
Folate-Equipped Cationic Liposomes Deliver Anti-MDR1-siRNA to the Tumor and Increase the Efficiency of Chemotherapy

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Liposome Preparation
2.2. siRNAs
2.3. Preparation of the Complexes of Cationic Liposomes and siRNA
2.4. Liposome Sizes and Zeta Potentials
2.5. Cell Lines and Culture Conditions
2.6. Mice and Tumor Models
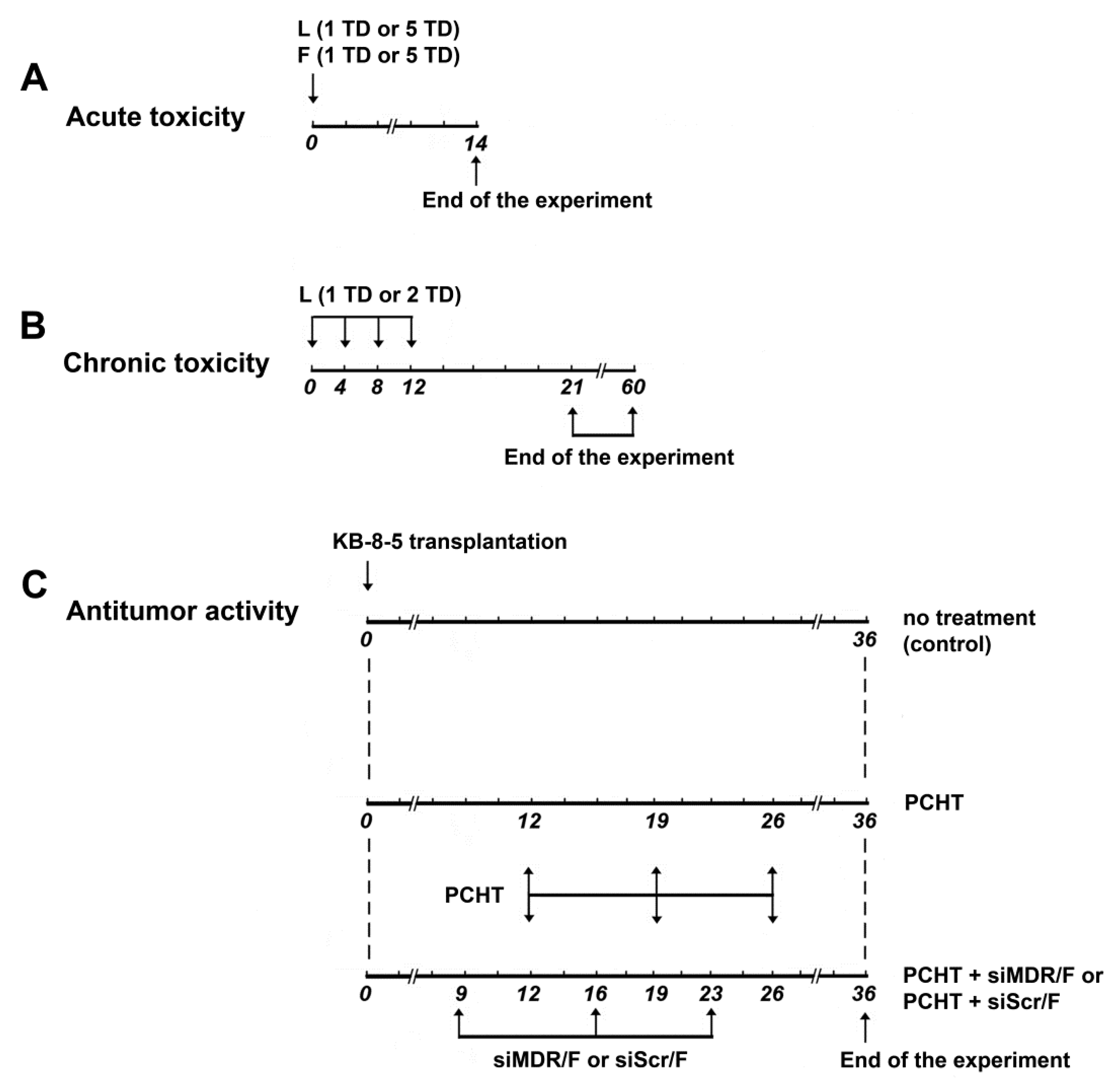
2.7. Acute and Chronic Toxicity
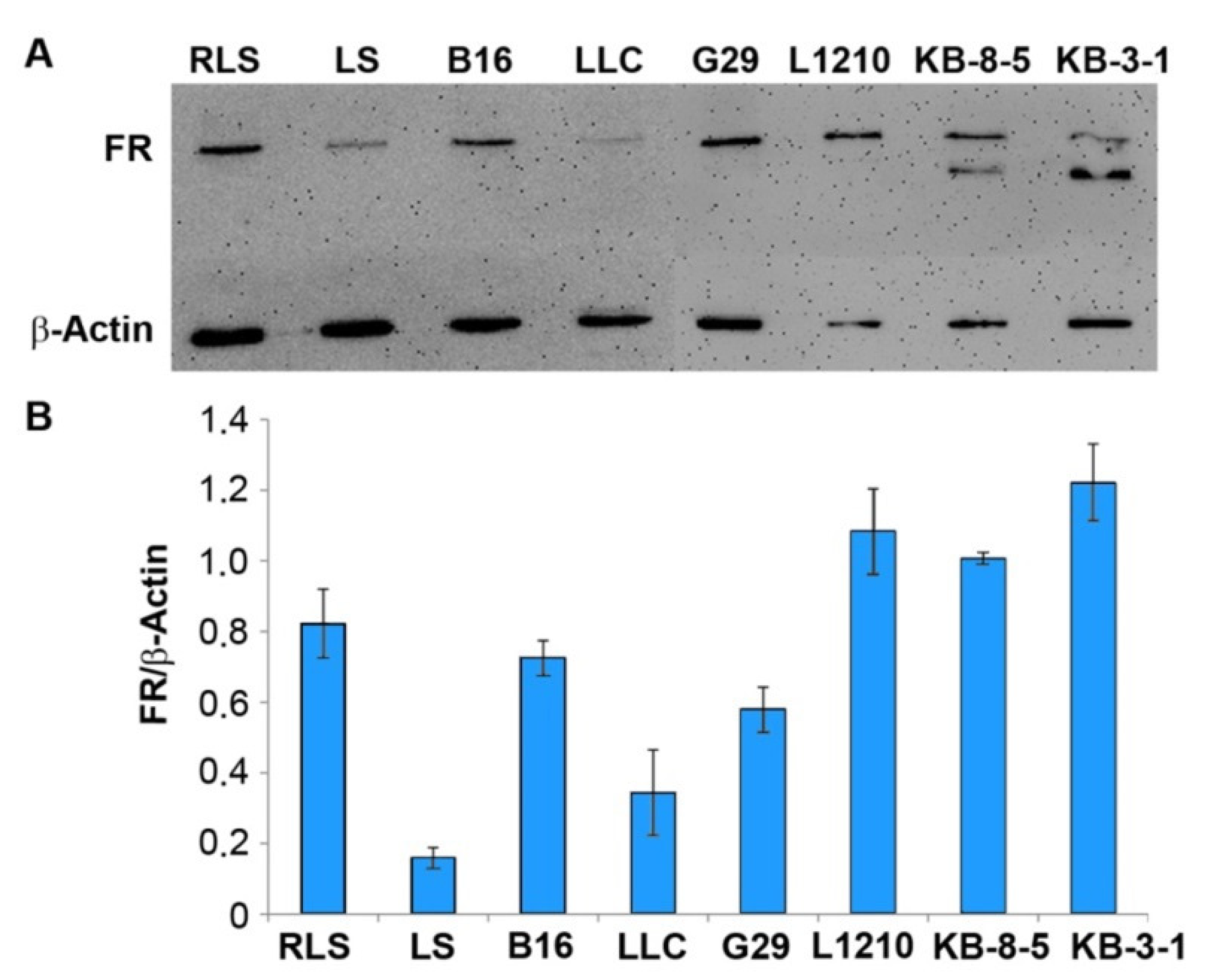
2.8. Evaluation of FR Expression by Western Blotting
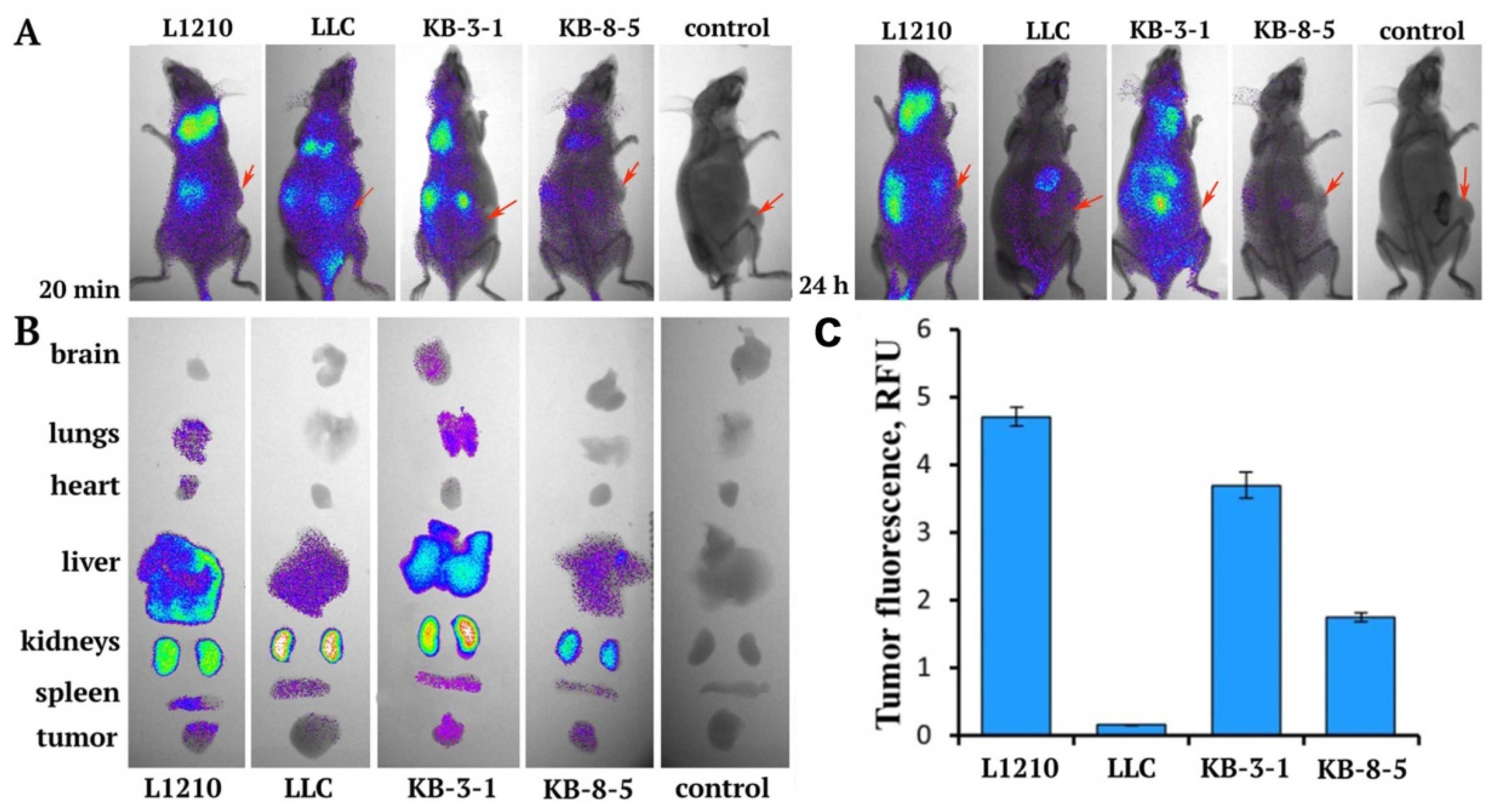
2.9. In Vivo Biodistribution Studies
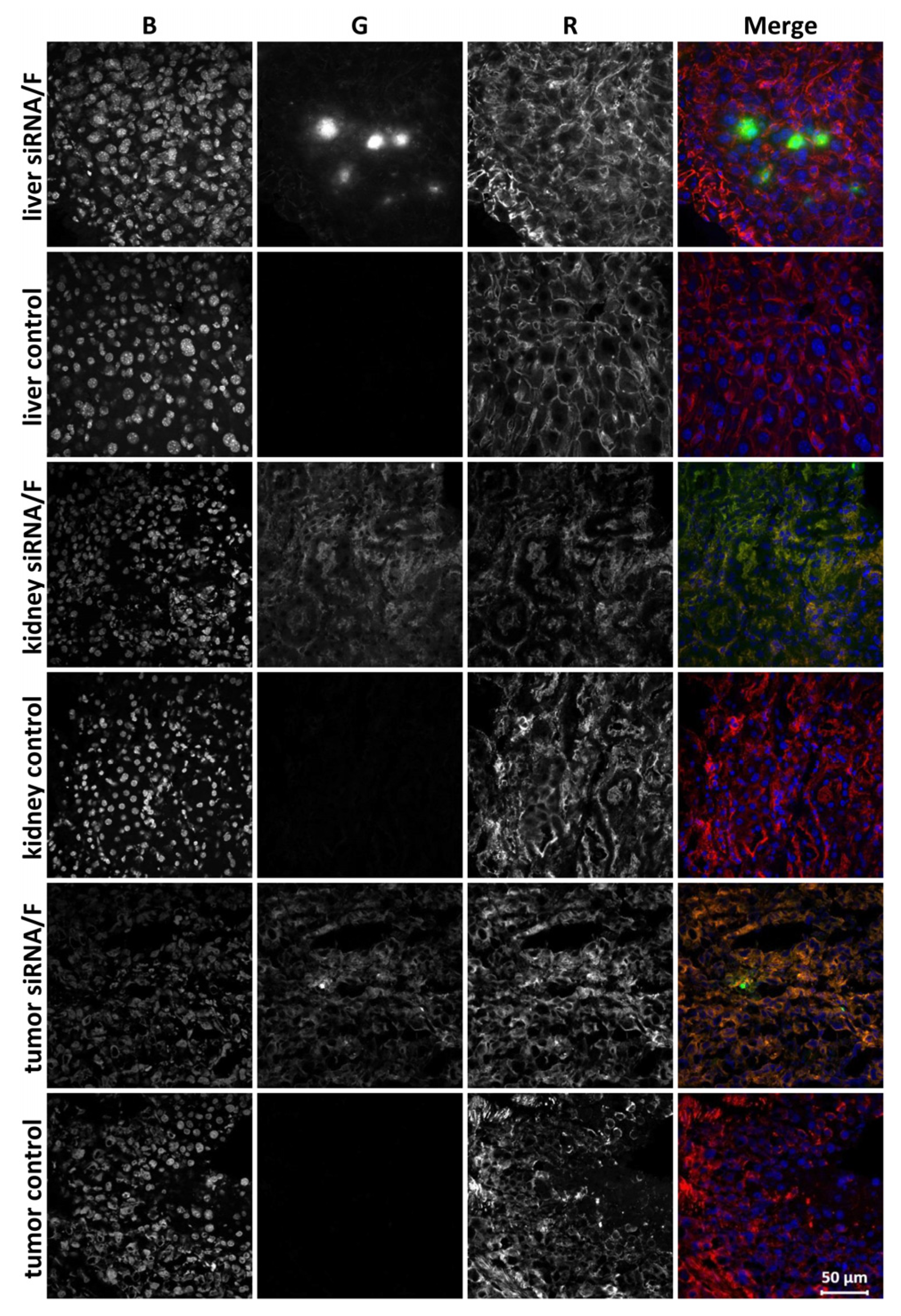
2.10. Confocal Microscopy
2.11. In Vivo Antitumor Assay
2.12. Histology
2.13. Statistical Analysis
3. Results
3.1. Formulations L and F Have No Acute and Chronic Toxicity in Mice
3.2. The Abundance of FR in Tumor Cells of Different Origin
3.3. Biodistribution of siMDR in Tumor-Bearing Mice and Confocal Microscopy of Tumor Sections
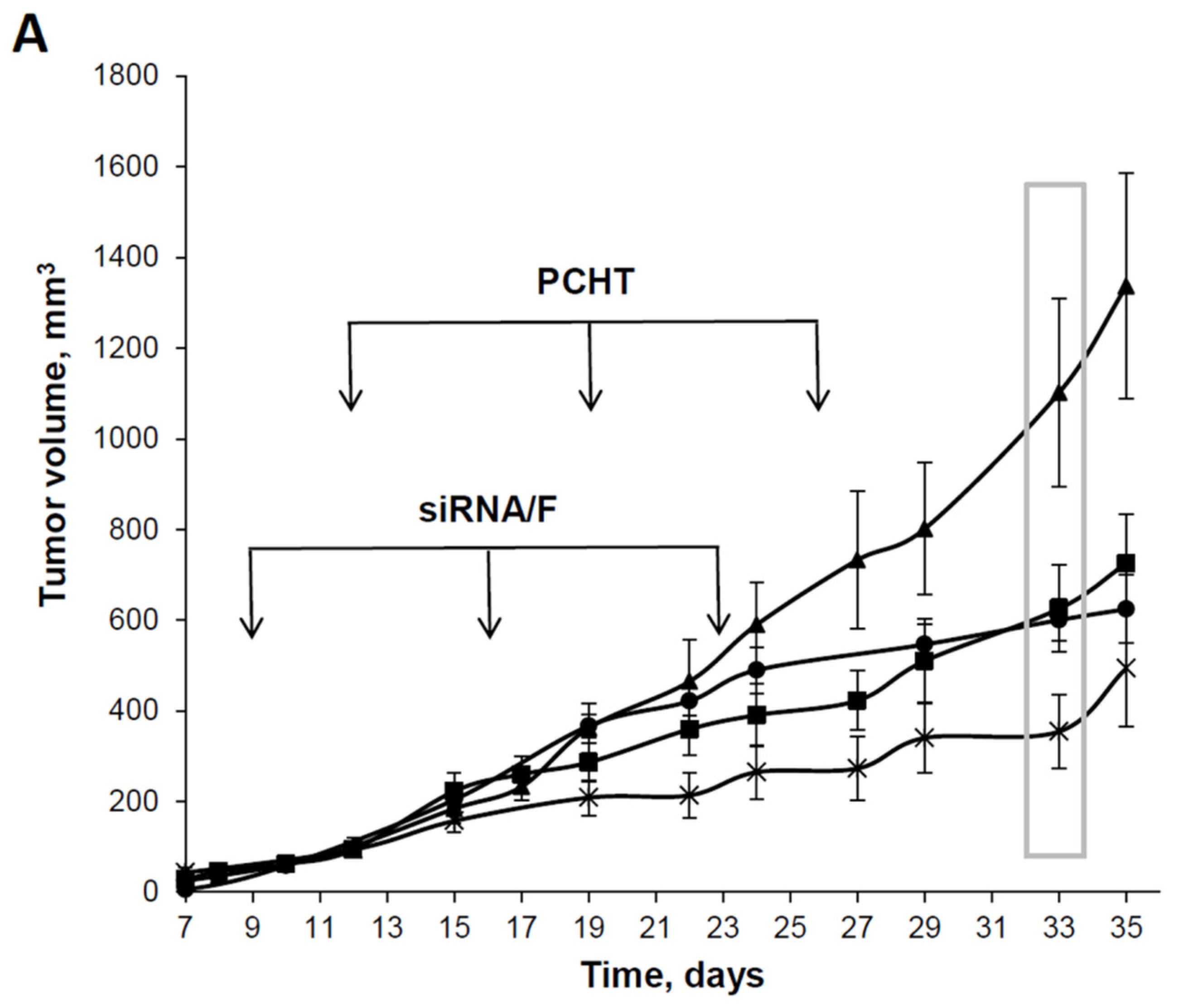
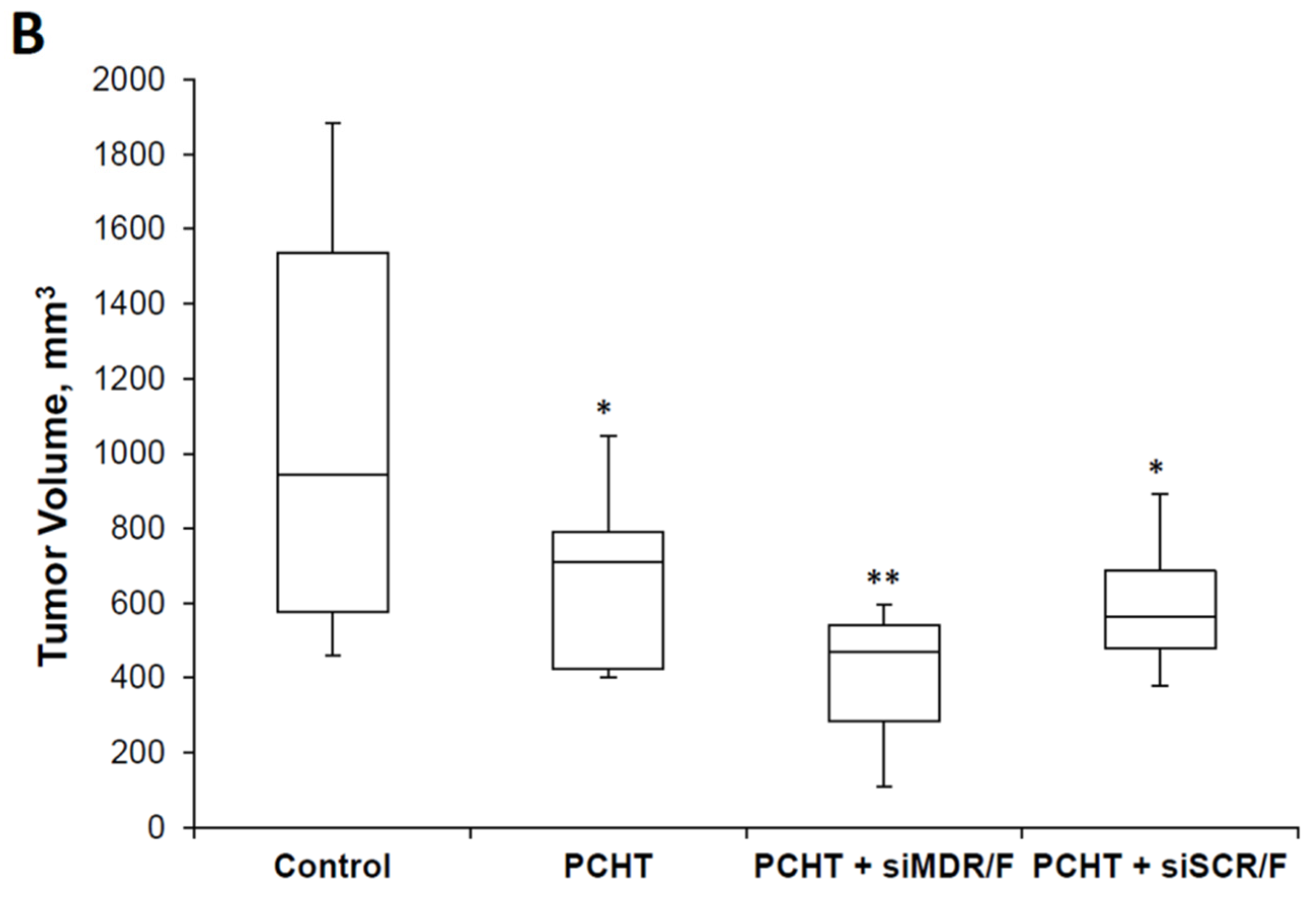
3.4. The Effects of Combination Therapy including Anti-MDR1 siRNA/F Complexes and PCHT on a Multidrug-Resistant KB-8-5 Tumor Xenograft in SCID Mice
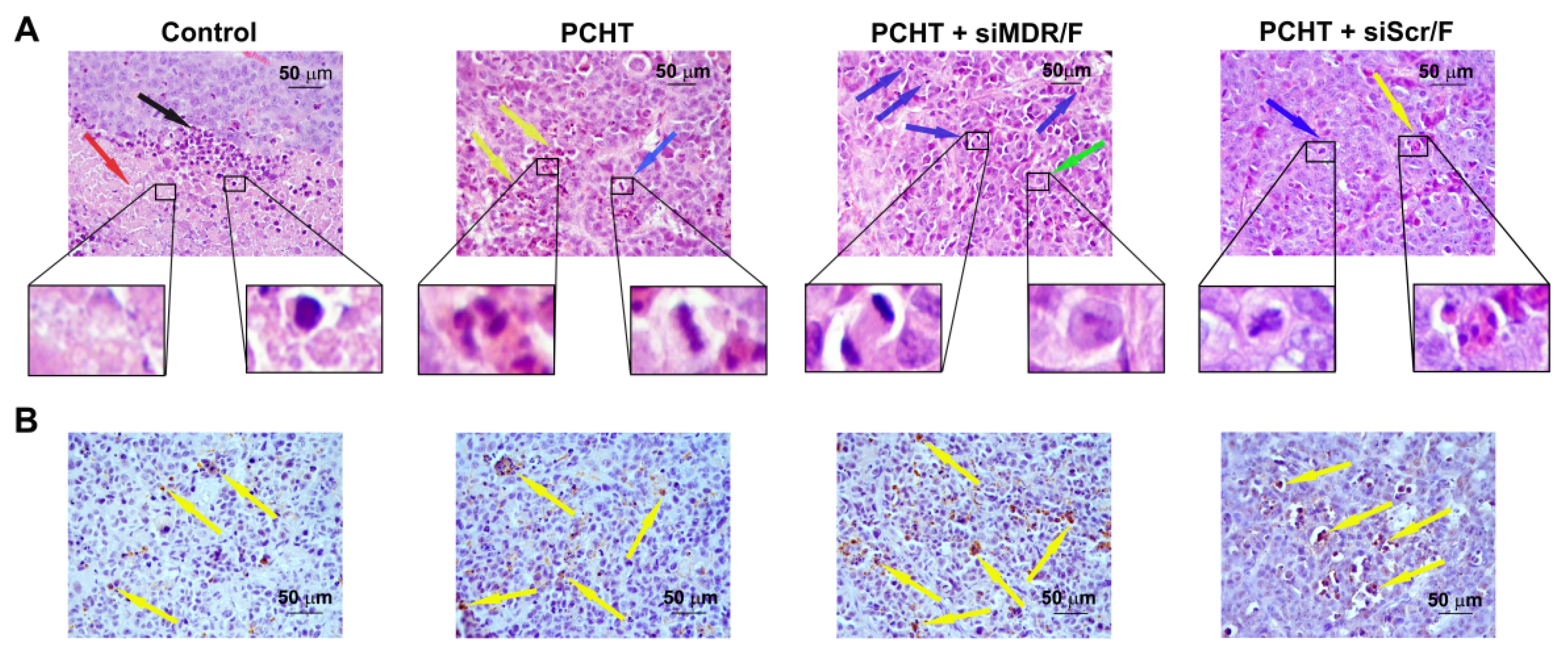
3.5. siMDR/F Combined with PCHT Reduces Necrosis and Inflammation, and Stimulates Apoptosis in KB-8-5 Tumor Tissue
3.6. siMDR/F Combined with PCHT Does Not Induce Liver Toxicity in KB-8-5 Tumor-Bearing Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ginn, S.L.; Alexander, I.E.; Edelstein, M.L.; Abedi, M.R.; Wixon, J. Gene therapy clinical trials worldwide to 2012—An update. J. Gene Med. 2013, 15, 65–77. [Google Scholar] [CrossRef]
- Kabilova, T.O.; Shmendel, E.V.; Gladkikh, D.V.; Chernolovskaya, E.L.; Markov, O.V.; Morozova, N.G.; Maslov, M.A.; Zenkova, M.A. Targeted delivery of nucleic acids into xenograft tumors mediated by novel folate-equipped liposomes. Eur. J. Pharm. Biopharm. 2018, 123, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Collin, R.W.J.; Garanto, A. Applications of antisense oligonucleotides for the treatment of inherited retinal diseases. Curr. Opin. Ophthalmol. 2017, 28, 260–266. [Google Scholar] [CrossRef]
- Wong, S.Y.; Pelet, J.M.; Putnam, D. Polymer systems for gene delivery-Past, present, and future. Prog. Polym. Sci. 2007, 32, 799–837. [Google Scholar] [CrossRef]
- Gu, F.X.; Karnik, R.; Wang, A.Z.; Alexis, F.; Levy-Nissenbaum, E.; Hong, S.; Langer, R.S.; Farokhzad, O.C. Targeted nanoparticles for cancer therapy. Nano Today 2007, 2, 14–21. [Google Scholar] [CrossRef]
- Gavrilov, K.; Saltzman, W.M. Therapeutic siRNA: Principles, challenges, and strategies. Yale J. Biol. Med. 2012, 85, 187–200. Available online: http://www.ncbi.nlm.nih.gov/pubmed/22737048 (accessed on 21 May 2018). [PubMed]
- Lee, H.; Lytton-Jean, A.K.R.; Chen, Y.; Love, K.T.; Park, A.I.; Karagiannis, E.D.; Sehgal, A.; Querbes, W.; Zurenko, C.S.; Jayaraman, M.; et al. Molecularly self-assembled nucleic acid nanoparticles for targeted in vivo siRNA delivery. Nat. Nanotechnol. 2012, 7, 389–393. [Google Scholar] [CrossRef]
- Akinc, A.; Querbes, W.; De, S.; Qin, J.; Frank-Kamenetsky, M.; Jayaprakash, K.N.; Jayaraman, M.; Rajeev, K.G.; Cantley, W.L.; Dorkin, J.R.; et al. Targeted Delivery of RNAi Therapeutics With Endogenous and Exogenous Ligand-Based Mechanisms. Mol. Ther. 2010, 18, 1357–1364. [Google Scholar] [CrossRef]
- Höbel, S.; Loos, A.; Appelhans, D.; Schwarz, S.; Seidel, J.; Voit, B.; Aigner, A. Maltose- and maltotriose-modified, hyperbranched poly(ethylene imine)s (OM-PEIs): Physicochemical and biological properties of DNA and siRNA complexes. J. Control. Release 2011, 149, 146–158. [Google Scholar] [CrossRef]
- Järver, P.; Mäger, I.; Langel, Ü. In vivo biodistribution and efficacy of peptide mediated delivery. Trends Pharmacol. Sci. 2010, 31, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Dufès, C.; al Robaian, M.; Somani, S. Transferrin and the transferrin receptor for the targeted delivery of therapeutic agents to the brain and cancer cells. Ther. Deliv. 2013, 4, 629–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markov, O.V.; Mironova, N.L.; Shmendel, E.V.; Maslov, M.A.; Zenkova, M.A. Systemic delivery of complexes of melanoma RNA with mannosylated liposomes activates highly efficient murine melanoma-specific cytotoxic T cells in vivo. Mol. Biol. 2017, 51, 118–125. [Google Scholar] [CrossRef]
- Li, S.D.; Huang, L. Pharmacokinetics and biodistribution of nanoparticles. Mol. Pharm. 2008, 5, 496–504. [Google Scholar] [CrossRef]
- Fanciullino, R.; Ciccolini, J. Liposome-encapsulated anticancer drugs: Still waiting for the magic bullet? Curr. Med. Chem. 2009, 16, 4361–4371. Available online: http://www.ncbi.nlm.nih.gov/pubmed/19835568 (accessed on 21 May 2018). [CrossRef] [PubMed]
- Parker, N.; Turk, M.J.; Westrick, E.; Lewis, J.D.; Low, P.S.; Leamon, C.P. Folate receptor expression in carcinomas and normal tissues determined by a quantitative radioligand binding assay. Anal. Biochem. 2005, 338, 284–293. [Google Scholar] [CrossRef]
- Low, P.S.; Kularatne, S.A. Folate-targeted therapeutic and imaging agents for cancer. Curr. Opin. Chem. Biol. 2009, 13, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Low, P.S. Folate-targeted therapies for cancer. J. Med. Chem. 2010, 53, 6811–6824. [Google Scholar] [CrossRef]
- Yoshizawa, T.; Hattori, Y.; Hakoshima, M.; Koga, K.; Maitani, Y. Folate-linked lipid-based nanoparticles for synthetic siRNA delivery in KB tumor xenografts. Eur. J. Pharm. Biopharm. 2008, 70, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Kaul, A.; Chaturvedi, S.; Attri, A.; Kalra, M.; Mishra, A.K. Targeted theranostic liposomes: Rifampicin and ofloxacin loaded pegylated liposomes for theranostic application in mycobacterial infections. RSC Adv. 2016, 6, 28919–28926. [Google Scholar] [CrossRef]
- Gabizon, A.; Horowitz, A.T.; Goren, D.; Tzemach, D.; Shmeeda, H.; Zalipsky, S. In Vivo Fate of Folate-Targeted Polyethylene-Glycol Liposomes in Tumor-Bearing Mice. Clin. Cancer Res. 2003, 9, 6551–6559. [Google Scholar]
- Duarte, S.; Faneca, H.; de Lima, M.C.P. Folate-associated lipoplexes mediate efficient gene delivery and potent antitumoral activity in vitro and in vivo. Int. J. Pharm. 2012, 423, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Wang, T.; Tang, R.; Wang, J.; Long, H.; Gao, X.; Tang, S. Silencing of the MYCN gene by siRNA delivered by folate receptor-targeted liposomes in LA-N-5 cells. Pediatr. Surg. Int. 2010, 26, 1185–1191. [Google Scholar] [CrossRef] [PubMed]
- Mokhtarzadeh, A.; Alibakhshi, A.; Hashemi, M.; Hejazi, M.; Hosseini, V.; de la Guardia, M.; Ramezani, M. Biodegradable nano-polymers as delivery vehicles for therapeutic small non-coding ribonucleic acids. J. Control. Release 2017, 245, 116–126. [Google Scholar] [CrossRef]
- Li, L.; Yang, Q.; Zhou, Z.; Zhong, J.; Huang, Y. Doxorubicin-loaded, charge reversible, folate modified HPMA copolymer conjugates for active cancer cell targeting. Biomaterials 2014, 35, 5171–5187. [Google Scholar] [CrossRef]
- Yang, T.; Li, B.; Qi, S.; Liu, Y.; Gai, Y.; Ye, P.; Yang, G.; Zhang, W.; Zhang, P.; He, X.; et al. Co-delivery of doxorubicin and Bmil siRNA by folate receptor targeted liposomes exhibits enhanced anti-tumor effects in vitro and in vivo. Theranostics 2014, 4, 1096–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Jeong, J.H.; Park, T.G. A new gene delivery formulation of polyethylenimine/DNA complexes coated with PEG conjugated fusogenic peptide. J. Control. Release 2001, 76, 183–192. [Google Scholar] [CrossRef]
- Li, J.M.; Wang, Y.Y.; Zhang, W.; Su, H.; Ji, L.N.; Mao, Z.W. Low-weight polyethylenimine cross-linked 2-hydroxypopyl-β-cyclodextrin and folic acid as an efficient and nontoxic siRNA carrier for gene silencing and tumor inhibition by VEGF siRNA. Int. J. Nanomed. 2013, 8, 2101–2117. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; He, L.; Liu, Y.; Xia, S.; Fang, A.; Xie, Y.; Gan, L.; He, Z.; Tan, X.; Jiang, C.; et al. Promising Nanocarriers for PEDF Gene Targeting Delivery to Cervical Cancer Cells Mediated by the Over-expressing FRα. Sci. Rep. 2016, 6, 32427. [Google Scholar] [CrossRef] [Green Version]
- He, Z.Y.; Wei, X.W.; Luo, M.; Luo, S.T.; Yang, Y.; Yu, Y.Y.; Chen, Y.; Ma, C.C.; Liang, X.; Guo, F.C.; et al. Folate-linked lipoplexes for short hairpin RNA targeting claudin-3 delivery in ovarian cancer xenografts. J. Control. Release 2013, 172, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Maslov, M.A.; Kabilova, T.O.; Petukhov, I.A.; Morozova, N.G.; Serebrennikova, G.A.; Vlassov, V.V.; Zenkova, M.A. Novel cholesterol spermine conjugates provide efficient cellular delivery of plasmid DNA and small interfering RNA. J. Control. Release 2012, 160, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Bellon, L. Oligoribonucleotides with 2′-O-( tert -Butyldimethylsilyl) Groups. Curr. Protoc. Nucleic Acid Chem. 2000, 1, 3–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shmendel, E.; Kabilova, T.; Morozova, N.; Zenkova, M.; Maslov, M. Effects of spacers within a series of novel folate-containing lipoconjugates on the targeted delivery of nucleic acids. J. Drug Deliv. Sci. Technol. 2020, 57, 101609. [Google Scholar] [CrossRef]
- Sen’kova, A.V.; Savin, I.A.; Kabilova, T.O.; Zenkova, M.A.; Chernolovskaya, E.L. Tumor-Suppressing, Immunostimulating, and Hepatotoxic Effects of Immunostimulatory RNA in Combination with Dacarbazine in a Murine Melanoma Model. Mol. Biol. 2020, 54, 267–277. [Google Scholar] [CrossRef]
- Shmendel, E.V.; Kabilova, T.O.; Morozova, N.G.; Zenkova, M.A.; Maslov, M.A. Targeted Delivery of Nucleic Acids by Folate-Containing Liposomes into KB-3-1 and HEK 293 Cells. Russ. J. Bioorg. Chem. 2019, 45, 719–725. [Google Scholar] [CrossRef]
- Chernikov, I.V.; Gladkikh, D.V.; Meschaninova, M.I.; Ven’yaminova, A.G.; Zenkova, M.A.; Vlassov, V.V.; Chernolovskaya, E.L. Cholesterol-Containing Nuclease-Resistant siRNA Accumulates in Tumors in a Carrier-free Mode and Silences MDR1 Gene. Mol. Ther. Nucleic Acids 2017, 6, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Sen’kova, A.V.; Mironova, N.L.; Patutina, O.A.; Ageeva, T.A.; Zenkova, M.A. The Toxic Effects of Polychemotherapy onto the Liver Are Accelerated by the Upregulated MDR of Lymphosarcoma. ISRN Oncol. 2012, 2012, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Mironova, N.L.; Petrushanko, I.Y.; Patutina, O.A.; Sen’kova, A.V.; Simonenko, O.V.; Mitkevich, V.A.; Markov, O.V.; Zenkova, M.A.; Makarov, A.A. Ribonuclease binase inhibits primary tumor growth and metastases via apoptosis induction in tumor cells. Cell Cycle 2013, 12, 2120–2131. [Google Scholar] [CrossRef]
- Petrova, N.S.; Meschaninova, M.I.; Venyaminova, A.G.; Zenkova, M.A.; Vlassov, V.V.; Chernolovskaya, E.L. Silencing activity of 2′-O-methyl modified anti-MDR1 siRNAs with mismatches in the central part of the duplexes. FEBS Lett. 2011, 585, 2352–2356. [Google Scholar] [CrossRef] [Green Version]
- Volkov, A.A.; Kruglova, N.S.; Meschaninova, M.I.; Venyaminova, A.G.; Zenkova, M.A.; Vlassov, V.V.; Chernolovskaya, E.L. Selective Protection of Nuclease-Sensitive Sites in siRNA Prolongs Silencing Effect. Oligonucleotides 2009, 19, 191–202. [Google Scholar] [CrossRef]
- Euliss, L.E.; DuPont, J.A.; Gratton, S.; DeSimone, J. Imparting size, shape, and composition control of materials for nanomedicine. Chem. Soc. Rev. 2006, 35, 1095–1104. [Google Scholar] [CrossRef]
- Tulve, N.S.; Stefaniak, A.B.; Vance, M.E.; Rogers, K.; Mwilu, S.; LeBouf, R.F.; Schwegler-Berry, D.; Willis, R.; Thomas, T.A.; Marr, L.C. Characterization of silver nanoparticles in selected consumer products and its relevance for predicting children’s potential exposures. Int. J. Hyg. Environ. Health 2015, 218, 345–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; Xue, J.; Gao, S.; Lu, A.; Yang, D.; Jiang, H.; Shi, K. Cleavable PEGylation: A strategy for overcoming the “ PEG dilemma ” in efficient drug delivery. Drug Deliv. 2017, 24, 22–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabilova, T.; Shmendel, E.; Gladkikh, D.; Morozova, N.; Maslov, M.; Chernolovskaya, E.; Vlassov, V.; Zenkova, M. Novel PEGylated liposomes enhance immunostimulating activity of isRNA. Molecules 2018, 23, 3101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farhood, H.; Serbina, N.; Huang, L. The role of dioleoyl phosphatidylethanolamine in cationic liposome mediated gene transfer. BBA Biomembr. 1995, 1235, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Mochizuki, S.; Kanegae, N.; Nishina, K.; Kamikawa, Y.; Koiwai, K.; Masunaga, H.; Sakurai, K. The role of the helper lipid dioleoylphosphatidylethanolamine (DOPE) for DNA transfection cooperating with a cationic lipid bearing ethylenediamine. Biochim. Biophys. Acta Biomembr. 2013, 1828, 412–418. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Lee, R.J. The role of helper lipids in lipid nanoparticles (LNPs) designed for oligonucleotide delivery. Adv. Drug Deliv. Rev. 2016, 99, 129–137. [Google Scholar] [CrossRef]
- Goncharova, E.P.; Sen‘kova, A.V.; Savin, I.A.; Kabilova, T.O.; Zenkova, M.A.; Vlassov, V.V.; Chernolovskaya, E.L. Immunostimulating rna delivered by p1500 pegylated cationic liposomes limits influenza infection in c57bl/6 mice. Pharmaceutics 2020, 12, 875. [Google Scholar] [CrossRef]
- Inglut, C.T.; Sorrin, A.J.; Kuruppu, T.; Vig, S.; Cicalo, J.; Ahmad, H.; Huang, H.C. Immunological and toxicological considerations for the design of liposomes. Nanomaterials 2020, 10, 190. [Google Scholar] [CrossRef] [Green Version]
- Perche, F.; Torchilin, V.P. Recent Trends in Multifunctional Liposomal Nanocarriers for Enhanced Tumor Targeting. J. Drug Deliv. 2013, 2013, 705265. [Google Scholar] [CrossRef] [Green Version]
- Semple, S.C.; Akinc, A.; Chen, J.; Sandhu, A.P.; Mui, B.L.; Cho, C.K.; Sah, D.W.Y.; Stebbing, D.; Crosley, E.J.; Yaworski, E.; et al. Rational design of cationic lipids for siRNA delivery. Nat. Biotechnol. 2010, 28, 172–176. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cationic Liposomes | N/P | Diameter, nm | Polydispersity Index (PI) | ξ-Potential, mV |
|---|---|---|---|---|
| L | - | 93.5 ± 1.9 | 0.4 ± 0.09 | 45.8 ± 1.1 |
| 1/1 | 152.6 ± 5.3 | 0.27 ± 0.04 | −14.9 ± 1.8 | |
| F | - | 117.8 ± 8.9 | 0.5 ± 0.05 | 21.9 ± 1.5 |
| 1/1 | 175.2 ± 22.6 | 0.3 ± 0.04 | −3 ± 0.043 |
| Group | Average Weight, g | Liver Index 1 | Renal Index 1 | Spleen Index 1 |
|---|---|---|---|---|
| Control | 22.4 ± 0.2 | 5.8 ± 0.2 | 0.7 ± 0.07 | 0.5 ± 0.01 |
| L (1 TD 2) | 23.2 ± 0.5 | 5.9 ± 0.2 | 0.8 ± 0.08 | 0.4 ± 0.01 |
| L (5 TD 2) | 22.6 ± 0.3 | 5.8 ± 0.1 | 0.7 ± 0.04 | 0.4 ± 0.01 |
| F (1 TD 2) | 24.0 ± 0.6 | 4.8 ± 0.3 | 0.6 ± 0.03 | 0.3 ± 0.02 |
| F (5 TD 2) | 25.6 ± 0.6 | 5.6 ± 0.3 | 0.7 ± 0.03 | 0.3 ± 0.02 |
| Group | Average Weight, g | Liver Index 1 | Renal Index 1 | Spleen Index 1 | Heart Index 1 | Lung Index 1 |
|---|---|---|---|---|---|---|
| Control | 20.7 ± 0.5 | 5.5 ± 0.1 | 0.7 ± 0.04 | 0.4 ± 0.02 | 0.5 ± 0.03 | 1.2 ± 0.06 |
| L (1 TD 2) | 21.0 ± 0.4 | 5.3 ± 0.1 | 0.7 ± 0.02 | 0.4 ± 0.02 | 0.6 ± 0.04 | 1.2 ± 0.1 |
| L (2 TD 2) | 20.9 ± 0.3 | 5.1 ± 0.2 | 0.6 ± 0.03 | 0.4 ± 0.03 | 0.6 ± 0.02 | 1.2 ± 0.04 |
| Blood Parameter | Day 0 1 | OptiMEM | L | ||||
|---|---|---|---|---|---|---|---|
| 1 TD 2 | 2 TD 2 | ||||||
| Day 21 | Day 60 | Day 21 | Day 60 | Day 21 | Day 60 | ||
| Leukocytes × 109/L | 2.5 ± 0.2 | 3.9 ± 0.1 | 3.0 ± 0.3 | 3.1 ± 0.2 | 2.7 ± 0.4 | 2.6 ± 0.3 | 2.4 ± 0.2 |
| Lymphocytes × 109/L | 2.2 ± 0.2 | 2.9 ± 0.3 | 2.5 ± 0.2 | 2.6 ± 0.2 | 2.4 ± 0.4 | 2.2 ± 0.2 | 1.9 ± 0.2 |
| Monocytes × 109/L | 0.2 ± 0.1 | 0.5 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 |
| Granulocytes × 109/L | 0.2 ± 0.1 | 0.5 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.3 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 |
| Erythrocytes × 1012/L | 9.7 ± 0.3 | 9.0 ± 0.2 | 8.6 ± 0.2 | 9.4 ± 0.2 | 8.7 ± 0.5 | 9.0 ± 0.2 | 8.8 ± 0.2 |
| Thrombocytes × 109/L | 514 ± 94.8 | 833 ± 113 | 757 ± 45.9 | 776 ± 43.6 | 735 ± 74.9 | 920 ± 39.9 | 757 ± 52.3 |
| Hemoglobin g/L | 128 ± 4.1 | 107 ± 3.4 | 107 ± 3.1 | 113 ± 2.9 | 105 ± 5.9 | 111 ± 3.1 | 115 ± 3.7 |
| Morphologic Parameters of Tumor | Control | PCHT | PCHT + siMDR/F | PCHT + siScr/F |
|---|---|---|---|---|
| Vv 1 normal tissue, % | 33.1 ± 2.9 | 45.3 ± 3.6 * | 77.7 ± 3.4 *,# | 52.4 ± 3.9 * |
| Vv necrosis, % | 33.7 ± 2.1 | 26.5 ± 2 * | 14.9 ± 2.5 *,# | 24.9 ± 4.3 * |
| Vv inflammation, % | 33.0 ± 2.6 | 27.7 ± 3.1 | 7.3 ± 1.1 *,# | 22.8 ± 1.8 * |
| Nv 2 mitosis | 2.2 ± 0.2 | 2.2 ± 0.3 | 1.4 ± 0.2 *,# | 2.4 ± 0.3 |
| Nv caspase-3 positive cells | 7.1 ± 1.8 | 16.2 ± 3.7 * | 23.9 ± 0.5 *,# | 12.9 ± 1 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gladkikh, D.V.; Sen′kova, A.V.; Chernikov, I.V.; Kabilova, T.O.; Popova, N.A.; Nikolin, V.P.; Shmendel, E.V.; Maslov, M.A.; Vlassov, V.V.; Zenkova, M.A.; et al. Folate-Equipped Cationic Liposomes Deliver Anti-MDR1-siRNA to the Tumor and Increase the Efficiency of Chemotherapy. Pharmaceutics 2021, 13, 1252. https://doi.org/10.3390/pharmaceutics13081252
Gladkikh DV, Sen′kova AV, Chernikov IV, Kabilova TO, Popova NA, Nikolin VP, Shmendel EV, Maslov MA, Vlassov VV, Zenkova MA, et al. Folate-Equipped Cationic Liposomes Deliver Anti-MDR1-siRNA to the Tumor and Increase the Efficiency of Chemotherapy. Pharmaceutics. 2021; 13(8):1252. https://doi.org/10.3390/pharmaceutics13081252
Chicago/Turabian StyleGladkikh, Daniil V., Aleksandra V. Sen′kova, Ivan V. Chernikov, Tatyana O. Kabilova, Nelly A. Popova, Valery P. Nikolin, Elena V. Shmendel, Mikhail A. Maslov, Valentin V. Vlassov, Marina A. Zenkova, and et al. 2021. "Folate-Equipped Cationic Liposomes Deliver Anti-MDR1-siRNA to the Tumor and Increase the Efficiency of Chemotherapy" Pharmaceutics 13, no. 8: 1252. https://doi.org/10.3390/pharmaceutics13081252
APA StyleGladkikh, D. V., Sen′kova, A. V., Chernikov, I. V., Kabilova, T. O., Popova, N. A., Nikolin, V. P., Shmendel, E. V., Maslov, M. A., Vlassov, V. V., Zenkova, M. A., & Chernolovskaya, E. L. (2021). Folate-Equipped Cationic Liposomes Deliver Anti-MDR1-siRNA to the Tumor and Increase the Efficiency of Chemotherapy. Pharmaceutics, 13(8), 1252. https://doi.org/10.3390/pharmaceutics13081252