Ferritin Receptors for the Selective Delivery of Imaging and Therapeutic Agents to Breast Cancer Cells. Nanoscale 2015, 7, 6527–6533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes-Jorge, L.; Ramos, D.; Valença, A.; López-Luppo, M.; Pires, V.M.R.; Catita, J.; Nacher, V.; Navarro, M.; Carretero, A.; Rodriguez-Baeza, A.; et al. L-Ferritin Binding to scara5: A New Iron Traffic Pathway Potentially Implicated in Retinopathy. PLoS ONE 2014, 9, e106974. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Cheng, C.; Wu, Y.; Guo, L.; Kong, D.; Zhang, Z.; Wang, Y.; Zheng, E.; Liu, Y.; He, Y. Interactions of Ferritin with Scavenger Receptor Class A Members. J. Biol. Chem. 2020, 295, 15727–15741. [Google Scholar] [CrossRef] [PubMed]
- Aslan, T.N.; Aşık, E.; Güray, N.T.; Volkan, M. The Potential Application of Gold-Apoferritin Nanocages Conjugated with 2-Amino-2-Deoxy-Glucose for Imaging of Breast Cancer Cells. J. Biol. Inorg. Chem. 2020, 25, 1139–1152. [Google Scholar] [CrossRef] [PubMed]
- Studier, F.W. Protein Production by Auto-Induction in High Density Shaking Cultures. Protein Expr. Purif. 2005, 41, 207–234. [Google Scholar] [CrossRef]
- Huang, X.; El-Sayed, M.A. Gold Nanoparticles: Optical Properties and Implementations in Cancer Diagnosis and Photothermal Therapy. J. Adv. Res. 2010, 1, 13–28. [Google Scholar] [CrossRef] [Green Version]
- Bastús, N.G.; Comenge, J.; Puntes, V. Kinetically Controlled Seeded Growth Synthesis of Citrate-Stabilized Gold Nanoparticles of up to 200 Nm: Size Focusing versus Ostwald Ripening. Langmuir 2011, 27, 11098–11105. [Google Scholar] [CrossRef] [PubMed]
- Keyes, J.D.; Hilton, R.J.; Farrer, J.; Watt, R.K. Ferritin as a Photocatalyst and Scaffold for Gold Nanoparticle Synthesis. J. Nanopart. Res. 2011, 13, 2563–2575. [Google Scholar] [CrossRef]
- Moglia, I.; Santiago, M.; Olivera-Nappa, Á.; Soler, M. An Optimized Low-Cost Protocol for Standardized Production of Iron-Free Apoferritin Nanocages with High Protein Recovery and Suitable Conformation for Nanotechnological Applications. J. Inorg. Biochem. 2018, 183, 184–190. [Google Scholar] [CrossRef]
- Petrucci, O.D.; Hilton, R.J.; Farrer, J.K.; Watt, R.K. A Ferritin Photochemical Synthesis of Monodispersed Silver Nanoparticles That Possess Antimicrobial Properties. J. Nanomater. 2019, 1–8. [Google Scholar] [CrossRef]
- Yang, Y.; Han, A.; Li, R.; Fang, G.; Liu, J.; Wang, S. Synthesis of Highly Fluorescent Gold Nanoclusters and Their Use in Sensitive Analysis of Metal Ions. Analyst 2017, 142, 4486–4493. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Zheng, Y.; Ying, J.Y. Protein-Directed Synthesis of Highly Fluorescent Gold Nanoclusters. J. Am. Chem. Soc. 2009, 131, 888–889. [Google Scholar] [CrossRef]
- Jiang, X.; Sun, C.; Guo, Y.; Nie, G.; Xu, L. Peroxidase-like Activity of Apoferritin Paired Gold Clusters for Glucose Detection. Biosens. Bioelectron. 2015, 64, 165–170. [Google Scholar] [CrossRef]
- Maity, B.; Hishikawa, Y.; Lu, D.; Ueno, T. Recent Progresses in the Accumulation of Metal Ions into the Apo-Ferritin Cage: Experimental and Theoretical Perspectives. Polyhedron 2019, 172, 104–111. [Google Scholar] [CrossRef]
- Melníková, L.; Mitróová, Z.; Timko, M.; Kováč, J.; Avdeev, M.V.; Petrenko, V.I.; Garamus, V.M.; Almásy, L.; Kopčanský, P. Structural Characterization of Magnetoferritin. Mendeleev Commun. 2014, 24, 80–81. [Google Scholar] [CrossRef] [Green Version]
- Balejcikova, L.; Molcan, M.; Kovac, J.; Kubovcikova, M.; Saksl, K.; Mitroova, Z.; Timko, M.; Kopcansky, P. Hyperthermic Effect in Magnetoferritin Aqueous Colloidal Solution. J. Mol. Liq. 2019, 283, 39–44. [Google Scholar] [CrossRef]
- Welch, K.D.; Van Eden, M.E.; Aust, S.D. Modification of Ferritin during Iron Loading. Free Radic. Biol. Med. 2001, 31, 999–1006. [Google Scholar] [CrossRef]
- Welch, K.D.; Reilly, C.A.; Aust, S.D. The Role of Cysteine Residues in the Oxidation of Ferritin. Free Radic. Biol. Med. 2002, 33, 399–408. [Google Scholar] [CrossRef]
- Yang, C.; Cao, C.; Cai, Y.; Xu, H.; Zhang, T.; Pan, Y. Effects of PEGylation on Biomimetic Synthesis of Magnetoferritin Nanoparticles. J. Nanopart. Res. 2017, 19, 101. [Google Scholar] [CrossRef]
- Cai, Y.; Cao, C.; He, X.; Yang, C.; Tian, L.; Zhu, R.; Pan, Y. Enhanced Magnetic Resonance Imaging and Staining of Cancer Cells Using Ferrimagnetic H-Ferritin Nanoparticles with Increasing Core Size. Int. J. Nanomedicine 2015, 10, 2619–2634. [Google Scholar]
- Cao, C.; Wang, X.; Cai, Y.; Sun, L.; Tian, L.; Wu, H.; He, X.; Lei, H.; Liu, W.; Chen, G.; et al. Targeted In Vivo Imaging of Microscopic Tumors with Ferritin-Based Nanoprobes across Biological Barriers. Adv. Mater. 2014, 26, 2566–2571. [Google Scholar] [CrossRef]
- Koralewski, M.; Balejčíková, L.; Mitróová, Z.; Pochylski, M.; Baranowski, M.; Kopčanský, P. Morphology and Magnetic Structure of the Ferritin Core during Iron Loading and Release by Magnetooptical and NMR Methods. ACS Appl. Mater. Interfaces 2018, 10, 7777–7787. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wei, W.; Wang, C.; Yue, H.; Ma, D.; Zhu, C.; Ma, G.; Du, Y. Apoferritin-Camouflaged Pt Nanoparticles: Surface Effects on Cellular Uptake and Cytotoxicity. J. Mater. Chem. 2011, 21, 7105–7110. [Google Scholar] [CrossRef]
- Kashanian, S.; Abasi Tarighat, F.; Rafipour, R.; Abbasi-Tarighat, M. Biomimetic Synthesis and Characterization of Cobalt Nanoparticles Using Apoferritin, and Investigation of Direct Electron Transfer of Co(NPs)-Ferritin at Modified Glassy Carbon Electrode to Design a Novel Nanobiosensor. Mol. Biol. Rep. 2012, 39, 8793–8802. [Google Scholar] [CrossRef]
- Lv, C.; Yin, S.; Zhang, X.; Hu, J.; Zhang, T.; Zhao, G. 16-Mer Ferritin-like Protein Templated Gold Nanoclusters for Bioimaging Detection of Methylmercury in the Brain of Living Mice. Anal. Chim. Acta 2020, 1127, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Alkilany, A.M.; Murphy, C.J. Toxicity and Cellular Uptake of Gold Nanoparticles: What We Have Learned so Far? J. Nanopart. Res. 2010, 12, 2313–2333. [Google Scholar] [CrossRef] [Green Version]
- De Vico, G.; Martano, M.; Maiolino, P.; Carella, F.; Leonardi, L. Expression of Transferrin Receptor-1 (TFR-1) in Canine Osteosarcomas. Vet. Med. Sci 2020, 6, 272–276. [Google Scholar] [CrossRef]
- Conti, L.; Lanzardo, S.; Ruiu, R.; Cadenazzi, M.; Cavallo, F.; Aime, S.; Geninatti Crich, S. L-Ferritin Targets Breast Cancer Stem Cells and Delivers Therapeutic and Imaging Agents. Oncotarget 2016, 7, 66713–66727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Tian, S.; Petros, R.A.; Napier, M.E.; Desimone, J.M. The Complex Role of Multivalency in Nanoparticles Targeting the Transferrin Receptor for Cancer Therapies. J. Am. Chem. Soc. 2010, 132, 11306–11313. [Google Scholar] [CrossRef] [Green Version]
- Fan, K.; Jia, X.; Zhou, M.; Wang, K.; Conde, J.; He, J.; Tian, J.; Yan, X. Ferritin Nanocarrier Traverses the Blood Brain Barrier and Kills Glioma. ACS Nano 2018, 12, 4105–4115. [Google Scholar] [CrossRef]
- Cui, C.; Cheng, X.; Yan, L.; Ding, H.; Guan, X.; Zhang, W.; Tian, X.; Hao, C. Downregulation of TfR1 Promotes Progression of Colorectal Cancer via the JAK/STAT Pathway. Cancer Manag. Res. 2019, 11, 6323–6341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhupanapadu Sunkesula, S.R.; Luo, X.; Das, D.; Singh, A.; Singh, N. Iron Content of Ferritin Modulates Its Uptake by Intestinal Epithelium: Implications for Co-Transport of Prions. Mol. Brain 2010, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Brylinski, M.; Skolnick, J. What Is the Relationship between the Global Structures of Apo and Holo Proteins? Proteins 2008, 70, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.J.; Benson, M.L.; Smith, R.D.; Carlson, H.A. Inherent versus Induced Protein Flexibility: Comparisons within and between Apo and Holo Structures. PLoS Comput. Biol. 2019, 15, e1006705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolf, T.B.; Grossfield, A.; Tychko, M. Differences between Apo and Three Holo Forms of the Intestinal Fatty Acid Binding Protein Seen by Molecular Dynamics Computer Calculations. Biophys. J. 2000, 78, 608–625. [Google Scholar] [CrossRef] [Green Version]
- Hunter, N.H.; Bakula, B.C.; Bruce, C.D. Molecular Dynamics Simulations of Apo and Holo Forms of Fatty Acid Binding Protein 5 and Cellular Retinoic Acid Binding Protein II Reveal Highly Mobile Protein, Retinoic Acid Ligand, and Water Molecules. J. Biomol. Struct. Dyn. 2018, 36, 1893–1907. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence 5′–3′ |
|---|---|
| FTH-F | TCGCCCATATGACGACCGCGTCCACCTC |
| FTH-R | TTGGTCTCGAGGGAAGTCACCCCACGGCTATG |
| FTL-F | AGCCACATATGAGCTCCCAGATTCGTCAG |
| FTL-R | GGGCCCTCGAGAAGTCGCTGGGCTCAGAAG |
| Apo FTL | FTL-AuNP | Apo FTH | FTH-AuNP | |
|---|---|---|---|---|
| Monomer | 60.1% | 74.4% | 73.3% | 64.3% |
| Oligomer | 39.9% | 25.6% | 26.7% | 35.7% |
| Ratio | 1.5:1 | 2.9:1 | 2.7:1 | 1.8:1 |
| Sample | Hd (nm) | Z-Pot (mV) |
|---|---|---|
| Apo FTH | 18 ± 12 | −13.5 |
| Apo FTL | 20 ± 9 | −13.7 |
| FTH-AuNP | 55 ± 29 | −10.2 |
| FTL-AuNP | 35 ± 16 | −7.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moglia, I.; Santiago, M.; Guerrero, S.; Soler, M.; Olivera-Nappa, A.; Kogan, M.J. Enhanced Cellular Uptake of H-Chain Human Ferritin Containing Gold Nanoparticles. Pharmaceutics 2021, 13, 1966. https://doi.org/10.3390/pharmaceutics13111966
Moglia I, Santiago M, Guerrero S, Soler M, Olivera-Nappa A, Kogan MJ. Enhanced Cellular Uptake of H-Chain Human Ferritin Containing Gold Nanoparticles. Pharmaceutics. 2021; 13(11):1966. https://doi.org/10.3390/pharmaceutics13111966
Chicago/Turabian StyleMoglia, Italo, Margarita Santiago, Simon Guerrero, Mónica Soler, Alvaro Olivera-Nappa, and Marcelo J. Kogan. 2021. "Enhanced Cellular Uptake of H-Chain Human Ferritin Containing Gold Nanoparticles" Pharmaceutics 13, no. 11: 1966. https://doi.org/10.3390/pharmaceutics13111966
APA StyleMoglia, I., Santiago, M., Guerrero, S., Soler, M., Olivera-Nappa, A., & Kogan, M. J. (2021). Enhanced Cellular Uptake of H-Chain Human Ferritin Containing Gold Nanoparticles. Pharmaceutics, 13(11), 1966. https://doi.org/10.3390/pharmaceutics13111966